Genetic Markers Associated with Field PRRSV-Induced Abortion Rates


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Pig Populations
2.2. DNA Isolation
2.3. Primer Design and Genotyping Protocol
2.4. HDAC6 Characterization and Genotyping
2.5. Statistical Analysis
3. Results
3.1. Characterization of Sequence Variants in Pig HDAC6
3.2. Allelic and Genotypic Frequencies of the Markers
3.3. Association Study between Markers and Abortion Rate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Karniychuk, U.U.; Nauwynck, H.J. Pathogenesis and prevention of placental and transplacental porcine reproductive and respiratory syndrome virus infection. Vet. Res. 2013, 44, 95. [Google Scholar] [CrossRef] [PubMed]
- Ladinig, A.; Wilkinson, J.; Ashley, C.; Detmer, S.E.; Lunney, J.K.; Plastow, G.; Harding, J.C.S. Variation in Fetal Outcome, Viral Load and ORF5 Sequence Mutations in a Large Scale Study of Phenotypic Responses to Late Gestation Exposure to Type 2 Porcine Reproductive and Respiratory Syndrome Virus. PLoS ONE 2014, 9, e96104. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.R.G.; Ait-Ali, T.; Clapperton, M.; Archibald, A.L.; Bishop, S. Genetic perspectives on host responses to porcine reproductive and respiratory syndrome (PRRS). Viral Immunol. 2007, 20, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Vissche, A.H.; Janss, L.L.G.; Niewold, T.A.; de Greef, K.H. Disease incidence and immunological traits for the selection of healthy pigs A review. Vet. Q. 2002, 24, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.R.G.; Torremorell, M.; Galina-Pantoja, L.; Bishop, S.C. Genetic parameters for performance traits in commercial sows estimated before and after an outbreak of porcine reproductive and respiratory syndrome. J. Anim. Sci. 2009, 87, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Boddicker, N.; Waide, E.H.; Rowland, R.R.R.; Lunney, J.K.; Garrick, D.J.; Reecy, J.M.; Dekkers, J.C.M. Evidence for a major QTL associated with host response to porcine reproductive and respiratory syndrome virus challenge. J. Anim. Sci. 2012, 90, 1733–1746. [Google Scholar] [CrossRef] [PubMed]
- Boddicker, N.J.; Bjorkquist, A.; Rowland, R.R.R.; Lunney, J.K.; Reecy, J.M.; Dekkers, J.C.M. Genome-wide association and genomic prediction for host response to porcine reproductive and respiratory syndrome virus infection. Genet. Sel. Evol. GSE 2014, 46, 18. [Google Scholar] [CrossRef]
- Koltes, J.E.; Fritz-Waters, E.; Eisley, C.J.; Choi, I.; Bao, H.; Kommadath, A.; Serão, N.V.L.; Boddicker, N.J.; Abrams, S.M.; Schroyen, M.; et al. Identification of a putative quantitative trait nucleotide in guanylate binding protein 5 for host response to PRRS virus infection. BMC Genom. 2015, 16, 412. [Google Scholar] [CrossRef]
- Waide, E.H.; Tuggle, C.K.; Serão, N.V.L.; Schroyen, M.; Hess, A.; Rowland, R.R.R.; Lunney, J.K.; Plastow, G.; Dekkers, J.C.M. Genomewide association of piglet responses to infection with one of two porcine reproductive and respiratory syndrome virus isolates. J. Anim. Sci. 2017, 95, 16–38. [Google Scholar] [CrossRef]
