BK Virus Replication in the Glomerular Vascular Unit: Implications for BK Virus Associated Nephropathy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. BK Virus Preparation, Cultivation, and Titration
2.3. BKV RNA Analysis
2.4. qRT-PCR Analysis of the Proinflammatory Cytokine Gene Expression
2.5. Immunofluorescence
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
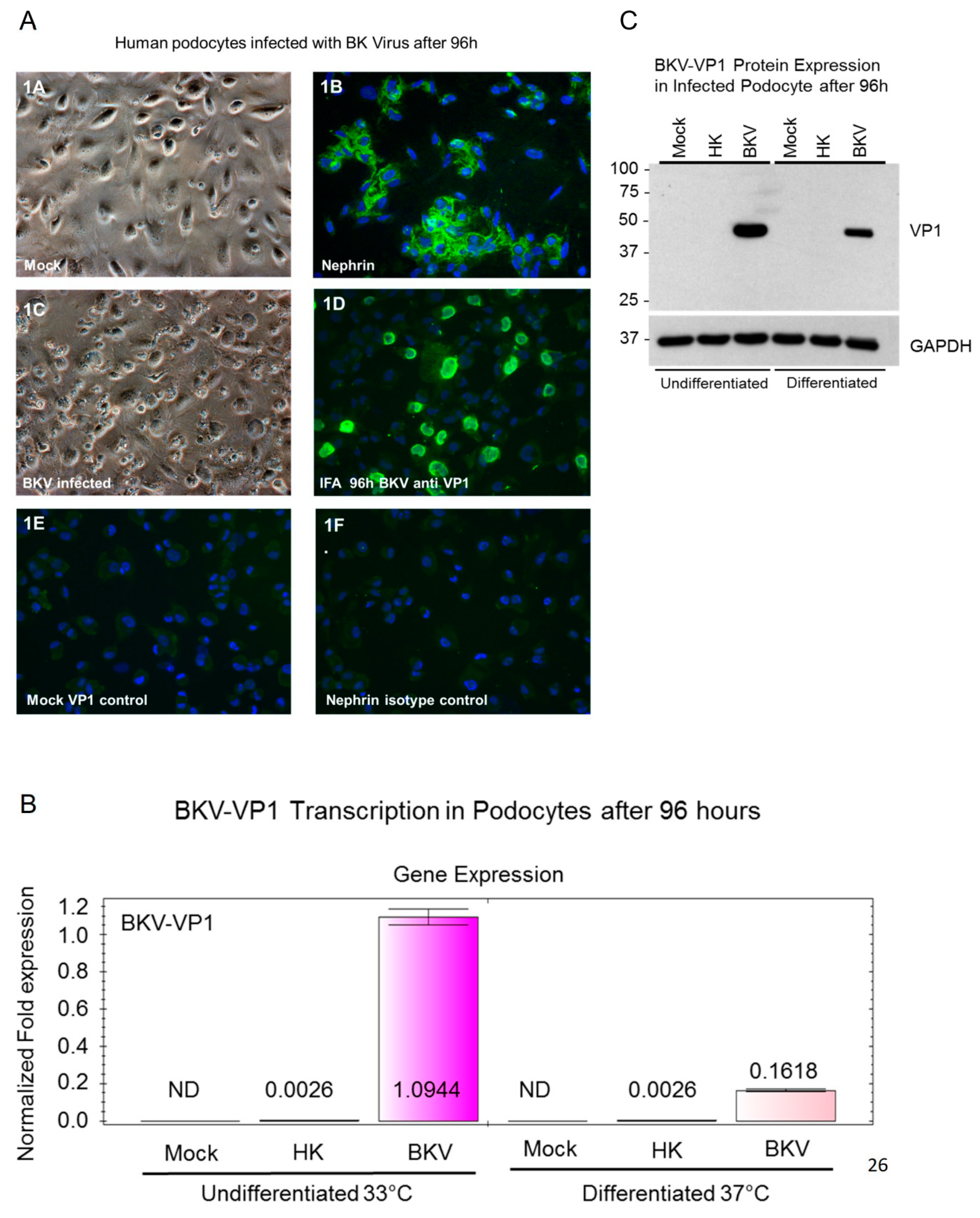
3.1. BKV Lytic Replication in Human Glomerular Podocytes
3.2. BKV Infected Undifferentiated Podocytes Express Higher Levels of VP1 Transcripts
