Partial Loss of Inheritable Type I Resistance of Codling Moth to Cydia pomonella granulovirus




Abstract
:1. Introduction
2. Material and Methods
2.1. Insects and Virus
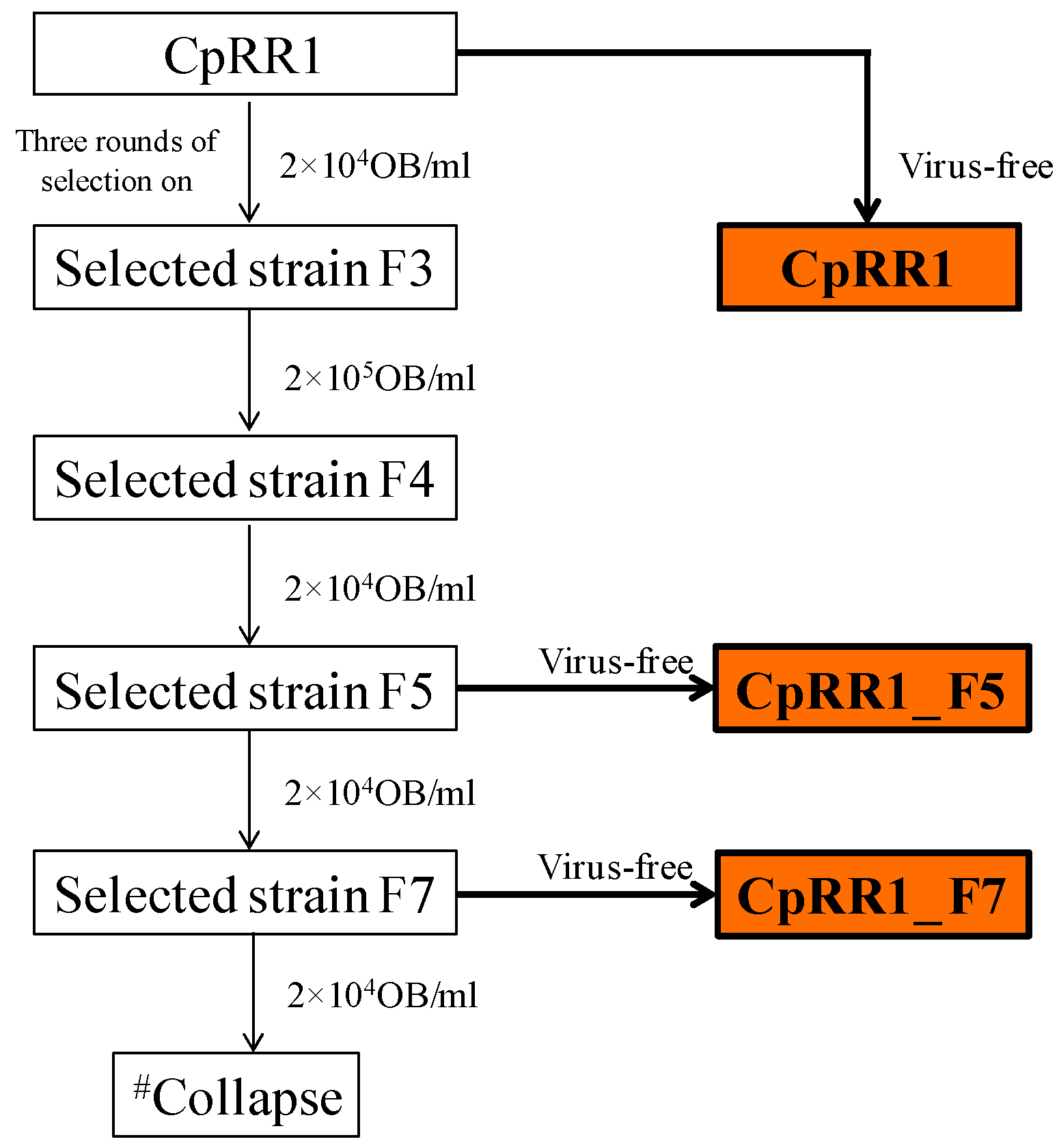
2.2. Re-Selection of CpRR1
2.3. Bioassays
2.4. Single-Pair Crossings
3. Results
3.1. Mass Crossing Selection
3.2. Resistance Ratio
3.3. Single-Pair Crossing
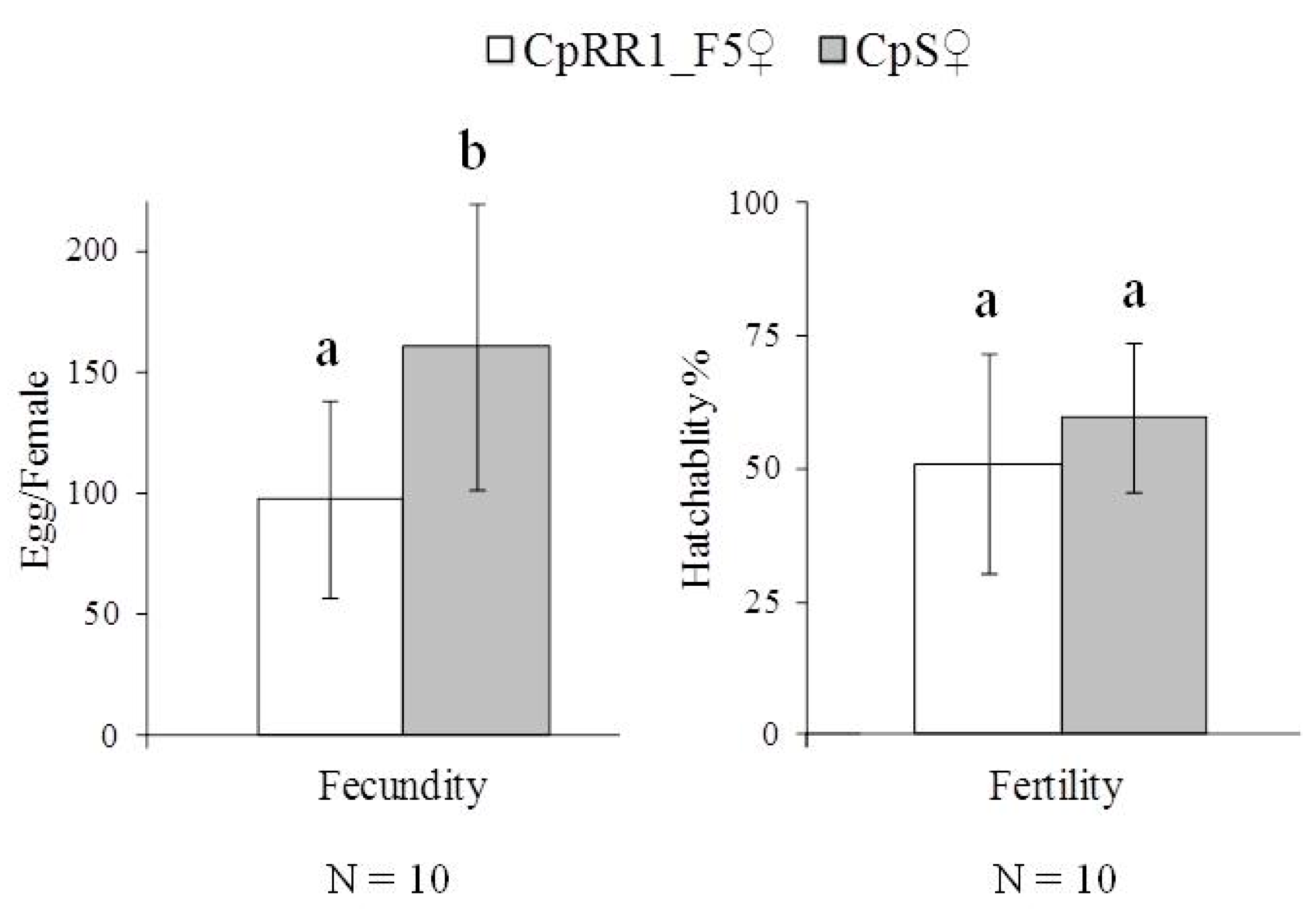
3.4. Fitness Cost
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Eberle, K.E.; Jehle, J.A. Field resistance of codling moth against Cydia pomonella granulovirus (CpGV) is autosomal and incompletely dominant inherited. J. Invertebr. Pathol. 2006, 93, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Thomson, D.; Vincent, C.; Arthurs, S.P. Codling moth granulovirus: A comprehensive review. Biocontrol. Sci. Technol. 2008, 18, 639–663. [Google Scholar] [CrossRef]
- Huber, J. Western Europe. In Insect Viruses and Pest Management; Hunter-Fujita, F.R., Entwistle, P.F., Evans, H.F., Crook, N.E., Eds.; Wiley & Sons: Chichester, UK, 1998; pp. 201–215. [Google Scholar]
- Fritsch, E.; Undorf-Spahn, K.; Kienzle, J.; Zebitz, C.P.W.; Huber, J. Apfelwickler-Granulovirus: Erste Hinweise auf Unterschiede in der Empfindlichkeit lokaler Apfelwickler-Populationen [Codling moth granulovirus: First indication of variations in the susceptibility of local codling moth populations]. Nachr. Deutsch. Pflanzenschutzd. 2005, 57, 29–34. [Google Scholar]
- Asser-Kaiser, S.; Fritsch, E.; Undorf-Spahn, K.; Kienzle, J.; Eberle, K.E.; Gund, N.A.; Reineke, A.; Zebitz, C.P.; Heckel, D.G.; Huber, J.; et al. Rapid emergence of baculovirus resistance in codling moth due to dominant, sex-linked inheritance. Science 2007, 317, 1916–1918. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Bisutti, I.L.; Ladurner, E.; Benuzzi, M.; Sauphanor, B.; Kienzle, J.; Zingg, D.; Undorf-Spahn, K.; Fritsch, E.; Huber, J.; et al. The occurrence and distribution of resistance of codling moth to Cydia pomonella granulovirus in Europe. J. Appl. Entomol. 2013, 137, 641–649. [Google Scholar] [CrossRef]
