Bioluminescent Ross River Virus Allows Live Monitoring of Acute and Long-Term Alphaviral Infection by In Vivo Imaging

 , , ,
, , ,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
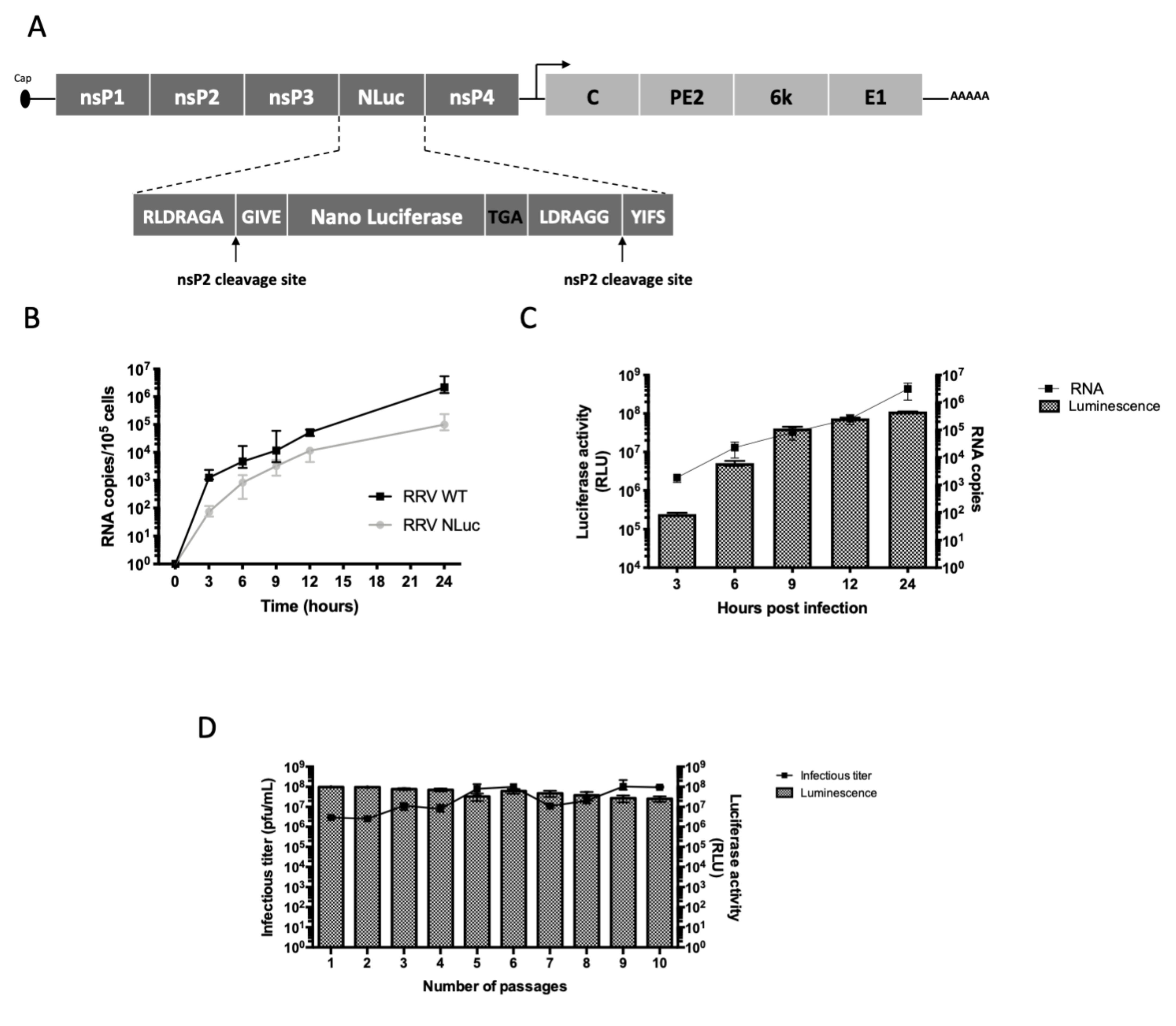
2.1. Design of the Recombinant RRV-NLuc
2.2. Production and Titration of the Viruses
2.3. In Vitro Replication Kinetics of the Recombinant and Parental Viruses
2.4. In Vitro Stability of the Recombinant Virus
2.5. Luciferase Activity Quantification
2.6. Viral RNA Extraction and Quantification
2.7. In Vivo Stability of the Recombinant Virus
2.8. Ethics Statement
2.9. Mouse Experiments
2.10. Tissue Collection for Ex Vivo Experiments
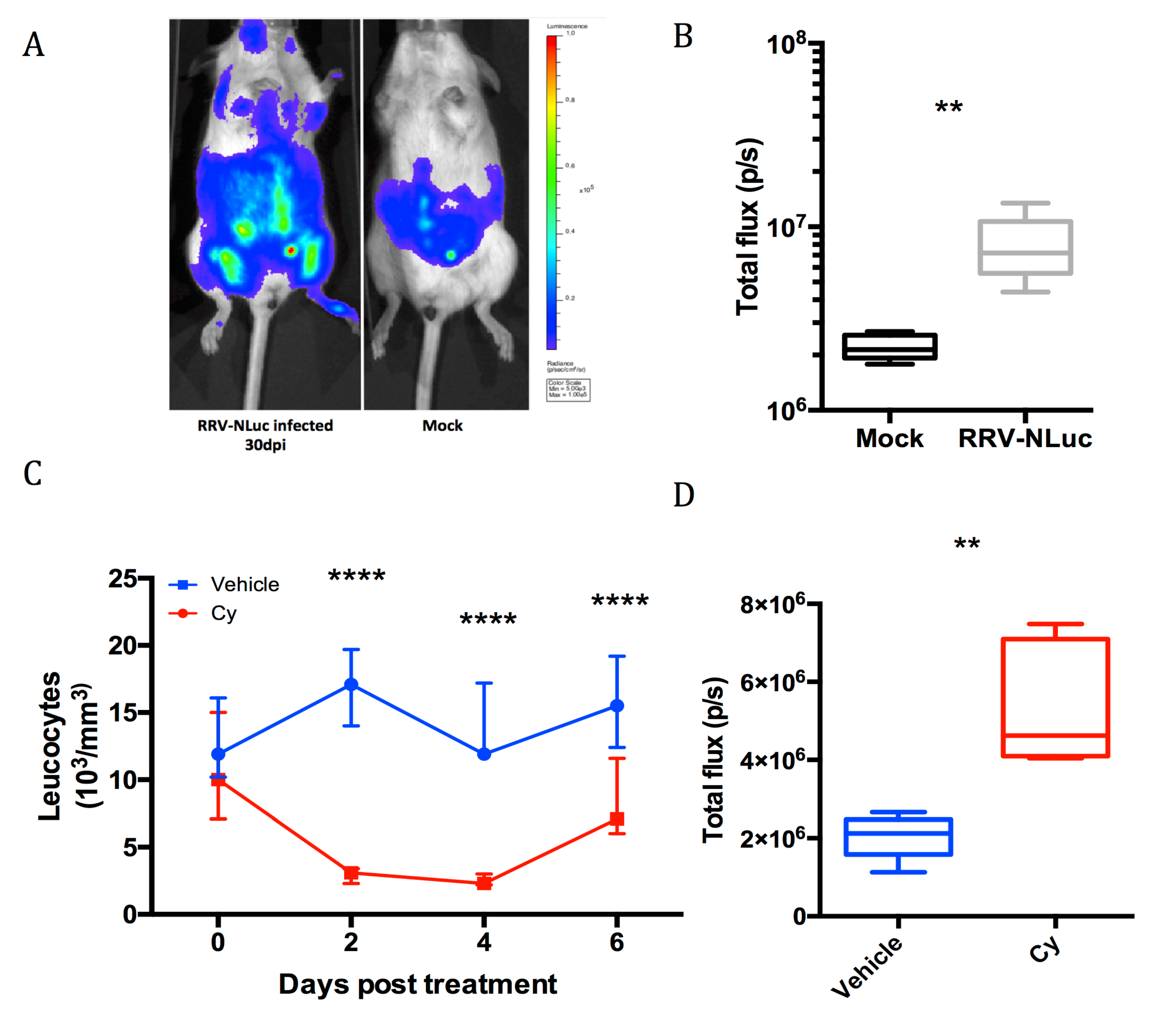
2.11. Cyclophosphamide Treatment
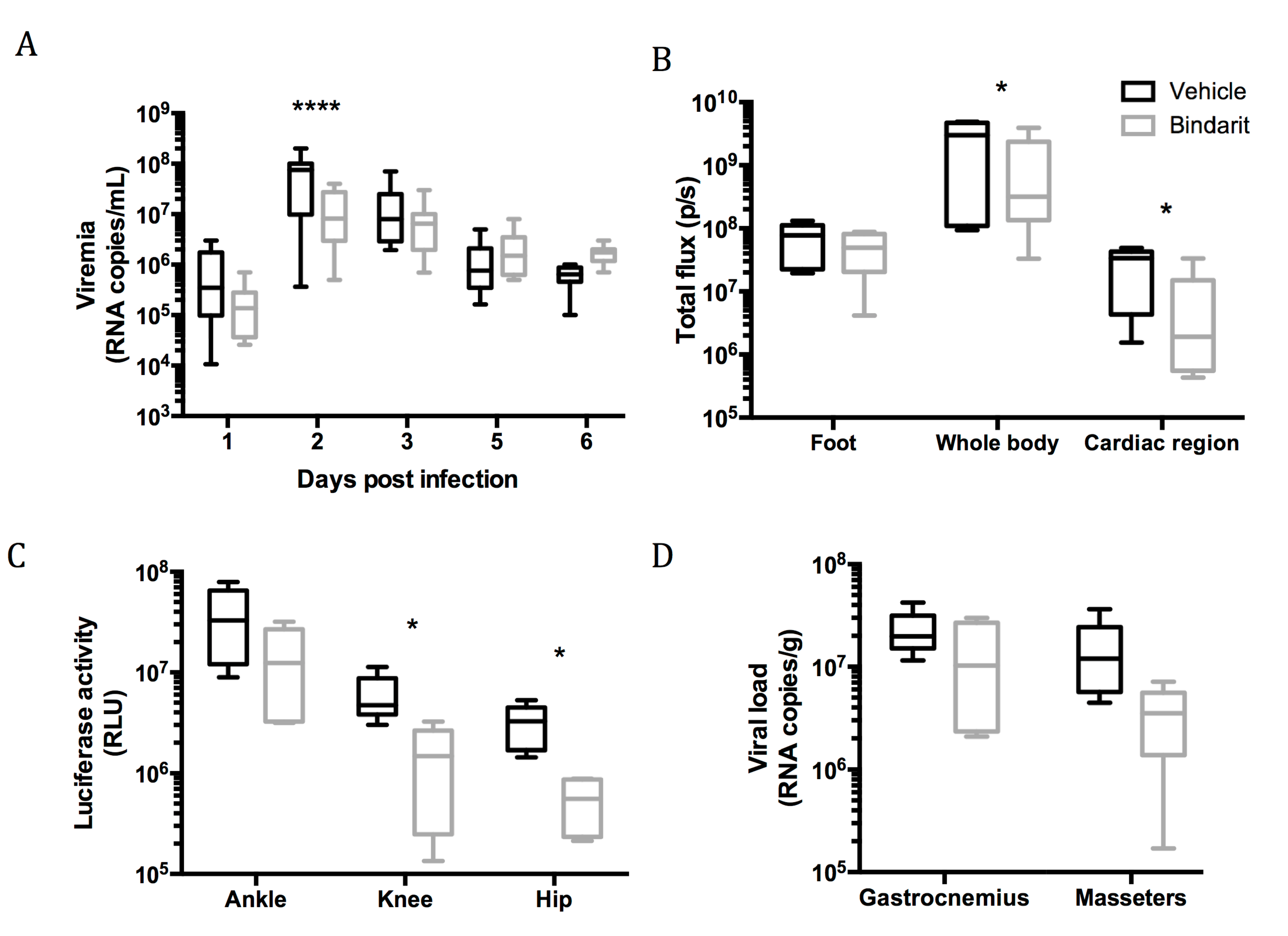
2.12. Bindarit Treatment
2.13. Bioluminescence Imaging
2.14. Statistical Analyses
3. Results
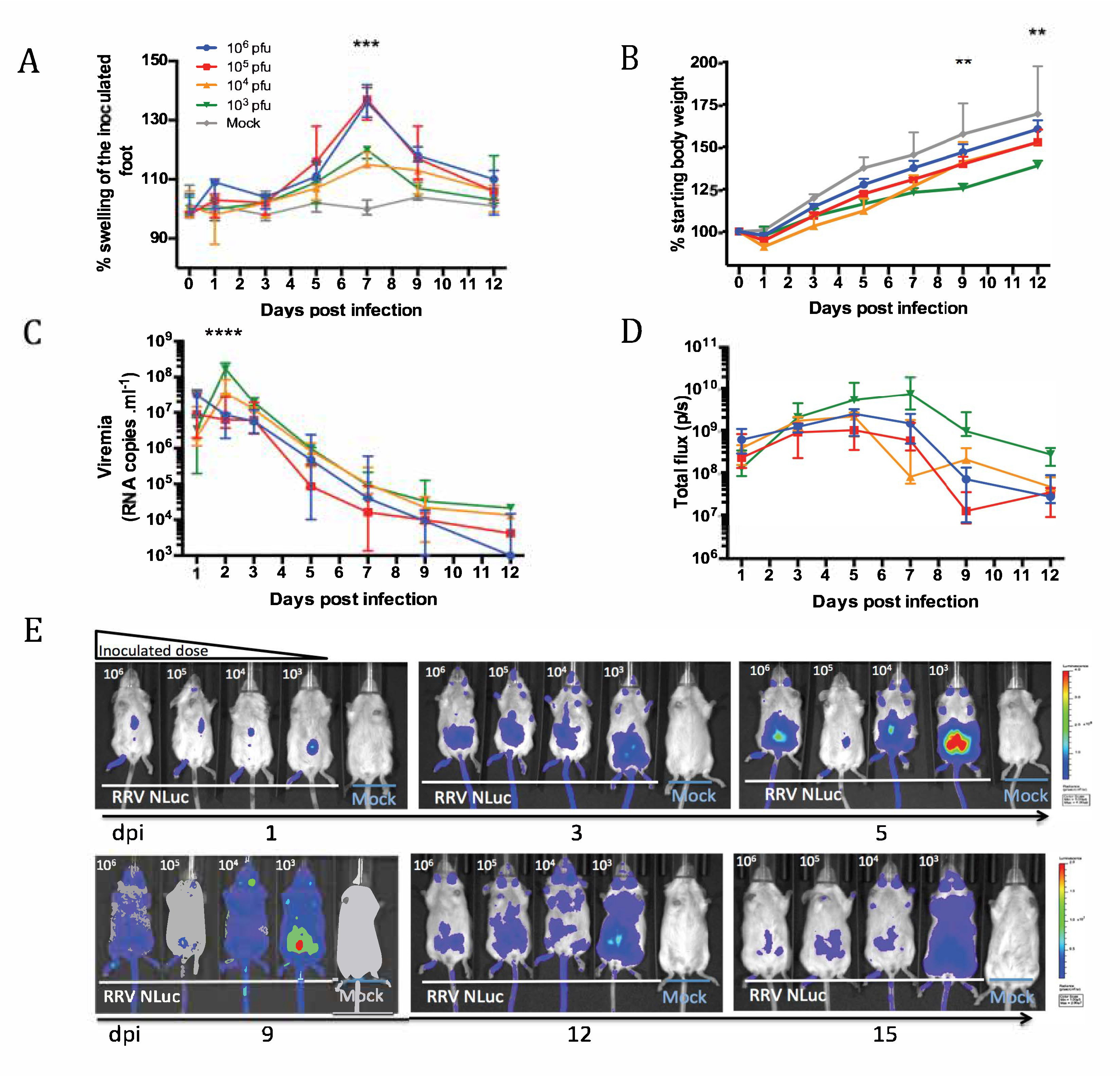
3.1. Design of Recombinant RRV-NLuc and In Vivo Infections
3.2. Dynamics of the Acute Phase of Infection with RRV-NLuc
3.3. Role of Macrophages During Viral Dissemination in the Acute Phase
3.4. Investigation of Long-Term Viral Replication
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Strauss, J.H.; Strauss, E.G. The alphaviruses: Gene expression, replication, and evolution. Microbiol. Mol. Biol. Rev. 1994, 58, 491–562. [Google Scholar]
- Suhrbier, A.; Jaffar-Bandjee, M.-C.; Gasque, P. Arthritogenic alphaviruses—an overview. Nat. Rev. Rheumatol. 2012, 8, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Harley, D.; Sleigh, A.; Ritchie, S. Ross River Virus Transmission, Infection, and Disease: A Cross-Disciplinary Review. Clin. Microbiol. Rev. 2001, 14, 909–932. [Google Scholar] [CrossRef] [PubMed]