3.3. VP1 Protein Expression in Infected Undifferentiated and Differentiated Podocytes
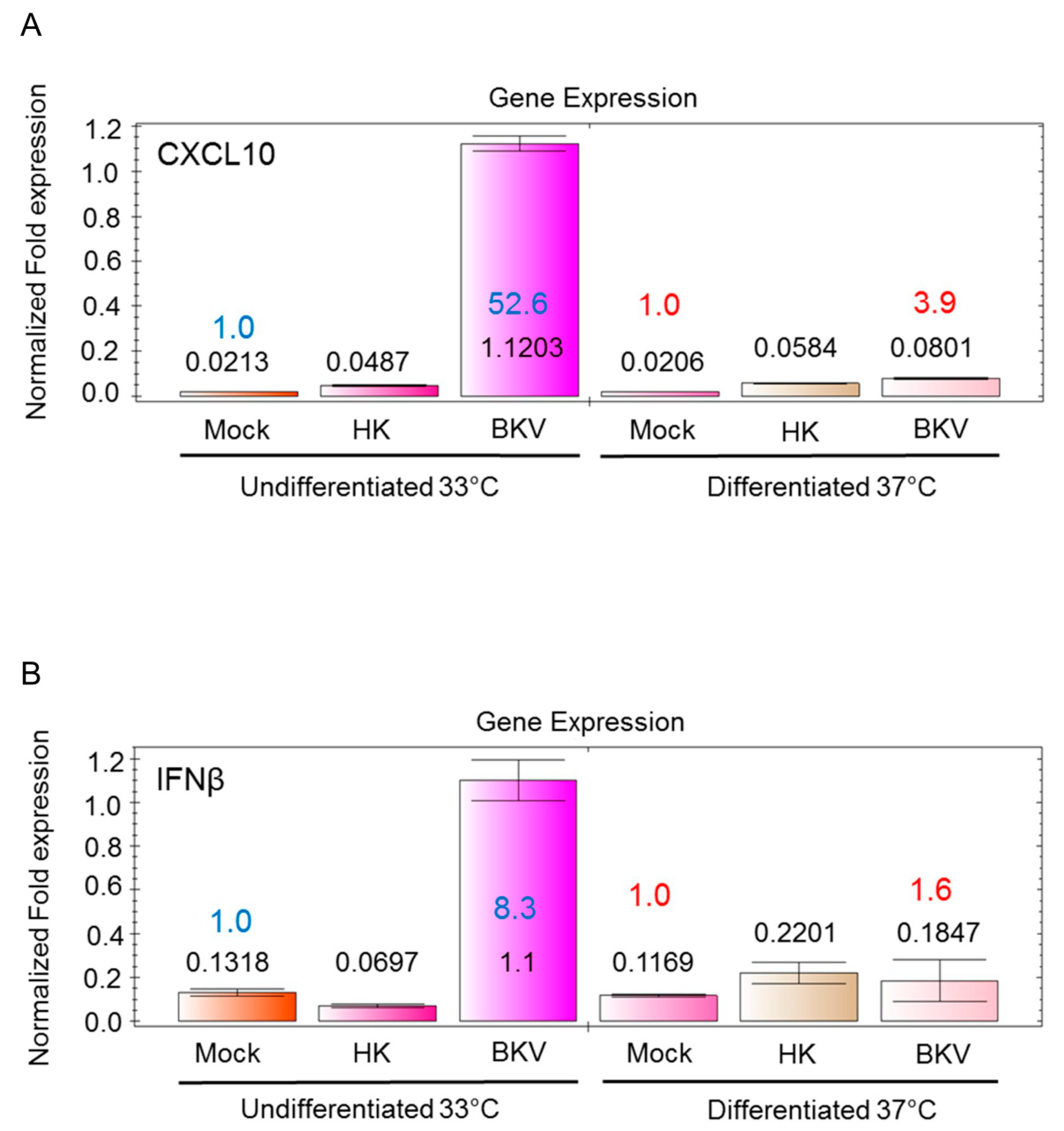
3.4. BKV Infection Induces Higher IFNβ and CXCL10 Gene Expression in Undifferentiated Podocytes
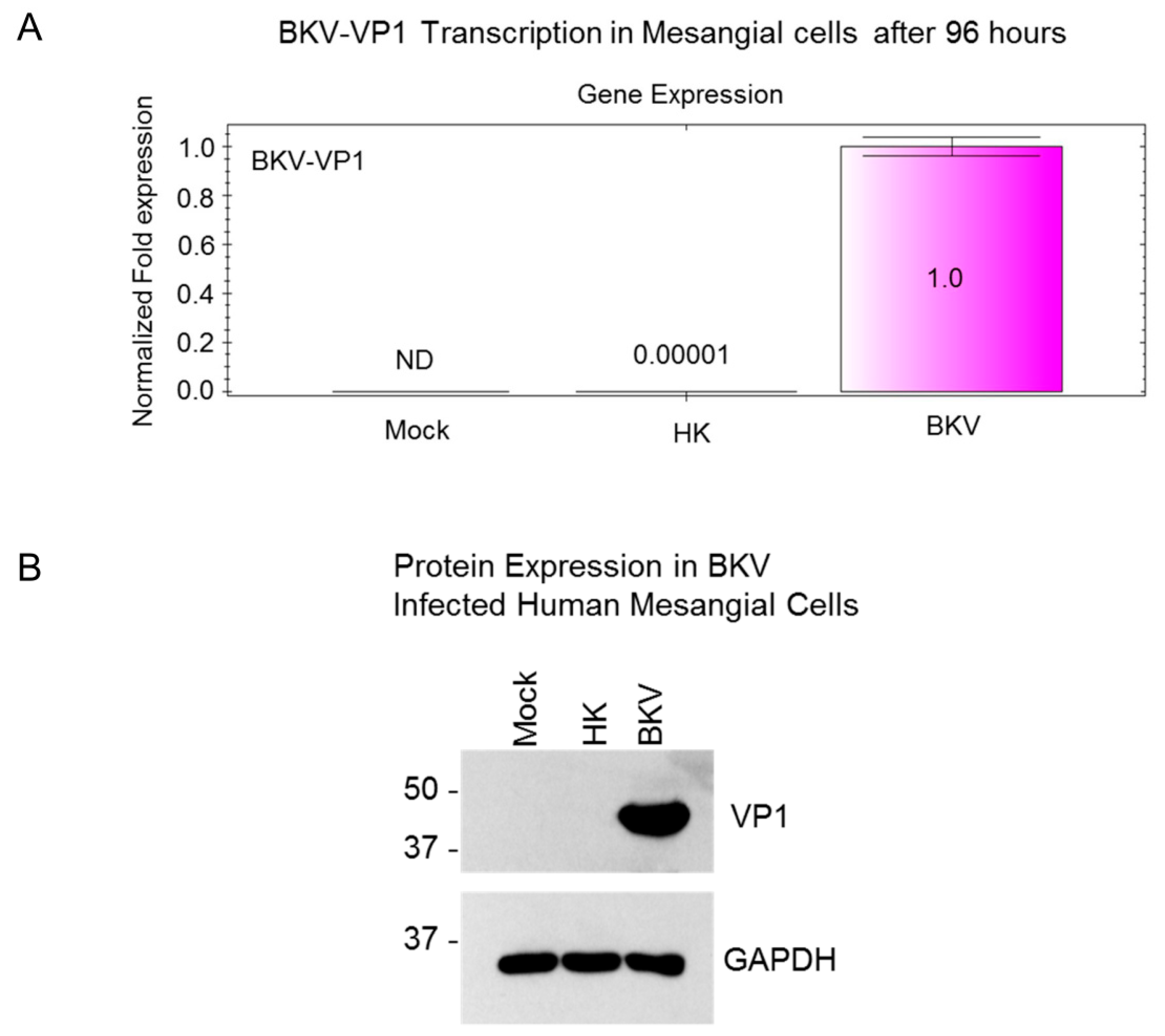
3.5. BKV Replication in Human Primary Renal Mesangial Cells