- Zichová, T.; Stará, J.; Kundu, J.K.; Eberle, K.E.; Jehle, J.A. Resistance to Cydia pomonella granulovirus follows a geographically widely distributed inheritance type within Europe. Biocontrol 2013, 58, 525–534. [Google Scholar] [CrossRef]
- Asser-Kaiser, S.; Heckel, D.G.; Jehle, J.A. Sex linkage of CpGV resistance in a heterogeneous field strain of the codling moth Cydia pomonella (L.). J. Invertebr. Pathol. 2010, 103, 59–64. [Google Scholar] [CrossRef]
- Sauphanor, B.; Berling, M.; Toubon, J.-F.; Reyes, M.; Delnatte, J.; Allemoz, P. Carpocapse des pommes cas de résistance au virus de la granulose en vergers biologiques: Fruits et légumes. Phytoma-La Déf. Vég. 2006, 590, 24–27. [Google Scholar]
- Jehle, J.A.; Schulze-Bopp, S.; Undorf-Spahn, K.; Fritsch, E. Evidence for a second type of resistance against Cydia pomonella granulovirus in field populations of codling moths. Appl. Environ. Microbiol. 2017, 83, e02330-16. [Google Scholar] [CrossRef]
- Undorf-Spahn, K.; Fritsch, E.; Huber, J.; Kienzle, J.; Zebitz, C.P.; Jehle, J.A. High stability and no fitness costs of the resistance of codling moth to Cydia pomonella granulovirus (CpGV-M). J. Invertebr. Pathol. 2012, 111, 136–142. [Google Scholar] [CrossRef]
- Berling, M.; Sauphanor, B.; Bonhomme, A.; Siegwart, M.; Lopez-Ferber, M. A single sex-linked dominant gene does not fully explain the codling moth’s resistance to granulovirus. Pest Manag. Sci. 2013, 69, 1261–1266. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, M.M.; Eberle, K.E.; Radtke, P.; Jehle, J.A. Baculovirus resistance in codling moth is virus isolate-dependent and the consequence of a mutation in viral gene pe38. Proc. Natl. Acad. Sci. USA 2014, 111, 15711–15716. [Google Scholar] [CrossRef] [PubMed]
