A Novel Alphabaculovirus from the Soybean Looper, Chrysodeixis includens, that Produces Tetrahedral Occlusion Bodies and Encodes Two Copies of he65


Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Production and Isolation
2.2. Electron Microscopy
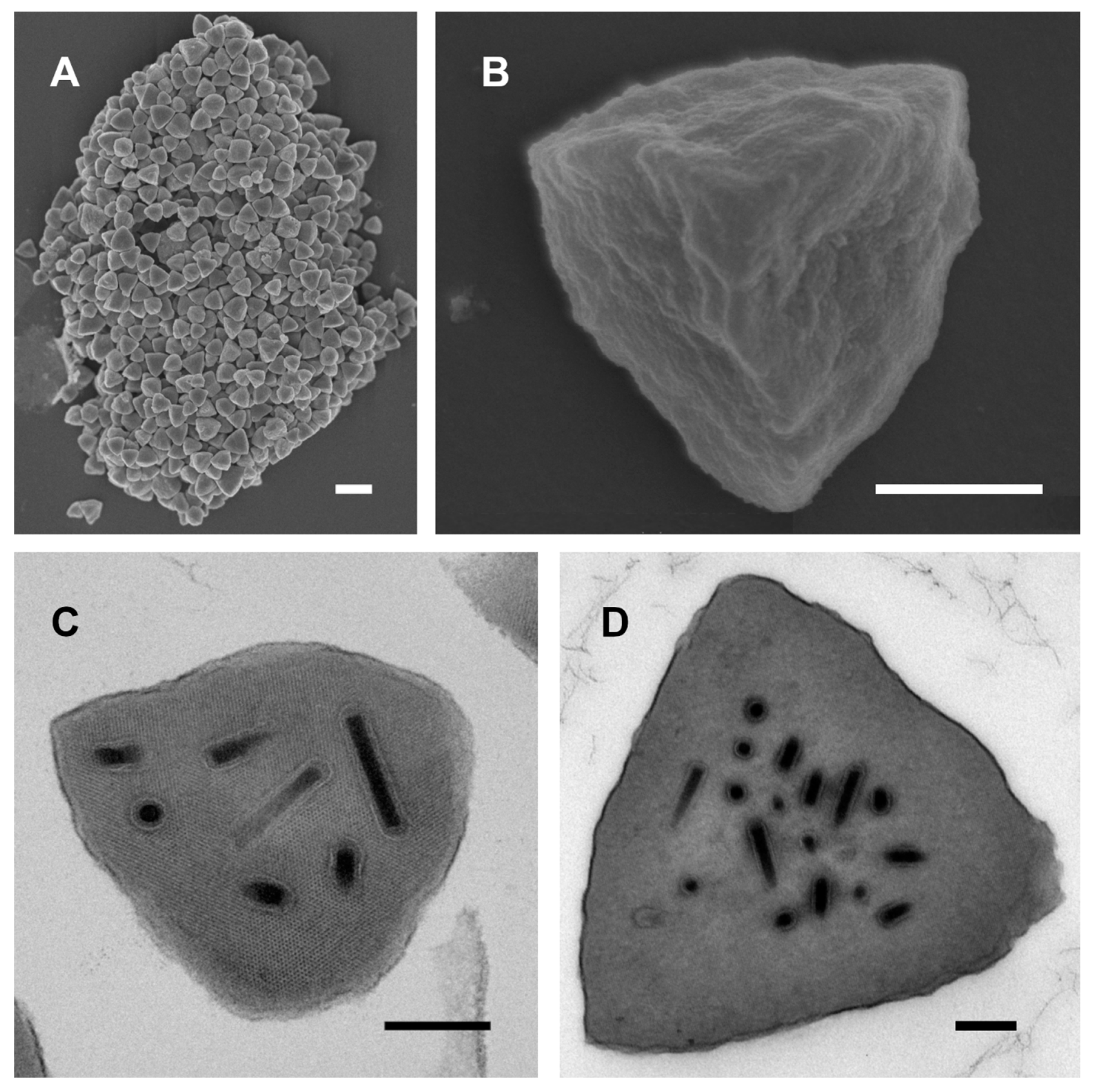