- Department of Health Human Services. Australia Surveillance of Notifiable Conditions in Victoria; Department of Health Human Services: Melbourne, VIC, Australia, 2017.
- Weaver, S.C.; Forrester, N.L. Chikungunya: Evolutionary history and recent epidemic spread. Antivir. Res. 2015, 120, 32–39. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, C.N.D.; Dupont-Rouzeyrol, M.; Sam, I.C.; Roques, P. Global spread of chikungunya virus: A lesson for Aedes-transmitted arboviruses? CAB Rev. 2016, 11, 1–12. [Google Scholar] [CrossRef]
- Schwartz, O.; Albert, M.L. Biology and pathogenesis of chikungunya virus. Nat. Rev. Microbiol. 2010, 8, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Schilte, C.; Staikovsky, F.; Couderc, T.; Madec, Y.; Carpentier, F.; Kassab, S.; Albert, M.L.; Lecuit, M.; Michault, A. Chikungunya Virus-associated Long-term Arthralgia: A 36-month Prospective Longitudinal Study. PLOS Neglected. Trop. Dis. 2013, 7, e2137. [Google Scholar] [CrossRef]
- Mims, C.A.; Murphy, F.A.; Taylor, W.P.; Marshall, I.D. Pathogenesis of Ross River virus infection in mice. I. Ependymal infection, cortical thinning, and hydrocephalus. J. Infect. Dis. 1973, 127, 121–128. [Google Scholar] [CrossRef]
- Murphy, F.A.; Taylor, W.P.; Mims, C.A.; Marshall, I.D. Pathogenesis of Ross River virus infection in mice. II. Muscle, heart, and brown fat lesions. J. Infect. Dis. 1973, 127, 129–138. [Google Scholar] [CrossRef]
- Morrison, T.E.; Whitmore, A.C.; Shabman, R.S.; Lidbury, B.A.; Mahalingam, S.; Heise, M.T. Characterization of Ross River Virus Tropism and Virus-Induced Inflammation in a Mouse Model of Viral Arthritis and Myositis. J. Virol. 2006, 80, 737–749. [Google Scholar] [CrossRef] [Green Version]