- Sauer, A.J.; Fritsch, E.; Undorf-Spahn, K.; Nguyen, P.; Marec, F.; Heckel, D.G.; Jehle, J.A. Novel resistance to Cydia pomonella granulovirus (CpGV) in codling moth shows autosomal and dominant inheritance and confers cross-resistance to different CpGV genome groups. PLoS ONE 2017, 12, e0179157. [Google Scholar] [CrossRef] [PubMed]
- Sauer, A.J.; Schulze-Bopp, S.; Fritsch, E.; Undorf-Spahn, K.; Jehle, J.A. A third type of resistance to Cydia pomonella granulovirus in codling moths shows a mixed Z-Linked and autosomal inheritance pattern. Appl. Environ. Microbiol. 2017, 83, e01036-17. [Google Scholar] [CrossRef] [PubMed]
- Pietruska, D. Vergleichende Transkriptomanalysen Des Cydia pomonella granulovirus (CpGV) in sensiblen Und resistenten Apfelwicklern (Cydia pomonella). Ph.D. Thesis, Johannes Gutenberg-University, Mainz, Germany, 2018; p. 199. [Google Scholar]
- Ivaldi-Sender, C. Techniques simples pour un elevage permanent de la Tordeuse orientale, Grapholita molesta (Lepidoptera, Tortricidae) sur milieu artificiel. Ann. Zool. Ecol. Anim. 1974, 6, 337–343. [Google Scholar]
- Tanada, Y.A. Granulosis virus of codling moth, Carpocapsa pomenella (Linnaeus) (Olethreutidae, Lepidoptera). J. Insect. Pathol. 1964, 6, 39–47. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Milks, M.L.; Myers, J.H. The development of larval resistance to a nucleopolyhedrovirus is not accompanied by an increased virulence in the virus. Evol. Ecol. 2000, 14, 645–664. [Google Scholar] [CrossRef]
- Asser-Kaiser, S.; Radtke, P.; El-Salamouny, S.; Winstanley, D.; Jehle, J.A. Baculovirus resistance in codling moth (Cydia pomonella L.) caused by early block of virus replication. Virology 2011, 410, 360–367. [Google Scholar] [CrossRef]
- Fan, J.; Wennmann, J.T.; Wang, D.; Jehle, J.A. Novel diversity and virulence patterns found in new isolates of Cydia pomonella granulovirus from China. 2019; Manuscript submitted. [Google Scholar]