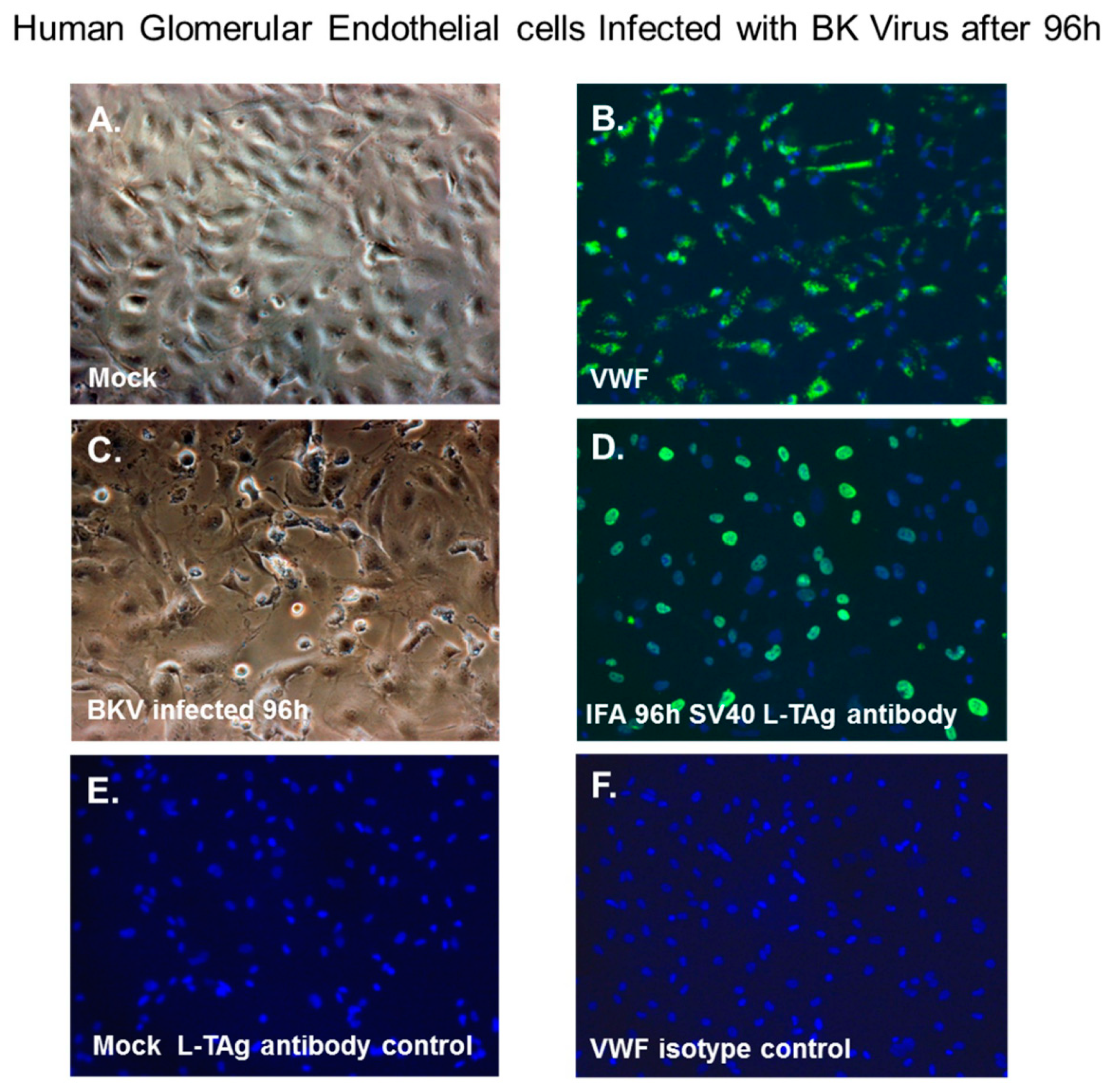
3.6. BKV Lytic Replication in Human Glomerular Endothelial Cells
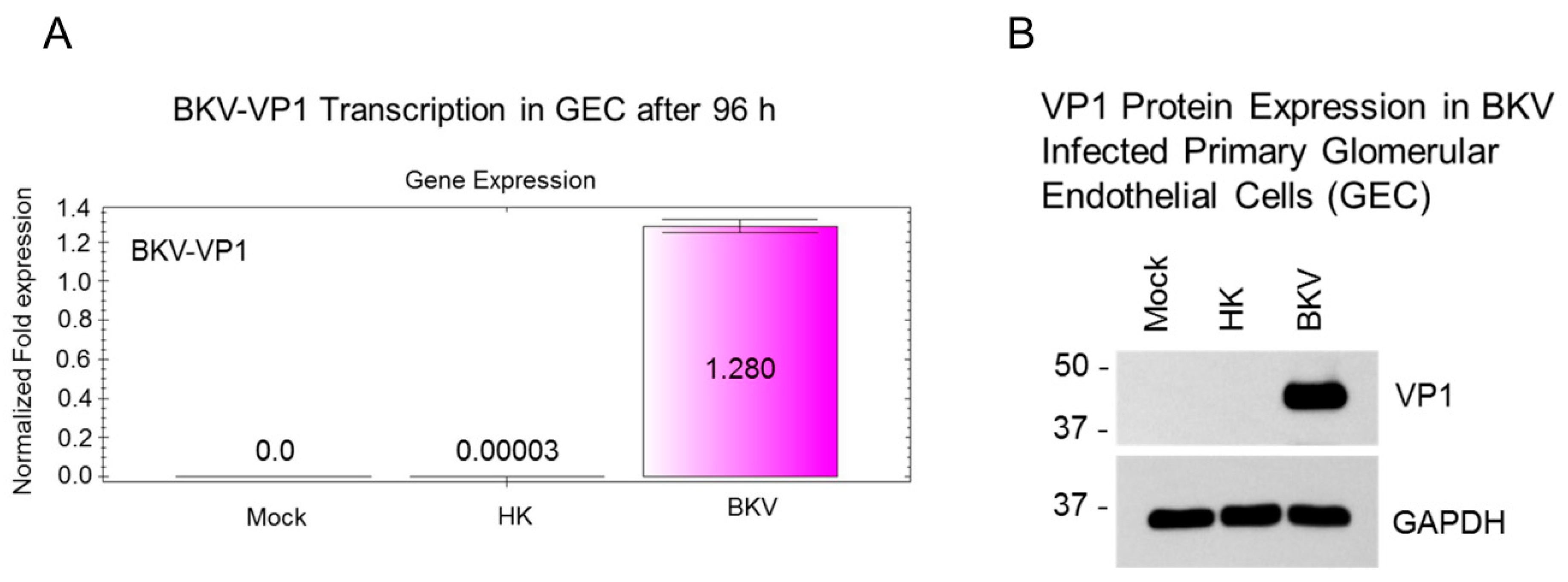
3.7. BKV Infected Glomerular Endothelial Cells Express High Levels of VP1 Transcripts and VP1 Protein