- Gardner, J.; Anraku, I.; Le, T.T.; Larcher, T.; Major, L.; Roques, P.; Schroder, W.A.; Higgs, S.; Suhrbier, A. Chikungunya Virus Arthritis in Adult Wild-Type Mice. J. Virol. 2010, 84, 8021–8032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labadie, K.; Larcher, T.; Joubert, C.; Mannioui, A.; Delache, B.; Brochard, P.; Guigand, L.; Dubreil, L.; Lebon, P.; Verrier, B.; et al. Chikungunya disease in nonhuman primates involves long-term viral persistence in macrophages. J. Clin. Invest. 2010, 120, 894–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, S.M.; McGregor, A. A bright future for bioluminescent imaging in viral research. Future Virol. 2015, 10, 169–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heaton, N.S.; Leyva-Grado, V.H.; Tan, G.S.; Eggink, D.; Hai, R.; Palese, P. In Vivo Bioluminescent Imaging of Influenza A Virus Infection and Characterization of Novel Cross-Protective Monoclonal Antibodies. J. Virol. 2013, 87, 8272–8281. [Google Scholar] [CrossRef]
- England, C.G.; Ehlerding, E.B.; Cai, W. NanoLuc: A Small Luciferase is Brightening up the Field of Bioluminescence. Bioconjug. Chem. 2016, 27, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.H.; Griffin, D.E. Luciferase Imaging of a Neurotropic Viral Infection in Intact Animals. J. Virol. 2003, 77, 5333–5338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.T.; Stauft, C.B.; Aboellail, T.A.; Toth, A.M.; Jarvis, D.L.; Powers, A.M.; Olson, K.E. Bioluminescent Imaging and Histopathologic Characterization of WEEV Neuroinvasion in Outbred CD-1 Mice. PLoS ONE 2013, 8, e53462. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.T.; Rico, A.B.; Stauft, C.B.; Hammond, S.L.; Aboellail, T.A.; Tjalkens, R.B.; Olson, K.E. Entry Sites of Venezuelan and Western Equine Encephalitis Viruses in the Mouse Central Nervous System following Peripheral Infection. J. Virol. 2016, 90, 5785–5796. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, S.A.; Nuckols, J.; McGee, C.E.; Huang, Y.-J.S.; Vanlandingham, D.L.; Tesh, R.B.; Higgs, S. In Vivo Imaging of Chikungunya Virus in Mice and Aedes Mosquitoes Using a Renilla Luciferase Clone. Vector Borne Zoonotic Dis. 2011, 11, 1471–1477. [Google Scholar] [CrossRef]
- Teo, T.-H.; Lum, F.-M.; Claser, C.; Lulla, V.; Lulla, A.; Merits, A.; Rénia, L.; Ng, L.F.P. A Pathogenic Role for CD4+ T Cells during Chikungunya Virus Infection in Mice. J. Immunol. 2013, 190, 259–269. [Google Scholar] [CrossRef]
- Sun, C.; Gardner, C.L.; Watson, A.M.; Ryman, K.D.; Klimstra, W.B. Stable, High-Level Expression of Reporter Proteins from Improved Alphavirus Expression Vectors To Track Replication and Dissemination during Encephalitic and Arthritogenic Disease. J. Virol. 2014, 88, 2035–2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaton, B.T.; Hapel, A.J. Persistent noncytolytic togavirus infection of primary mouse muscle cells. Virology 1976, 72, 266–271. [Google Scholar] [CrossRef]
- Journeaux, S.F.; Brown, W.G.; Aaskov, J.G. Prolonged Infection of Human Synovial Cells with Ross River Virus. J. Gen. Virol. 1987, 68, 3165–3169. [Google Scholar] [CrossRef] [PubMed]
- Way, S.J.R.; Lidbury, B.A.; Banyer, J.L. Persistent Ross River Virus Infection of Murine Macrophages: An in Vitro Model for the Study of Viral Relapse and Immune Modulation during Long-Term Infection. Virology 2002, 301, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Ozden, S.; Huerre, M.; Riviere, J.-P.; Coffey, L.L.; Afonso, P.V.; Mouly, V.; de Monredon, J.; Roger, J.-C.; Amrani, M.E.; Yvin, J.-L.; et al. Human Muscle Satellite Cells as Targets of Chikungunya Virus Infection. PLoS ONE 2007, 2, e527. [Google Scholar] [CrossRef]