2.2.1. Scanning Electron Microscopy (SEM)
2.2.2. Transmission Electron Microscopy (TEM)
2.3. Bioassays
2.4. Viral DNA Isolation and Sequencing
2.5. Genome Feature Identification
2.6. Sequence Comparison and Phylogeny
3. Results
3.1. Ultrastructural Features of ChinNPV#1 OBs
3.2. Pathogenicity of ChinNPV#1
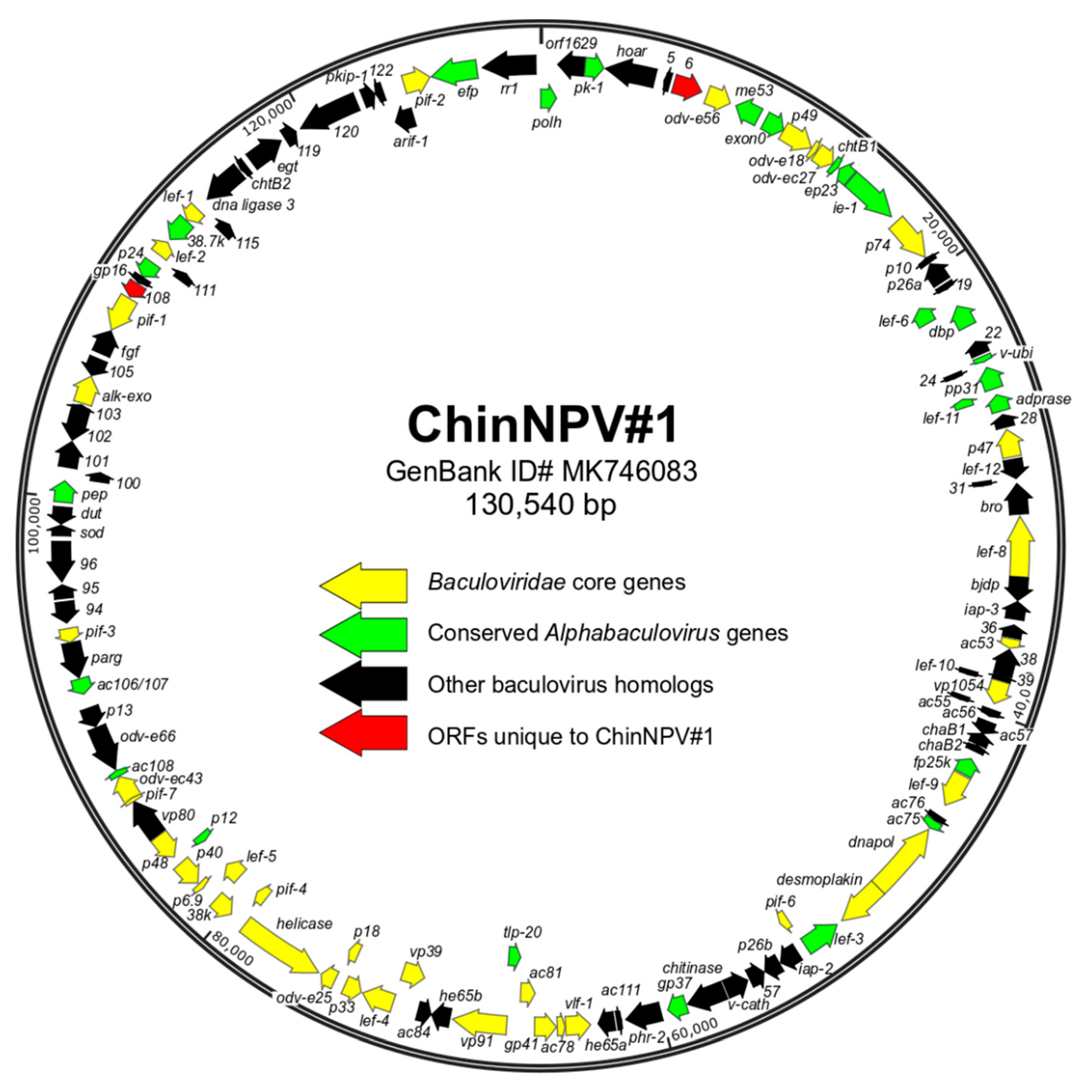
3.3. Characteristics of the ChinNPV#1 Genome Sequence
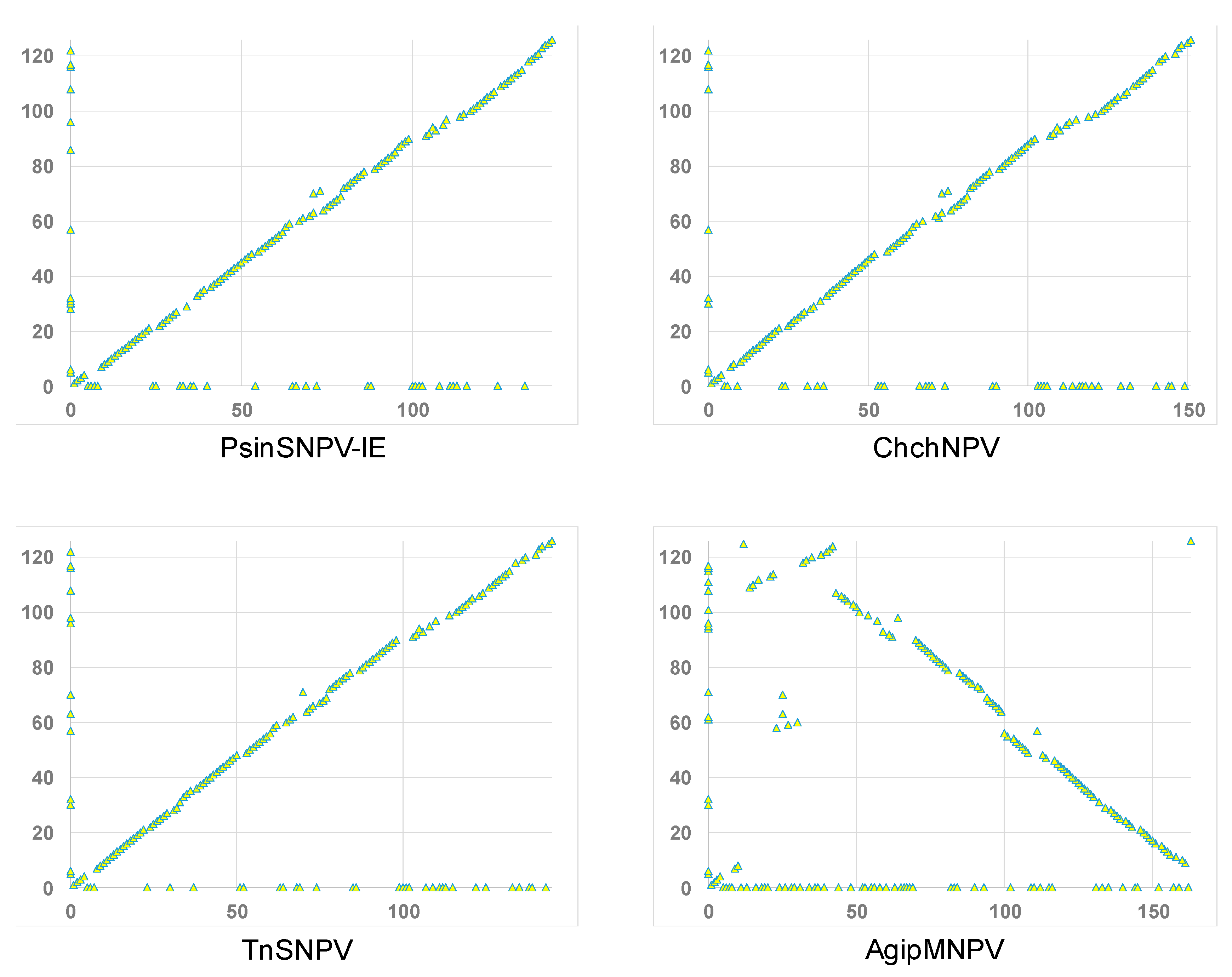
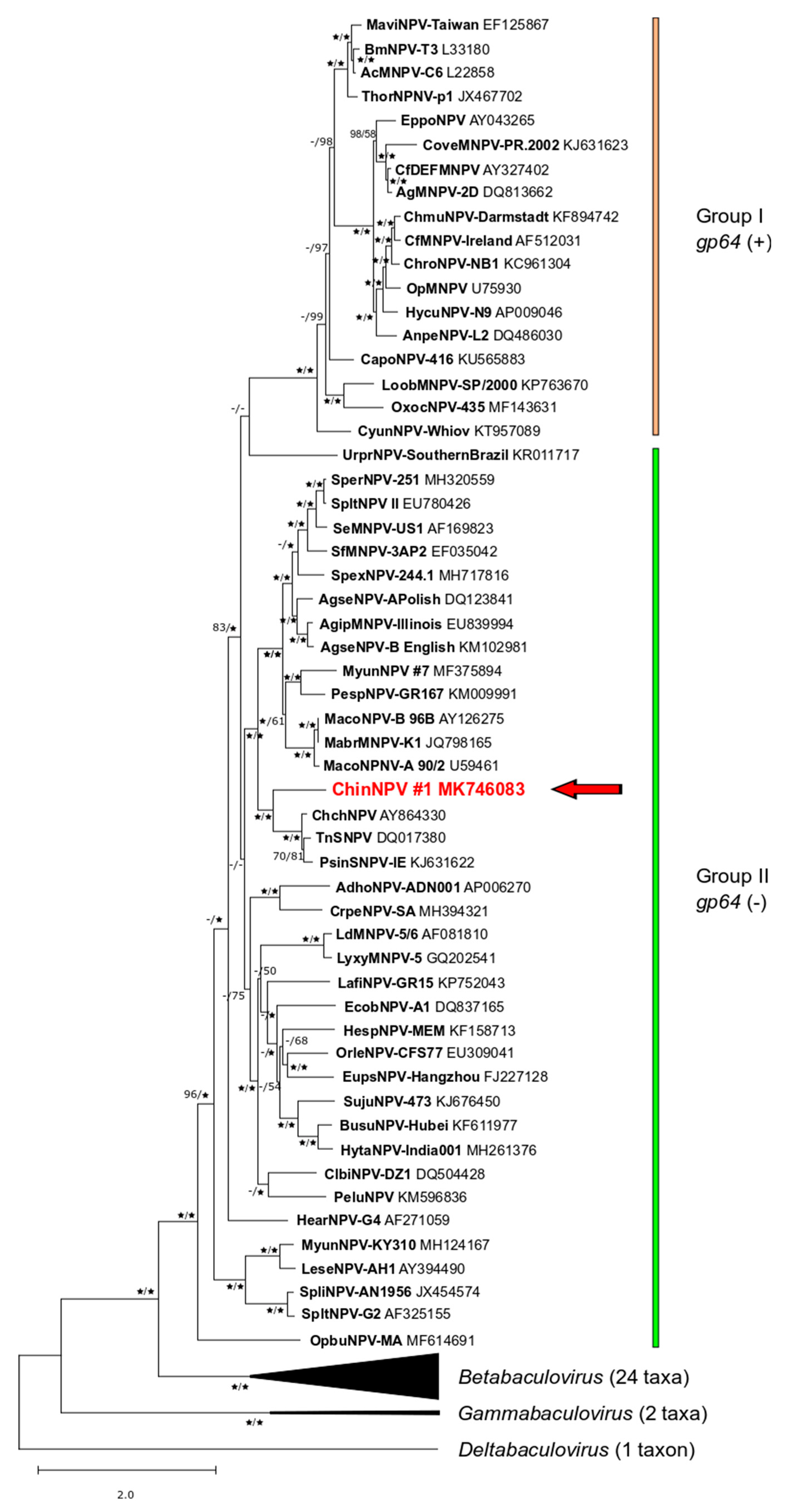
3.4. Relationship of ChinNPV#1 to Other Baculoviruses
3.5. ORF content of ChinNPV#1
3.5.1. Core Genes, Gene Families, and ORFs Missing from/Unique to ChinNPV#1