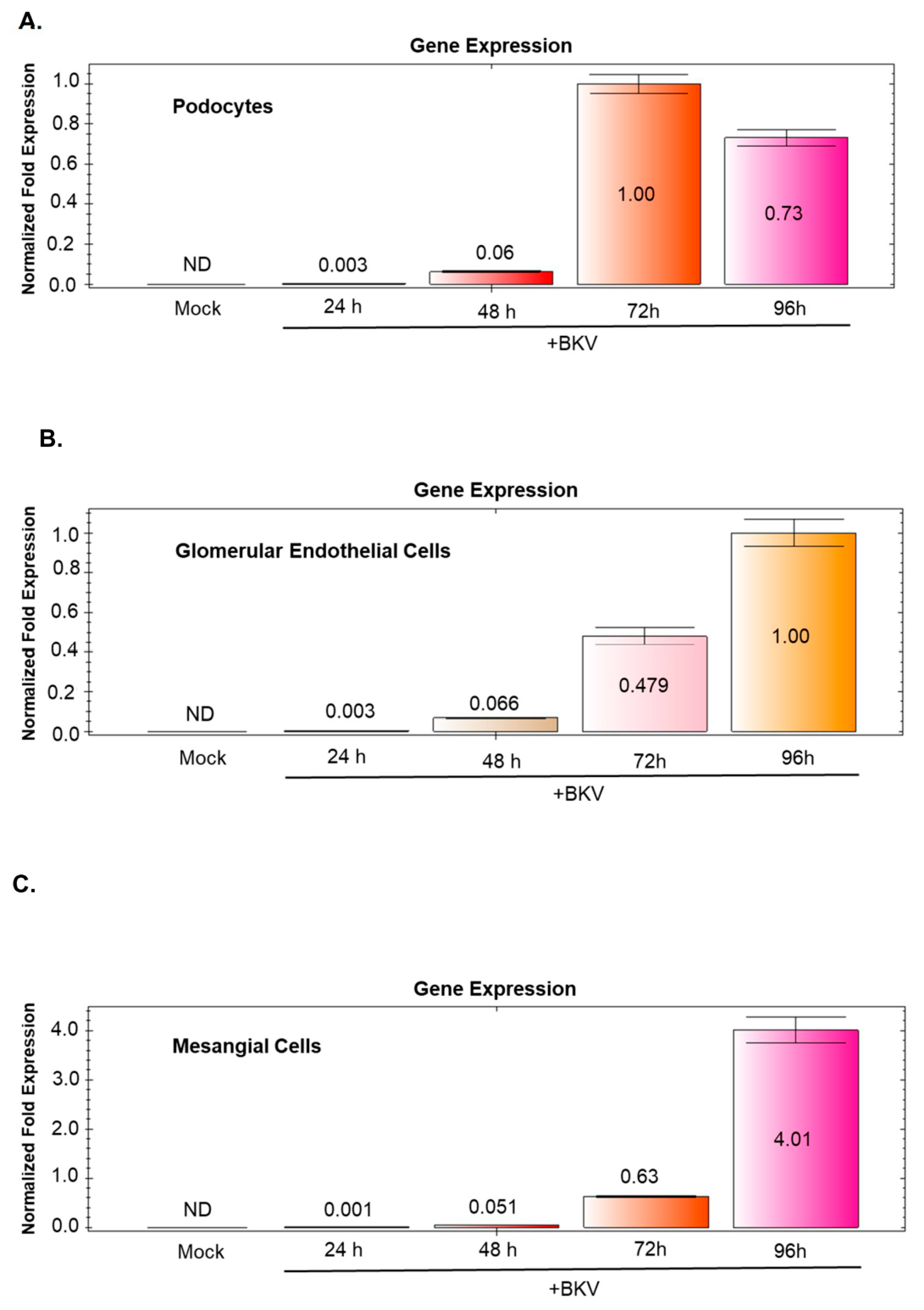
3.8. BKV-Induced VP1 Transcription Increases Over Time in BKV Infected GVU Cells
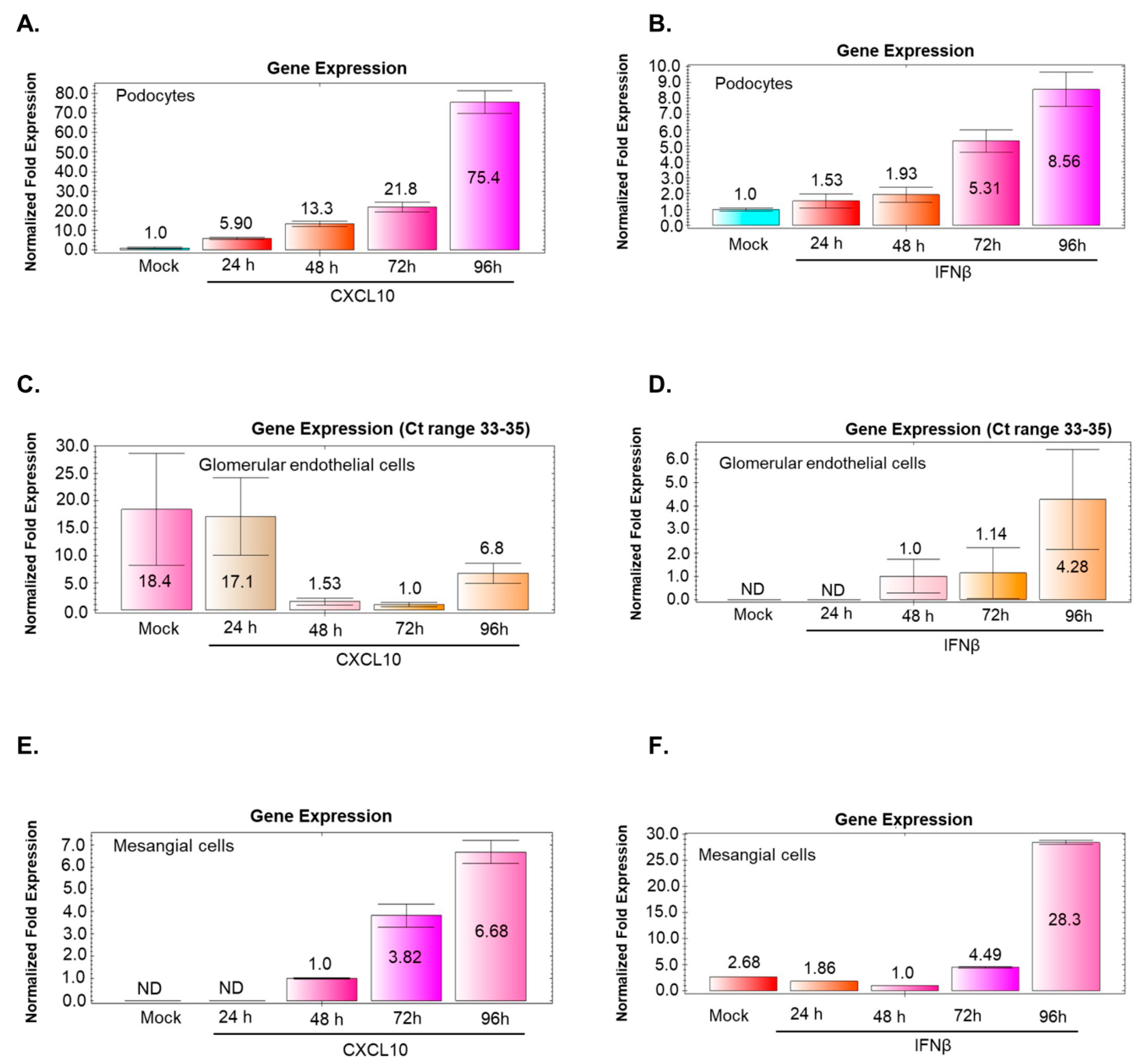
3.9. BKV Infection of Podocytes and Mesangial Cells Induces CXCL10 and IFNβ Gene Expression that Correlates with Increased Virus Replication