- Hoarau, J.-J.; Bandjee, M.-C.J.; Trotot, P.K.; Das, T.; Li-Pat-Yuen, G.; Dassa, B.; Denizot, M.; Guichard, E.; Ribera, A.; Henni, T.; et al. Persistent Chronic Inflammation and Infection by Chikungunya Arthritogenic Alphavirus in Spite of a Robust Host Immune Response. J. Immunol. 2010, 184, 5914–5927. [Google Scholar] [CrossRef] [Green Version]
- McInnes, I.B.; Schett, G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef]
- Ryman, K.D.; Klimstra, W.B. Closing the gap between viral and noninfectious arthritis. Proc. Natl. Acad. Sci. USA 2014, 111, 5767–5768. [Google Scholar] [CrossRef] [Green Version]
- Mora, E.; Guglielmotti, A.; Biondi, G.; Sassone-Corsi, P. Bindarit. Cell Cycle 2012, 11, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, R.J.; Niesters, H.G.; Hong, Z.; Strauss, J.H. Infectious RNA transcripts from Ross River virus cDNA clones and the construction and characterization of defined chimeras with Sindbis virus. Virology 1991, 182, 430–441. [Google Scholar] [CrossRef]
- Tamberg, N.; Lulla, V.; Fragkoudis, R.; Lulla, A.; Fazakerley, J.K.; Merits, A. Insertion of EGFP into the replicase gene of Semliki Forest virus results in a novel, genetically stable marker virus. J. Gen. Virol. 2007, 88, 1225–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henrik Gad, H.; Paulous, S.; Belarbi, E.; Diancourt, L.; Drosten, C.; Kümmerer, B.M.; Plate, A.E.; Caro, V.; Desprès, P. The E2-E166K substitution restores Chikungunya virus growth in OAS3 expressing cells by acting on viral entry. Virology 2012, 434, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafa, H.H.; Vogel, P.; Srinivasan, A.; Russell, C.J. Non-invasive Imaging of Sendai Virus Infection in Pharmacologically Immunocompromised Mice: NK and T Cells, but not Neutrophils, Promote Viral Clearance after Therapy with Cyclophosphamide and Dexamethasone. PLOS Pathog. 