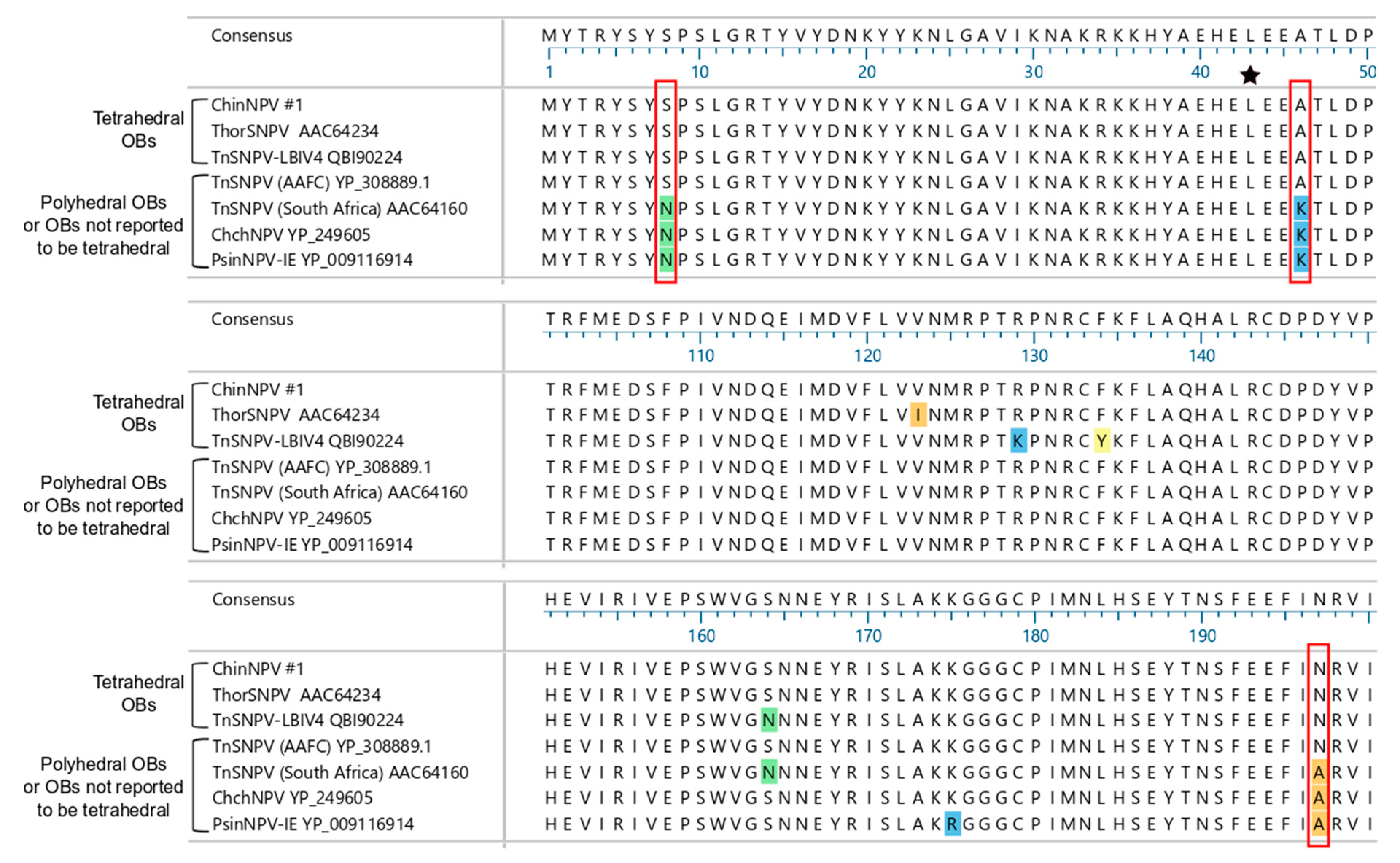
3.5.2. Polyhedrin
3.5.3. Lef-12
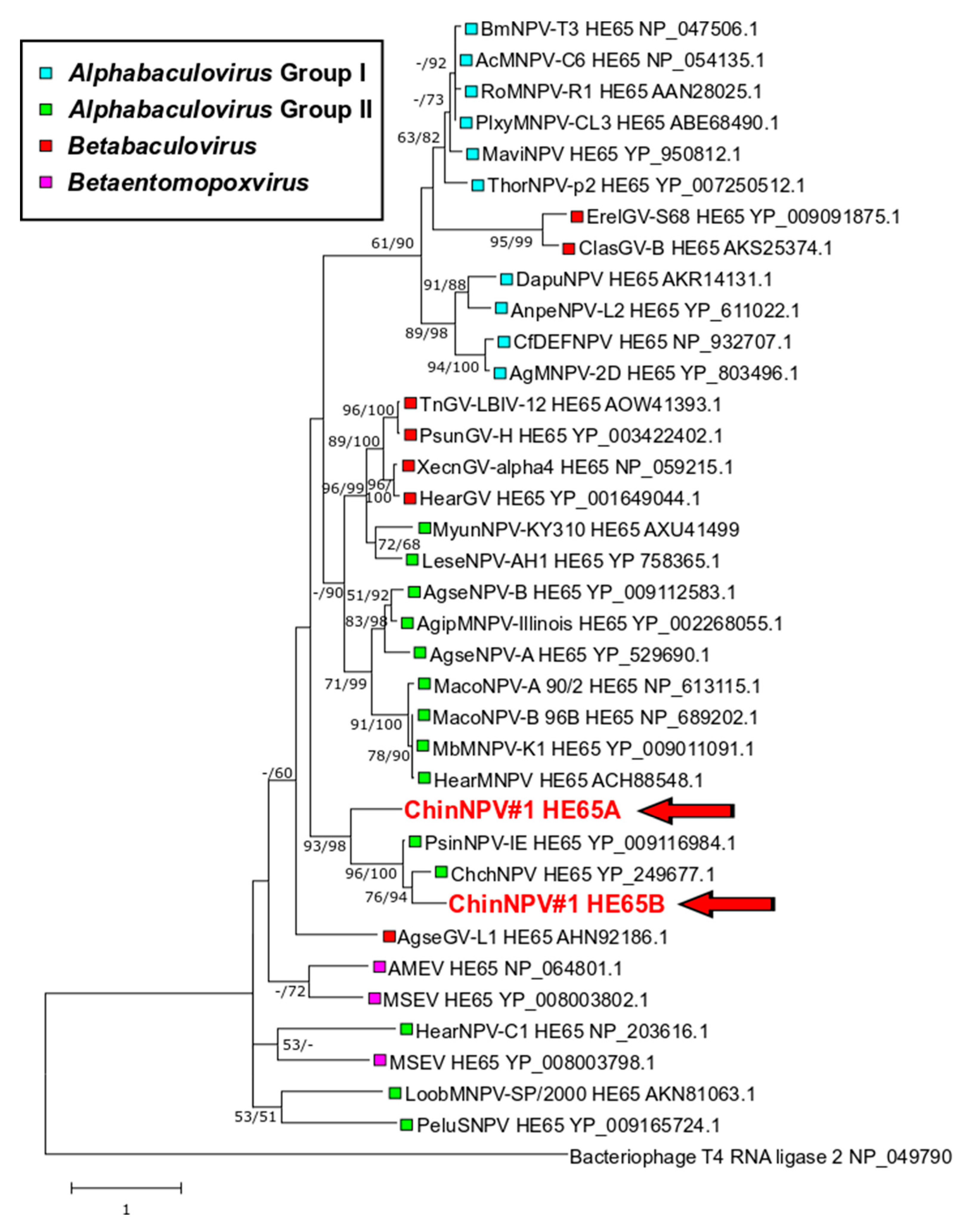
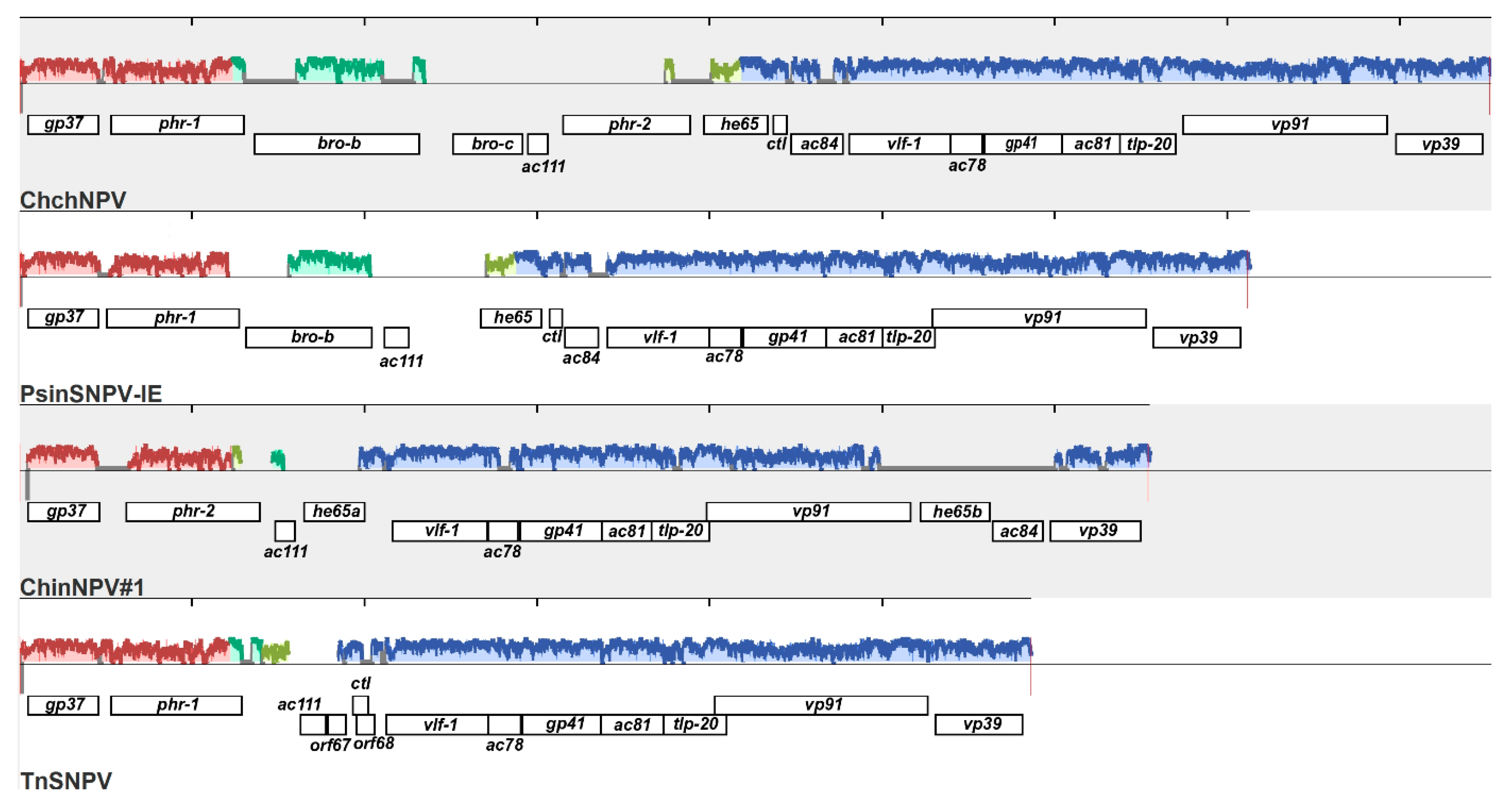
3.5.4. Two Full-Length Copies of he65
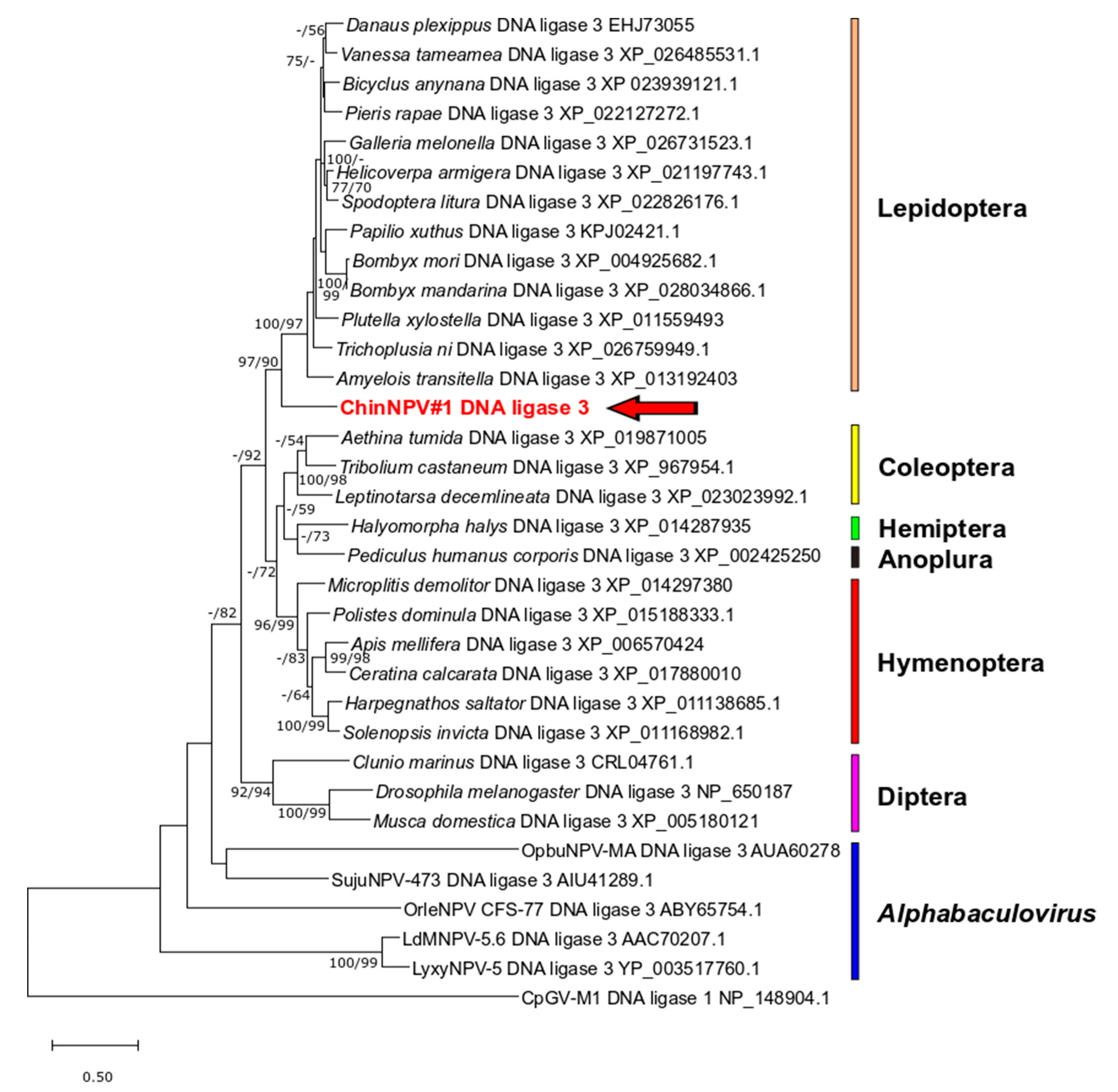
3.5.5. DNA Ligase 3
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eichlin, T.D.; Cunningham, H.B. The Plusiinae (Lepidoptera: Noctuidae) of America north of Mexico, Emphasizing Genitalic and Larval Morphology; Technical Bulletin no. 1567; United States Department of Agriculture, Agricultural Research Service: Washington, DC, USA, 1978; p. 122.