3.10. BKV Infection of Glomerular Endothelial Cells does not Noticeably Stimulate CXCL10 or IFNβ Gene Expression
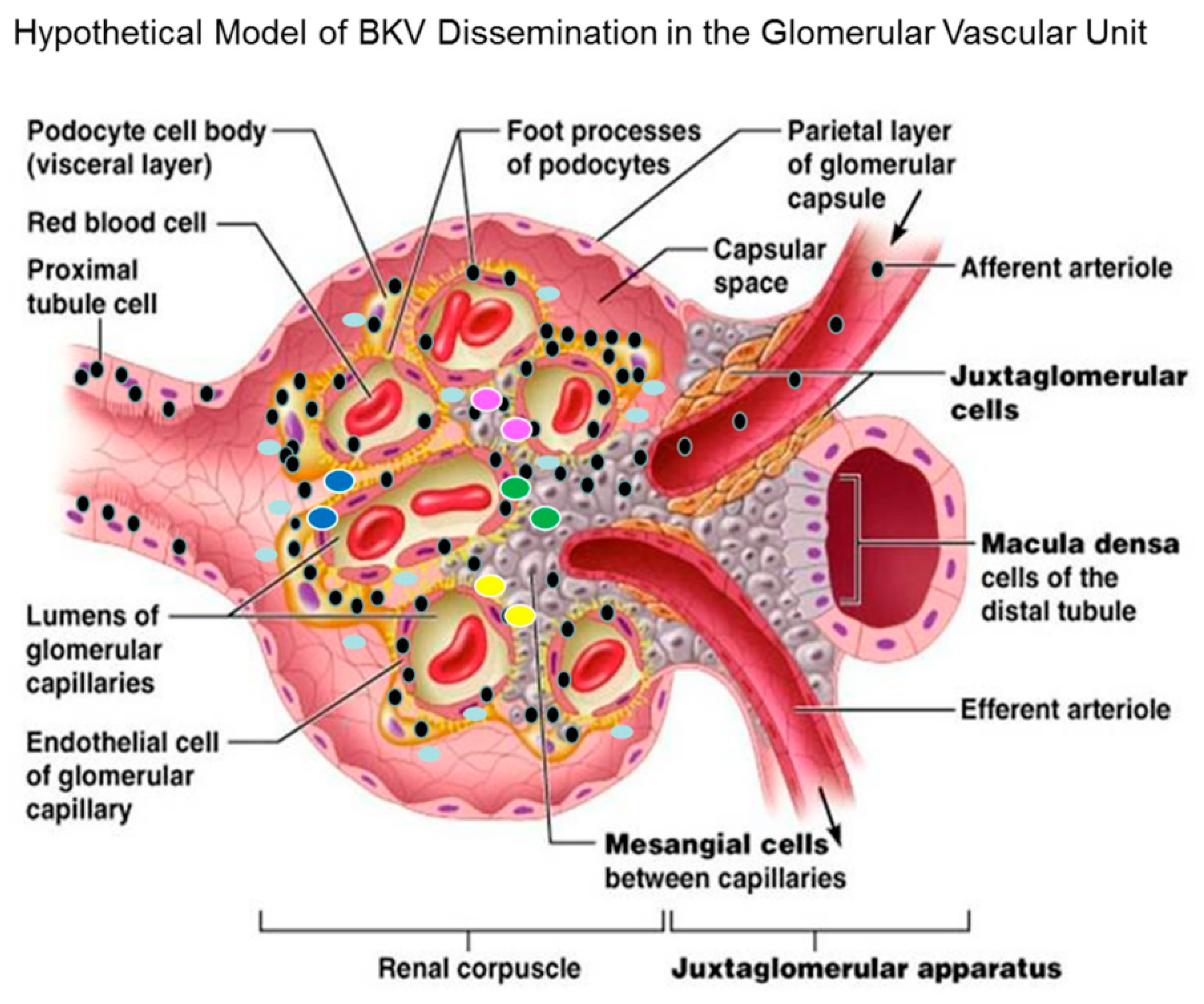
3.11. Hypothetical Model for BKV Dissemination in Glomerulus.
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gardner, S.D.; Field, A.M.; Coleman, D.V.; Hulme, B. New human papovavirus (B.K.) isolated from urine after renal transplantation. Lancet 1971, 297, 1253–1257. [Google Scholar] [CrossRef]
- Shah, K.V. Polyomaviruses. In Virology; Fields, B.N., Knipe, K.M., Howley, P.M., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 1990; pp. 2027–2043. [Google Scholar]
- Hirsch, H.H. BK virus: Opportunity makes a pathogen. Clin. Infect. Dis. 2005, 41, 354–360. [Google Scholar] [CrossRef]
- Ahsan, N.; Shah, K.V. Polyomaviruses and Human Diseases; Springer: New York, NY, USA, 2006. [Google Scholar]
- Siguier, M.; Sellier, P.; Bergmann, J.-F. BK-virus infections: A literature review. Med. Mal. Infect. 2012, 42, 181–187. [Google Scholar] [CrossRef]
- Nickeleit, V.; Singh, H.K. Polyomaviruses and disease: Is there more to know than viremia and viruria? Curr. Opin. Organ Transplant. 2015, 20, 348–358. [Google Scholar] [CrossRef]
- Teutsch, K.; Schweitzer, F.; Knops, E.; Kaiser, R.; Pfister, H.; Verheyen, J.; Göbel, H.; Clingöz, T.; Di Cristanziano, V. Early identification of renal transplant recipients with high risk of polyomavirus-associated nephropathy. Med. Microbiol. Immunol. 2015, 204, 657–664. [Google Scholar] [CrossRef]
- Bennett, J.E.; Dolin, R.; Blaser, M.J. Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases; Saunders: Philadelphia, PA, USA, 2015. [Google Scholar]
- Pinto, M.; Dobson, S. BK and JC virus: A review. J. Infect. 2014, 68, S2–S8. [Google Scholar] [CrossRef]
- Vago, L.; Cinque, P.; Sala, E.; Nebuloni, M.; Caldarelli, R.; Racca, S.; Ferrante, P.; Trabattoni, G.R.; Costanzi, G. JCV-DNA and BKV-DNA in the CNS tissue and CSF of AIDS patients and normal subjects. Study of 41 cases and review of the literature. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1996, 12, 139–146. [Google Scholar] [CrossRef]
- Lesprit, P.; Chaline-Lehmann, D.; Authier, F.-J.; Ponnelle, T.; Gray, F.; Levy, Y. BK virus encephalitis in a patient with AIDS and lymphoma. AIDS 2001, 15, 1196–1199. [Google Scholar] [CrossRef]
- Jiang, M.; Abend, J.R.; Johnson, S.F.; Imperiale, M.J. The role of polyomaviruses in human disease. Virology 2009, 384, 266–273. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, H.H.; Randhawa, P. AST Infectious Diseases Community of Practice. BK polyomavirus in solid organ transplantation. Am. J. Transplant. 2013, 13 (Suppl. 4), 179–188. [Google Scholar] [CrossRef]
- Dalianis, T.; Hirsch, H.H. Human polyomaviruses in disease and cancer. Virology 2013, 437, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Binet, I.; Nickeleit, V.; Hirsch, H.H.; Prince, O.; Dalquen, P.; Gudat, F.; Mihatsch, M.J.; Thiel, G. Polyomavirus disease under new immunosuppressive drugs: A cause of renal graft dysfunction and graft loss. Transplantation 1999, 67, 918–922. [Google Scholar] [CrossRef]