2016, 12, e1005875. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Foo, S.-S.; Taylor, A.; Lulla, A.; Merits, A.; Hueston, L.; Forwood, M.R.; Walsh, N.C.; Sims, N.A.; Herrero, L.J.; et al. Bindarit, an Inhibitor of Monocyte Chemotactic Protein Synthesis, Protects against Bone Loss Induced by Chikungunya Virus Infection. J. Virol. 2015, 89, 581–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seay, A.R.; Griffin, D.E.; Johnson, R.T. Experimental viral polymyositis: Age dependency and immune responses to Ross River virus infection in mice. Neurology 1981, 31, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Herrero, L.J.; Nelson, M.; Srikiatkhachorn, A.; Gu, R.; Anantapreecha, S.; Fingerle-Rowson, G.; Bucala, R.; Morand, E.; Santos, L.L.; Mahalingam, S. Critical role for macrophage migration inhibitory factor (MIF) in Ross River virus-induced arthritis and myositis. PNAS 2011, 108, 12048–12053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, V.; Moser, L.A.; Poole, D.S.; Mehle, A. Highly Sensitive Real-Time In Vivo Imaging of an Influenza Reporter Virus Reveals Dynamics of Replication and Spread. J. Virol. 2013, 87, 13321–13329. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.; Wu, T.-T.; Tong, L.M.; Kim, K.S.; Martinez-Guzman, D.; Colantonio, A.D.; Uittenbogaart, C.H.; Sun, R. Persistent Gammaherpesvirus Replication and Dynamic Interaction with the Host In Vivo. J. Virol. 2008, 82, 12498–12509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sourisseau, M.; Schilte, C.; Casartelli, N.; Trouillet, C.; Guivel-Benhassine, F.; Rudnicka, D.; Sol-Foulon, N.; Roux, K.L.; Prevost, M.-C.; Fsihi, H.; et al. Characterization of Reemerging Chikungunya Virus. PLOS Pathog. 2007, 3, e89. [Google Scholar] [CrossRef] [PubMed]