- Lim, B.; Khatun, A.; Kim, S.W.; Nazki, S.; Jeong, C.G.; Gu, S.; Lee, J.; Lee, K.T.; Park, C.K.; Lee, S.M.; et al. Polymorphisms in the porcine CD163 associated with response to PRRSV infection. Anim. Genet. 2018, 49, 98–99. [Google Scholar] [CrossRef] [PubMed]
- Niu, P.; Shabir, N.; Khatun, A.; Seo, B.J.; Gu, S.; Lee, S.M.; Lim, S.K.; Kim, K.S.; Kim, W.I. Effect of polymorphisms in the GBP1, Mx1 and CD163 genes on host responses to PRRSV infection in pigs. Vet. Microbiol. 2016, 182, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liang, S.; Liu, H.; Sun, Y.; Kang, L.; Jiang, Y. Identification of a short interspersed repetitive element insertion polymorphism in the porcine MX1 promoter associated with resistance to porcine reproductive and respiratory syndrome virus infection. Anim. Genet. 2015, 46, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Lillico, S.G.; Barnett, M.W.; Whitelaw, C.B.A.; Archibald, A.L.; Ait-Ali, T. USP18 restricts PRRSV growth through alteration of nuclear translocation of NF-κB p65 and p50 in MARC-145 cells. Virus Res. 2012, 169, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Song, Z.; Li, Q.; Li, Z.; Wang, M.; Liu, L.; Tian, K.; Li, N. Overexpression of Histone Deacetylase 6 Enhances Resistance to Porcine Reproductive and Respiratory Syndrome Virus in Pigs. PLoS ONE 2017, 12, e0169317. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, Y.; Xing, F.; Kang, L.; Wang, P.; Wang, L.; Liu, H.; Li, Y.; Jiang, Y. Identification of a single nucleotide promoter polymorphism regulating the transcription of ubiquitin specific protease 18 gene related to the resistance to porcine reproductive and respiratory syndrome virus infection. Vet. Immunol. Immunopathol. 2014, 162, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Holtkamp, D.J.; Polson, D.D.; Torremorell, M.; Morrison, B.; Classen, D.M.; Becton, L.; Henry, S.; Rodibaugh, M.T.; Rowland, R.R.; Snelson, H.; et al. Terminology for classifying the porcine reproductive and respiratory syndrome virus (PRRSV) status of swine herds. Tierarztl. Prax. Ausg. G Grosstiere Nutztiere 2011, 39, 101–112. [Google Scholar] [PubMed]
- Abella, G.; Pena, R.N.; Nogareda, C.; Armengol, R.; Vidal, A.; Moradell, L.; Tarancon, V.; Novell, E.; Estany, J.; Fraile, L. A WUR SNP is associated with European Porcine Reproductive and Respiratory Virus Syndrome resistance and growth performance in pigs. Res. Vet. Sci. 2016, 104, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Abella, G.; Novell, E.; Tarancon, V.; Varona, L.; Pena, R.N.; Estany, J.; Fraile, L. Identification of resilient sows in porcine reproductive and respiratory syndrome virus-infected farms1. J. Anim. Sci. 2019, 97, 3228–3236. [Google Scholar] [CrossRef]
- Material not intended for publication: LJ Fraile, University of Lleida, 25198 Lleida, Spain. Sequencing of the GP5 regions of field strains from samples of sows affected by PRRSV during the cited outbreaks, 2018.