- Kogan, M.; Turnipseed, S.G. Ecology and management of soybean arthropods. Ann. Rev. Entomol. 1987, 32, 507–538. [Google Scholar] [CrossRef]
- Bortolotto, O.C.; de F. Bueno, R.C.O.; de F. Bueno, A.; da Kruz, A.K.S.; Queiroz, A.P.; Sanzovo, A.; Ferreira, R.B. The use of soybean integrated pest management in Brazil: A review. Agron. Sci. Biotechnol. 2015, 1, 25–32. [Google Scholar]
- Mascarenhas, R.N.; Boethel, D.J. Responses of field-collected strains of soybean looper (Lepidoptera:Noctuidae) to selected insecticides using an artificial diet overlay bioassay. J. Econ. Entomol. 1997, 90, 1117–1124. [Google Scholar] [CrossRef]
- Owen, L.N.; Catchot, A.L.; Musser, F.R.; Gore, J.; Cook, D.C.; Jackson, R. Susceptibility of Chrysodeixis includens (Lepidoptera:Noctuidae) to reduced-risk insecticides. Fla. Entomol. 2013, 96, 554–559. [Google Scholar] [CrossRef]
- Bueno, R.C.O.D.; Parra, J.R.P.; Bueno, A.D. Trichogramma pretiosum parasitism of Pseudoplusia includens and Anticarsia gemmatalis eggs at different temperatures. Biol. Control 2012, 60, 154–162. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, S.; Sciocco-Cap, A.; Romanowski, V. Baculovirus insecticides in Latin America: Historical overview, current status and future perspectives. Viruses 2015, 7, 2230–2267. [Google Scholar] [CrossRef] [PubMed]
- Rohrmann, G.F. Baculovirus Molecular Biology, 3rd ed.; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2013.
- Harrison, R.L.; Herniou, E.A.; Jehle, J.A.; Theilmann, D.A.; Burand, J.P.; Becnel, J.J.; Krell, P.J.; van Oers, M.M.; Mowery, J.D.; Bauchan, G.R.; et al. ICTV Virus Taxonomy Profile: Baculoviridae. J. Gen. Virol. 2018, 99, 1185–1186. [Google Scholar] [CrossRef] [PubMed]
- Livingston, J.M.; Yearian, W.C. Nuclear polyhedrosis virus of Pseudoplusia includens (Lepidoptera-Noctuidae). J. Invert. Path. 1972, 19, 107. [Google Scholar] [CrossRef]
- Alexandre, T.M.; Ribeiro, Z.M.; Craveiro, S.R.; Cunha, F.; Fonseca, I.C.; Moscardi, F.; Castro, M.E. Evaluation of seven viral isolates as potential biocontrol agents against Pseudoplusia includens (Lepidoptera: Noctuidae) caterpillars. J. Invertebr. Pathol. 2010, 105, 98–104. [Google Scholar] [CrossRef]
- Livingston, J.M.; Mcleod, P.J.; Yearian, W.C.; Young, S.Y. Laboratory and field evaluation of a nuclear polyhedrosis virus of the soybean looper, Pseudoplusia includens. J. Georgia Entomol. So. 1980, 15, 194–199. [Google Scholar]
- Mcleod, P.J.; Young, S.Y.; Yearian, W.C. Application of a baculovirus of Pseudoplusia includens to soybean—efficacy and seasonal persistence (Lepidoptera, Noctuidae). Environ. Entomol. 1982, 11, 412–416. [Google Scholar] [CrossRef]
- Muraro, D.S.; Giacomelli, T.; Stacke, R.F.; Godoy, D.N.; Marcon, P.; Popham, H.J.R.; Bernardi, O. Baseline susceptibility of Brazilian populations of Chrysodeixis includens (Lepidoptera: Noctuidae) to C. includens nucleopolyhedrovirus and diagnostic concentration for resistance monitoring. J. Econ. Entomol. 2019, 112, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Godoy, D.N.; Führ, F.M.; Stacke, R.F.; Muraro, D.S.; Marçon, P.; Popham, H.J.R.; Bernardi, O. No cross-resistance between ChinNPV and chemical insecticides in Chrysodeixis includens (Lepidoptera: Noctuidae). J. Invert. Path. 2019, 164, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Craveiro, S.R.; Melo, F.L.; Ribeiro, Z.M.; Ribeiro, B.M.; Bao, S.N.; Inglis, P.W.; Castro, M.E. Pseudoplusia includens single nucleopolyhedrovirus: Genetic diversity, phylogeny and hypervariability of the pif-2 gene. J. Invertebr. Pathol. 2013, 114, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Craveiro, S.R.; Inglis, P.W.; Togawa, R.C.; Grynberg, P.; Melo, F.L.; Ribeiro, Z.M.; Ribeiro, B.M.; Bao, S.N.; Castro, M.E. The genome sequence of Pseudoplusia includens single nucleopolyhedrovirus and an analysis of p26 gene evolution in the baculoviruses. BMC Genom. 2015, 16, 127. [Google Scholar] [CrossRef] [PubMed]
- Craveiro, S.R.; Santos, L.A.V.M.; Togawa, R.C.; Inglis, P.W.; Grynberg, P.; Ribeiro, Z.M.A.; Ribeiro, B.M.; Castro, M.E.B. Complete genome sequences of six Chrysodeixis includens nucleopolyhedrovirus isolates from Brazil and Guatemala. Microbiol. Resour. Ann. 2016, 4, e01192-16. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2016). Arch. Virol. 2016, 161, 2921–2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, P.R.; van Beek, N.A.M.; Wood, H.A. A modified droplet feeding method for rapid assay of Bacillus thuringiensis and baculoviruses in noctuid larvae. J. Invert. Path. 1986, 48, 187–192. [Google Scholar] [CrossRef]