- Clarris, B.J.; Doherty, R.L.; Fraser, J.R.; French, E.L.; Muirden, K.D. Epidemic polyarthritis: A cytological, virological and immunochemical study. Aust N Z J. Med. 1975, 5, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.R.; Cunningham, A.L.; Clarris, B.J.; Aaskov, J.G.; Leach, R. Cytology of synovial effusions in epidemic polyarthritis. Aust. N. Z. J. Med. 1981, 11, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Rulli, N.E.; Guglielmotti, A.; Mangano, G.; Rolph, M.S.; Apicella, C.; Zaid, A.; Suhrbier, A.; Mahalingam, S. Amelioration of alphavirus-induced arthritis and myositis in a mouse model by treatment with bindarit, an inhibitor of monocyte chemotactic proteins. Arthritis Rheum. 2009, 60, 2513–2523. [Google Scholar] [CrossRef] [PubMed]
- Rulli, N.E.; Rolph, M.S.; Srikiatkhachorn, A.; Anantapreecha, S.; Guglielmotti, A.; Mahalingam, S. Protection from arthritis and myositis in a mouse model of acute chikungunya virus disease by bindarit, an inhibitor of monocyte chemotactic protein-1 synthesis. J. Infect. Dis. 2011, 204, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Soden, M.; Vasudevan, H.; Roberts, B.; Coelen, R.; Hamlin, G.; Vasudevan, S.; Brooy, J.L. Detection of viral ribonucleic acid and histologic analysis of inflamed synovium in Ross River virus infection. Arthritis & Rheum. 2000, 43, 365–369. [Google Scholar]
- Rouse, B.T.; Sehrawat, S. Immunity and immunopathology to viruses: What decides the outcome? Nat. Rev. Immunol. 2010, 10, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; Carrara, A.-S.; Aguilar, P.V.; Weaver, S.C. Evaluation of methods to assess transmission potential of Venezuelan equine encephalitis virus by mosquitoes and estimation of mosquito saliva titers. Am. J. Trop. Med. Hyg. 2005, 73, 33–39. [Google Scholar] [CrossRef]