- Comoli, P.; Binggeli, S.; Ginevri, F.; Hirsch, H.H. Polyomavirus-associated nephropathy: Update on BK virus-specific immunity. Transplant. Infect. Dis. 2006, 8, 86–94. [Google Scholar] [CrossRef]
- Han, S.B.; Cho, B.; Kang, J.H. BK virus-associated hemorrhagic cystitis after pediatric stem cell transplantation. Korean J. Pediatr. 2014, 57, 514–519. [Google Scholar] [CrossRef] [Green Version]
- Kariminik, A.; Dabiri, S.; Yaghobi, R. Polyomavirus BK induces inflammation via up-regulation of CXCL10 at translation levels in renal transplant patients with nephropathy. Inflammation 2016, 39, 1514–1519. [Google Scholar] [CrossRef]
- Egli, A.; Binggeli, S.; Bodaghi, S.; Dumoulin, A.; Funk, G.A.; Khanna, N.; Leuenberger, D.; Gosert, R.; Hirsch, H.H. Cytomegalovirus and polyomavirus BK posttransplant. Nephrol. Dial. Transplant. 2007, 22 (Suppl. 8), viii72–viii82. [Google Scholar] [CrossRef] [Green Version]
- Burrows, N.R.; Li, Y.; Williams, D.E. Racial and ethnic differences in trends of end-stage renal disease: United States, 1995 to 2005. Adv. Chronic Kidney Dis. 2008, 15, 147–152. [Google Scholar] [CrossRef]
- Palmer Alves, T.; Lewis, J. Racial differences in chronic kidney disease (CKD) and end-stage renal disease (ESRD) in the United States: A social and economic dilemma. Clin. Nephrol. 2010, 74 (Suppl. 1), S72–S77. [Google Scholar]
- Regunathan-Shenk, R.; Hussain, F.N.; Ganda, A. Chronic kidney disease and end-stage renal disease in disadvantaged communities of North America: An investigational challenge to limit disease progression and cardiovascular risk. Clin. Nephrol. 2016, 86 (Suppl. 13), 37–40. [Google Scholar] [CrossRef]
- Albertus, P.; Morgenstern, H.; Robinson, B.; Saran, R. Risk of ESRD in the United States. Am. J. Kidney Dis. 2016, 68, 862–872. [Google Scholar] [CrossRef] [Green Version]
- Alcendor, D.J. Zika Virus Infection of the Human Glomerular Cells: Implications for Viral Reservoirs and Renal Pathogenesis. J. Infect. Dis. 2017, 216, 162–171. [Google Scholar] [CrossRef]
- Popik, W.; Correa, H.; Khatua, A.; Aronoff, D.M.; Alcendor, D.J. Mesangial cells, specialized renal Pericytes, and cytomegalovirus infectivity: Implications for HCMV pathology in the glomerular vascular unit and post-transplant renal disease. J. Transl. Sci. 2018, 5, 1–12. [Google Scholar] [CrossRef]
- Saleem, M.A.; O’Hare, M.J.; Reiser, J. A conditionally immortalized human podocyte cell line demonstrating nephrin and podocin expression. J. Am. Soc. Nephrol. 2002, 13, 630–638. [Google Scholar]
- Khatua, A.K.; Taylor, H.E.; Hildreth, J.E.K.; Popik, W. Non-productive HIV-1 infection of human glomerular and urinary podocytes. Virology 2010, 408, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Seif, I.; Khoury, G.; Dhar, R. The genome of human papovavirus BKV. Cell 1979, 18, 963–977. [Google Scholar] [CrossRef]
- Takemoto, K.K.; Mullarkey, M.F. Human papovavirus, BK strain: Biological studies including antigenic relationship to simian virus 40. J. Virol. 1973, 12, 625–631. [Google Scholar]
- Wilkerson, I.; Laban, J.; Mitchell, J.M.; Sheibani, N.; Alcendor, D.J. Retinal pericytes and cytomegalovirus infectivity: Implications for HCMV-induced retinopathy and congenital ocular disease. J. Neuroinflamm. 2015, 12, 2. [Google Scholar] [CrossRef]