- Harrison, R.L.; Rowley, D.L.; Mowery, J.D.; Bauchan, G.R.; Burand, J.P. The Operophtera brumata nucleopolyhedrovirus (OpbuNPV) represents an early, divergent lineage within genus Alphabaculovirus. Viruses 2017, 9, 307. [Google Scholar] [CrossRef]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nuc. Acids. Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, L.; Stephens, A.; Nam, S.Z.; Rau, D.; Kubler, J.; Lozajic, M.; Gabler, F.; Soding, J.; Lupas, A.N.; Alva, V. A completely reimplemented MPI bioinformatics toolkit with a new HHpred server at its core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nuc. Acids. Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Jehle, J.A.; Lange, M.; Wang, H.; Hu, Z.; Wang, Y.; Hauschild, R. Molecular identification and phylogenetic analysis of baculoviruses from Lepidoptera. Virology 2006, 346, 180–193. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.H.; Arif, B.M.; Jin, F.; Martens, J.W.; Chen, X.W.; Sun, J.S.; Zuidema, D.; Goldbach, R.W.; Vlak, J.M. Distinct gene arrangement in the Buzura suppressaria single-nucleocapsid nucleopolyhedrovirus genome. J. Gen. Virol. 1998, 79, 2841–2851. [Google Scholar] [CrossRef]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.W.; Carner, G.R. Characterization of a single-nucleocapsid nucleopolyhedrovirus of Thysanoplusia orichalcea L. (Lepidoptera: Noctuidae) from Indonesia. J. Invert. Path. 2000, 75, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Van Oers, M.M.; Vlak, J.M. Baculovirus genomics. Curr. Drug Targets 2007, 8, 1051–1068. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, M.J.; Miele, S.A.; Iserte, J.A.; Belaich, M.N.; Ghiringhelli, P.D. The ac53, ac78, ac101, and ac103 genes are newly discovered core genes in the family Baculoviridae. J. Virol. 2012, 86, 12069–12079. [Google Scholar] [CrossRef] [PubMed]
- van Oers, M.M.; Abma-Henkens, M.H.; Herniou, E.A.; de Groot, J.C.; Peters, S.; Vlak, J.M. Genome sequence of Chrysodeixis chalcites nucleopolyhedrovirus, a baculovirus with two DNA photolyase genes. J. Gen. Virol. 2005, 86, 2069–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, L.G.; Seipp, R.; Stewart, T.M.; Erlandson, M.A.; Theilmann, D.A. Sequence analysis of the complete genome of Trichoplusia ni single nucleopolyhedrovirus and the identification of a baculoviral photolyase gene. Virology 2005, 338, 209–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theze, J.; Lopez-Vaamonde, C.; Cory, J.S.; Herniou, E.A. Biodiversity, evolution and ecological specialization of baculoviruses: A treasure trove for future applied research. Viruses 2018, 10, 366. [Google Scholar] [CrossRef] [PubMed]
- Javed, M.A.; Biswas, S.; Willis, L.G.; Harris, S.; Pritchard, C.; van Oers, M.M.; Donly, B.C.; Erlandson, M.A.; Hegedus, D.D.; Theilmann, D.A. Autographa californica multiple nucleopolyhedrovirus AC83 is a per os infectivity factor (PIF) protein required for occlusion-derived virus (ODV) and budded virus nucleocapsid assembly as well as assembly of the PIF complex in ODV envelopes. J. Virol. 2017, 91, e02115-16. [Google Scholar] [CrossRef]
- Harrison, R.L.; Puttler, B.; Popham, H.J. Genomic sequence analysis of a fast-killing isolate of Spodoptera frugiperda multiple nucleopolyhedrovirus. J. Gen. Virol. 2008, 89, 775–790. [Google Scholar] [CrossRef]
- Wolff, J.L.; Valicente, F.H.; Martins, R.; Oliveira, J.V.; Zanotto, P.M. Analysis of the genome of Spodoptera frugiperda nucleopolyhedrovirus (SfMNPV-19) and of the high genomic heterogeneity in group II nucleopolyhedroviruses. J. Gen. Virol. 2008, 89, 1202–1211. [Google Scholar] [CrossRef]
- del Rincón-Castro, M.C.; Ibarra, J.E. Genotypic divergence of three single nuclear polyhedrosis virus (SNPV) strains from the cabbage looper, Trichoplusia ni. Biochem. Syst. Ecol. 1997, 25, 287–295. [Google Scholar] [CrossRef]
- van Oers, M.M.; Herniou, E.A.; Usmany, M.; Messelink, G.J.; Vlak, J.M. Identification and characterization of a DNA photolyase-containing baculovirus from Chrysodeixis chalcites. Virology 2004, 330, 460–470. [Google Scholar] [CrossRef]
- Fielding, B.C.; Davison, S. The characterization and phylogenetic relationship of the Trichoplusia ni single capsid nuclear polyhedrosis virus polyhedrin gene. Virus Genes 1999, 19, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Kumar, C.M.S.; Jacob, T.K.; Devasahayam, S.; D’Silva, S.; Jinsha, J.; Rajna, S. Occurrence and characterization of a tetrahedral nucleopolyhedrovirus from Spilarctia obliqua (Walker). J. Invert. Path. 2015, 132, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.W.; Carner, G.R.; Fescemyer, H.W. Polyhedrin sequence determines the tetrahedral shape of occlusion bodies in Thysanoplusia orichalcea single-nucleocapsid nucleopolyhedrovirus. J. Gen. Virol. 1998, 79, 2549–2556. [Google Scholar] [CrossRef]
- Todd, J.W.; Passarelli, A.L.; Miller, L.K. Eighteen baculovirus genes, including lef-11, p35, 39K, and p47, support late gene expression. J. Virol. 1995, 69, 968–974. [Google Scholar] [PubMed]