- Fuxa, J.R.; Richter, A.R. Repeated reversion of resistance to nucleopolyhedrovirus by Anticarsia gemmatalis. J. Invertebr. Pathol. 1998, 71, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Iwata, K.; Haas-Stapleton, E.; Kunimi, Y.; Inoue, M.N.; Nakai, M. Midgut-based resistance to oral infection by a nucleopolyhedrovirus in the laboratory-selected strain of the smaller tea tortrix, Adoxophyes honmai (Lepidoptera: Tortricidae). J. Gen. Virol. 2017, 98, 296–304. [Google Scholar] [CrossRef]
- Nakai, M.; Takahashi, K.; Iwata, K.; Tanaka, K.; Koyanagi, J.; Ookuma, A.; Takatsuka, J.; Okuno, S.; Kunimi, Y. Acquired resistance to a nucleopolyhedrovirus in the smaller tea tortrix Adoxophyes honmai (Lepidoptera: Tortricidae) after selection by serial viral administration. J. Invertebr. Pathol. 2017, 145, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Berling, M.; Blachere-Lopez, C.; Soubabere, O.; Lery, X.; Bonhomme, A.; Sauphanor, B.; Lopez-Ferber, M. Cydia pomonella granulovirus genotypes overcome virus resistance in the codling moth and improve virus efficiency by selection against resistant hosts. Appl. Environ. Microbiol. 2009, 75, 925–930. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Generation | N | Larval Instar | CpGV Seletion (OBs/mL) | Surviving Pupae (%) | Surviving Adult (%) | Sex Ratio (% Male) [m:f] |
|---|---|---|---|---|---|---|
| F0 | 300 | L5 + L4 | 2 × 104 | 27.3 | 22.0 | 53.7 [36:31] |
| F1 | 100 | L4 | 2 × 104 | 11.0 | 9.0 | 66.7 [6:3] |
| F2 | 326 | L1 | 2 × 104 | 8.6 | 7.7 | 64.0 [16:9] |
| F3 | 196 | L1 | 2 × 105 | 4.1 | 3.1 | 16.7 [1:5] |
| F4 | 363 | L1 | 2 × 104 | 17.6 | 12.4 | 53.3 [24:21] |
| F5 | 95 | L1 | 2 × 104 | 2.1 | 2.1 | 0 [0:2] |
| F6a | 92 | L1 | 2 × 104 | 5.4 | 5.4 | 60.0 [3:2] |
| F7# | 163 | L1 | 2 × 104 | 0.6 | 0.6 | collapse [1:0] |
| Strain | N | 7 dpi | 14 dpi | ||||||
|---|---|---|---|---|---|---|---|---|---|
| LC50 (95% CI) [OBs/mL] | Slope ± SE | χ2 (df) | Potency | LC50 (95% CI) [OBs/mL] | Slope ± SE | χ2 (df) | Potency | ||
| CpS | 1075 | 1.99 (0.98–4.05) × 103 | 1.26 ± 0.04 | 30.81 (4) | 1 | 4.50 (2.06–11.90) × 102 | 1.01 ± 0.04 | 14.72 (4) | 1 |