- Dabiri, S.; Kariminik, A.; Kennedy, D. The role of CXCR3 and its ligands in renal transplant outcome. Eur. Cytokine Netw. 2016, 2, 34–40. [Google Scholar]
- Brealey, J.K. Ultrastructural observations in a case of BK virus nephropathy with viruses in glomerular subepithelial humps. Ultrastruct. Pathol. 2007, 31, 1–7. [Google Scholar] [CrossRef]
- Lee, J.Y.; Song, S.H.; Kim, Y.S.; Lim, B.J.; Kim, S.I.; Kim, M.S.; Jeong, H.J. Tubuloreticular inclusions in peritubular capillaries of renal allografts. Pathol. Res. Pract. 2017, 213, 1185–1190. [Google Scholar] [CrossRef]
- Assetta, B.; De Cecco, M.; O’Hara, B.; Altwood, W.J. JC polyomavirus infection of primary human renal epithelial cells is controlled by a type I IFN-induced response. MBio 2016, 7, e00903-16. [Google Scholar] [CrossRef]
- Jia, L.; Fu, W.; Jia, R. Identification of potential key protein interaction networks of BK virus nephropathy in patients receiving kidney transplantation. Sci. Rep. 2018, 8, 5017. [Google Scholar] [CrossRef]
- Hu, H.; Aizenstein, B.D.; Puchalski, A.; Burmania, J.A.; Hamawy, M.M.; Knechtle, S.J. Elevation of CXCR3-binding chemokines in urine indicates acute renal-allograft dysfunction. Am. J. Transplant. 2004, 4, 432–437. [Google Scholar] [CrossRef]
- Jackson, J.A.; Kim, E.J.; Begley, B.; Cheeseman, J.; Harden, T.; Perez, S.D.; Thomas, S.; Warshaw, B.; Kirk, A.D. Urinary chemokines CXCL9 and CXCL10 are noninvasive markers of renal allograft rejection and BK viral infection. Am. J. Transplant. 2011, 11, 2228–2234. [Google Scholar] [CrossRef]
- Hirt-Minkowski, P.; Amico, P.; Ho, J.; Gao, A.; Bestland, J.; Hopfer, H.; Steiger, J.; Dickenmann, M.; Burkhalter, F.; Rush, D.; et al. Detection of clinical and subclinical tubulo-interstitial inflammation by the urinary CXCL10 chemokine in a real-life setting. Am. J. Transplant. 2012, 12, 1811–1823. [Google Scholar] [CrossRef]
- Schaub, S.; Nickerson, P.; Rush, D.; Mayr, M.; Hess, C.; Golian, M.; Stefura, W.; HayGlass, K. Urinary CXCL9 and CXCL10 levels correlate with the extent of subclinical tubulitis. Am. J. Transplant. 2009, 9, 1347–1353. [Google Scholar] [CrossRef]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popik, W.; Khatua, A.K.; Fabre, N.F.; Hildreth, J.E.K.; Alcendor, D.J. BK Virus Replication in the Glomerular Vascular Unit: Implications for BK Virus Associated Nephropathy. Viruses 2019, 11, 583. https://doi.org/10.3390/v11070583
Popik W, Khatua AK, Fabre NF, Hildreth JEK, Alcendor DJ. BK Virus Replication in the Glomerular Vascular Unit: Implications for BK Virus Associated Nephropathy. Viruses. 2019; 11(7):583. https://doi.org/10.3390/v11070583
Chicago/Turabian StylePopik, Waldemar, Atanu K. Khatua, Noyna F. Fabre, James E. K. Hildreth, and Donald J. Alcendor. 2019. "BK Virus Replication in the Glomerular Vascular Unit: Implications for BK Virus Associated Nephropathy" Viruses 11, no. 7: 583. https://doi.org/10.3390/v11070583
APA StylePopik, W., Khatua, A. K., Fabre, N. F., Hildreth, J. E. K., & Alcendor, D. J. (2019). BK Virus Replication in the Glomerular Vascular Unit: Implications for BK Virus Associated Nephropathy. Viruses, 11(7), 583. https://doi.org/10.3390/v11070583