- Hawman, D.W.; Stoermer, K.A.; Montgomery, S.A.; Pal, P.; Oko, L.; Diamond, M.S.; Morrison, T.E. Chronic Joint Disease Caused by Persistent Chikungunya Virus Infection Is Controlled by the Adaptive Immune Response. J. Virol. 2013, 87, 13878–13888. [Google Scholar] [CrossRef] [Green Version]
- Hawman, D.W.; Fox, J.M.; Ashbrook, A.W.; May, N.A.; Schroeder, K.M.S.; Torres, R.M.; Crowe, J.E.; Dermody, T.S.; Diamond, M.S.; Morrison, T.E. Pathogenic Chikungunya Virus Evades B Cell Responses to Establish Persistence. Cell Rep. 2016, 16, 1326–1338. [Google Scholar] [CrossRef] [Green Version]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belarbi, E.; Legros, V.; Basset, J.; Desprès, P.; Roques, P.; Choumet, V. Bioluminescent Ross River Virus Allows Live Monitoring of Acute and Long-Term Alphaviral Infection by In Vivo Imaging. Viruses 2019, 11, 584. https://doi.org/10.3390/v11070584
Belarbi E, Legros V, Basset J, Desprès P, Roques P, Choumet V. Bioluminescent Ross River Virus Allows Live Monitoring of Acute and Long-Term Alphaviral Infection by In Vivo Imaging. Viruses. 2019; 11(7):584. https://doi.org/10.3390/v11070584
Chicago/Turabian StyleBelarbi, Essia, Vincent Legros, Justine Basset, Philippe Desprès, Pierre Roques, and Valérie Choumet. 2019. "Bioluminescent Ross River Virus Allows Live Monitoring of Acute and Long-Term Alphaviral Infection by In Vivo Imaging" Viruses 11, no. 7: 584. https://doi.org/10.3390/v11070584
APA StyleBelarbi, E., Legros, V., Basset, J., Desprès, P., Roques, P., & Choumet, V. (2019). Bioluminescent Ross River Virus Allows Live Monitoring of Acute and Long-Term Alphaviral Infection by In Vivo Imaging. Viruses, 11(7), 584. https://doi.org/10.3390/v11070584