- Rapp, J.C.; Wilson, J.A.; Miller, L.K. Nineteen baculovirus open reading frames, including LEF-12, support late gene expression. J. Virol. 1998, 72, 10197–10206. [Google Scholar]
- Liu, X.; Yin, F.; Zhu, Z.; Hou, D.; Wang, J.; Zhang, L.; Wang, M.; Wang, H.; Hu, Z.; Deng, F. Genomic sequencing and analysis of Sucra jujuba nucleopolyhedrovirus. PLoS ONE 2014, 9, e110023. [Google Scholar] [CrossRef]
- Becker, D.; Knebel-Morsdorf, D. Sequence and temporal appearance of the early transcribed baculovirus gene HE65. J. Virol. 1993, 67, 5867–5872. [Google Scholar] [Green Version]
- Harrison, R.L.; Mowery, J.D.; Rowley, D.L.; Bauchan, G.R.; Theilmann, D.A.; Rohrmann, G.F.; Erlandson, M.A. The complete genome sequence of a third distinct baculovirus isolated from the true armyworm, Mythimna unipuncta, contains two copies of the lef-7 gene. Virus Genes 2018, 54, 297–310. [Google Scholar] [CrossRef]
- Li, L.; Donly, C.; Li, Q.; Willis, L.G.; Keddie, B.A.; Erlandson, M.A.; Theilmann, D.A. Identification and genomic analysis of a second species of nucleopolyhedrovirus isolated from Mamestra configurata. Virology 2002, 297, 226–244. [Google Scholar] [CrossRef] [PubMed]
- Bourner, T.C.; Cory, J.S. Host range of an NPV and a GV isolated from the common cutworm, Agrotis segetum: Pathogenicity within the cutworm complex. Biol. Control 2004, 31, 372–379. [Google Scholar] [CrossRef]
- Wennmann, J.T.; Gueli Alletti, G.; Jehle, J.A. The genome sequence of Agrotis segetum nucleopolyhedrovirus B (AgseNPV-B) reveals a new baculovirus species within the Agrotis baculovirus complex. Virus Genes 2015, 50, 260–276. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.L.; Mowery, J.D.; Bauchan, G.R.; Theilmann, D.A.; Erlandson, M.A. The complete genome sequence of a second alphabaculovirus from the true armyworm, Mythimna unipuncta: Implications for baculovirus phylogeny and host specificity. Virus Genes 2019, 55, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Herniou, E.A.; Jehle, J.A. Baculovirus phylogeny and evolution. Curr. Drug Targets 2007, 8, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Guarino, L.A.; Mistretta, T.A.; Dong, W. Baculovirus lef-12 is not required for viral replication. J. Virol. 2002, 76, 12032–12043. [Google Scholar] [CrossRef] [PubMed]
- Lauzon, H.A.; Jamieson, P.B.; Krell, P.J.; Arif, B.M. Gene organization and sequencing of the Choristoneura fumiferana defective nucleopolyhedrovirus genome. J. Gen. Virol. 2005, 86, 945–961. [Google Scholar] [CrossRef]
- Oliveira, J.V.; Wolff, J.L.; Garcia-Maruniak, A.; Ribeiro, B.M.; de Castro, M.E.; de Souza, M.L.; Moscardi, F.; Maruniak, J.E.; Zanotto, P.M. Genome of the most widely used viral biopesticide: Anticarsia gemmatalis multiple nucleopolyhedrovirus. J. Gen. Virol. 2006, 87, 3233–3250. [Google Scholar] [CrossRef]
- Brito, A.F.; Braconi, C.T.; Weidmann, M.; Dilcher, M.; Alves, J.M.; Gruber, A.; Zanotto, P.M. The pangenome of the Anticarsia gemmatalis multiple nucleopolyhedrovirus (AgMNPV). Genome Biol. Evol. 2015, 8, 94–108. [Google Scholar] [CrossRef]
- Harrison, R.L. Concentration- and time-response characteristics of plaque isolates of Agrotis ipsilon multiple nucleopolyhedrovirus derived from a field isolate. J. Invertebr. Pathol. 2013, 112, 159–161. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fiducial Limits | Median | ||||||
|---|---|---|---|---|---|---|---|
| Virus | LC50a (OBs/mL) | Lower Limit | Upper Limit | Slope | LT50ab (hr p.i.) | 95% CL | % Mortality |
| ChinNPV#1 | 1.71 × 105 a | 9.97 × 104 | 2.90 × 105 | 1.8 ± 0.3 | 166.9 a | 164.5–169.3 | 76.7 |
| ChinNPV-460 | 1.87 × 105 a | 9.00 × 104 | 3.43 × 105 | 1.5 ± 0.3 | 91.3 b | 61.2–121.4 | 86.7 |
| AcMNPV | 1.40 × 106 b | 8.46 × 105 | 2.31 × 106 | 1.7 ± 0.3 | 77.6 b | 75.3–79.9 | 93.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harrison, R.L.; Rowley, D.L.; Popham, H.J.R. A Novel Alphabaculovirus from the Soybean Looper, Chrysodeixis includens, that Produces Tetrahedral Occlusion Bodies and Encodes Two Copies of he65. Viruses 2019, 11, 579. https://doi.org/10.3390/v11070579
Harrison RL, Rowley DL, Popham HJR. A Novel Alphabaculovirus from the Soybean Looper, Chrysodeixis includens, that Produces Tetrahedral Occlusion Bodies and Encodes Two Copies of he65. Viruses. 2019; 11(7):579. https://doi.org/10.3390/v11070579
Chicago/Turabian StyleHarrison, Robert L., Daniel L. Rowley, and Holly J. R. Popham. 2019. "A Novel Alphabaculovirus from the Soybean Looper, Chrysodeixis includens, that Produces Tetrahedral Occlusion Bodies and Encodes Two Copies of he65" Viruses 11, no. 7: 579. https://doi.org/10.3390/v11070579
APA StyleHarrison, R. L., Rowley, D. L., & Popham, H. J. R. (2019). A Novel Alphabaculovirus from the Soybean Looper, Chrysodeixis includens, that Produces Tetrahedral Occlusion Bodies and Encodes Two Copies of he65. Viruses, 11(7), 579. https://doi.org/10.3390/v11070579