| CpRR1 | 1550 | 1.27 (1.08–1.63) × 105 | 2.36 ± 0.15 | 2.37 (4) | 64 | 1.60 (1.41–1.81) × 104 | 1.97 ± 0.05 | 6.47 (4) | 36 |
| CpRR1_F5 | 362 | >2 × 107 | 0.45 ± 0.10 | 3.17 (3) | >10,000 | 2.32 (0.98–13.22) × 105 | 0.77 ± 0.09 | 4.26 (3) | 517 |
| Strain | Crosses | N, nn | Progeny Genotypes, by Z Hypothesis$ | Observed Mortality (%) at 7 dpi | Observed Mortality (%) at 14 dpi | Pupae | |
|---|---|---|---|---|---|---|---|
| progeny | Exp. mort. at 7 dpi (%) | Mean ± SD | Mean ± SD | Sex Ratio (% male) [m:f] | |||
| CpS | CpSf × CpSm | 4, 318 | ZSZS, ZSW | 100 | 96.7 ± 3.9 | 98.3 ± 3.4 | - |
| CpRR1 | CpRR1f × CpRR1m | 5, 437 | ZRZR, ZRW | 0 | 40.9 ± 4.3 | 79.2 ± 8.5 | - |
| CpRR1_F5 | CpRR1_F5f × CpRR1_F5m | 7, 363 | ZRZR, ZRW | 0 | 49.2 ± 25.1 | 85.8 ± 14.5 | n.d. |
| CpRR1_F5 | CpRR1_F5f × CpSm | 8, 302 | ZRZS, ZSW | 50 | 63.4 ± 15.9 | 87.2 ± 14.1 | n.d. |
| CpRR1_F5m × CpSf | 11, 638 | ZRZS, ZRW | 0 | 12.1 ± 8.9 | 57.4 ± 23.4 | 28.6 [2:5] | |
| CpRR1_F7 | CpRR1_F7f × CpSm | 13, 518 | ZRZS, ZSW | 50 | 16.0 ± 8.4 | 16.3 ± 13.1 | 41.2 [7:10] |
| CpRR1_F7m × CpSf | 8, 452 | ZRZS, ZRW | 0 | *3.8 ± 5.2 | *4.1 ± 7.7 | n.d. | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Wennmann, J.T.; Jehle, J.A. Partial Loss of Inheritable Type I Resistance of Codling Moth to Cydia pomonella granulovirus. Viruses 2019, 11, 570. https://doi.org/10.3390/v11060570
Fan J, Wennmann JT, Jehle JA. Partial Loss of Inheritable Type I Resistance of Codling Moth to Cydia pomonella granulovirus. Viruses. 2019; 11(6):570. https://doi.org/10.3390/v11060570
Chicago/Turabian StyleFan, Jiangbin, Jörg T. Wennmann, and Johannes A. Jehle. 2019. "Partial Loss of Inheritable Type I Resistance of Codling Moth to Cydia pomonella granulovirus" Viruses 11, no. 6: 570. https://doi.org/10.3390/v11060570
APA StyleFan, J., Wennmann, J. T., & Jehle, J. A. (2019). Partial Loss of Inheritable Type I Resistance of Codling Moth to Cydia pomonella granulovirus. Viruses, 11(6), 570. https://doi.org/10.3390/v11060570