- Green, M.R.; Sambrook, J.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012. [Google Scholar]
- Hung, J.H.; Weng, Z. Designing Polymerase Chain Reaction Primers Using Primer3Plus. Cold Spring Harb. Protoc. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinform. Oxf. Engl. 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Serão, N.V.L.; Matika, O.; Kemp, R.A.; Harding, J.C.S.; Bishop, S.C.; Plastow, G.S.; Dekkers, J.C.M. Genetic analysis of reproductive traits and antibody response in a PRRS outbreak herd. J. Anim. Sci. 2014, 92, 2905–2921. [Google Scholar] [CrossRef] [PubMed]
- Reiner, G. Genetic resistance—An alternative for controlling PRRS? Porc. Health Manag. 2016, 2, 27. [Google Scholar] [CrossRef] [PubMed]
- Serão, N.V.L.; Kemp, R.A.; Mote, B.E.; Willson, P.; Harding, J.C.S.; Bishop, S.C.; Plastow, G.S.; Dekkers, J.C.M. Genetic and genomic basis of antibody response to porcine reproductive and respiratory syndrome (PRRS) in gilts and sows. Genet. Sel. Evol. 2016, 48, 51. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wilkinson, J.; Wang, Z.; Ladinig, A.; Harding, J.; Plastow, G. A genome-wide association study of fetal response to type 2 porcine reproductive and respiratory syndrome virus challenge. Sci. Rep. 2016, 6, 20305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scanlan, C.L.; Putz, A.M.; Gray, K.A.; Serão, N.V.L. Genetic analysis of reproductive performance in sows during porcine reproductive and respiratory syndrome (PRRS) and porcine epidemic diarrhea (PED) outbreaks. J. Anim. Sci. Biotechnol. 2019, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Gol, S.; Estany, J.; Fraile, L.J.; Pena, R.N. Expression profiling of the GBP1 gene as a candidate gene for porcine reproductive and respiratory syndrome resistance. Anim. Genet. 2015, 46, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Hess, A.S.; Islam, Z.; Hess, M.K.; Rowland, R.R.R.; Lunney, J.K.; Doeschl-Wilson, A.; Plastow, G.S.; Dekkers, J.C.M. Comparison of host genetic factors influencing pig response to infection with two North American isolates of porcine reproductive and respiratory syndrome virus. Genet. Sel. Evol. 2016, 48, 43. [Google Scholar] [CrossRef] [PubMed]
- Boddicker, N.J.; Garrick, D.J.; Rowland, R.R.R.; Lunney, J.K.; Reecy, J.M.; Dekkers, J.C.M. Validation and further characterization of a major quantitative trait locus associated with host response to experimental infection with porcine reproductive and respiratory syndrome virus. Anim. Genet. 2014, 45, 48–58. [Google Scholar] [CrossRef]
- Ladinig, A.; Ashley, C.; Detmer, S.E.; Wilkinson, J.M.; Lunney, J.K.; Plastow, G.; Harding, J.C.S. Maternal and fetal predictors of fetal viral load and death in third trimester, type 2 porcine reproductive and respiratory syndrome virus infected pregnant gilts. Vet. Res. 2015, 46, 107. [Google Scholar] [CrossRef]
- Wang, F.; Qiu, H.; Zhang, Q.; Peng, Z.; Liu, B. Association of two porcine reproductive and respiratory syndrome virus (PRRSV) receptor genes, CD163 and SN with immune traits. Mol. Biol. Rep. 2012, 39, 3971–3976. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, K.M.; Rowland, R.R.R.; Ewen, C.L.; Trible, B.R.; Kerrigan, M.A.; Cino-Ozuna, A.G.; Samuel, M.S.; Lightner, J.E.; McLaren, D.G.; Mileham, A.J.; et al. Gene-edited pigs are protected from porcine reproductive and respiratory syndrome virus. Nat. Biotechnol. 2016, 34, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Burkard, C.; Lillico, S.G.; Reid, E.; Jackson, B.; Mileham, A.J.; Ait-Ali, T.; Whitelaw, C.B.A.; Archibald, A.L. Precision engineering for PRRSV resistance in pigs: Macrophages from genome edited pigs lacking CD163 SRCR5 domain are fully resistant to both PRRSV genotypes while maintaining biological function. PLoS Pathog. 2017, 13, e1006206. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Staeheli, P.; Hufbauer, M.; Koerner, I.; Martínez-Sobrido, L.; Solórzano, A.; García-Sastre, A.; Haller, O.; Kochs, G. Replication fitness determines high virulence of influenza A virus in mice carrying functional Mx1 resistance gene. Proc. Natl. Acad. Sci. USA 2007, 104, 6806–6811. [Google Scholar] [CrossRef] [PubMed]
- Ciancanelli, M.J.; Abel, L.; Zhang, S.Y.; Casanova, J.L. Host genetics of severe influenza: From mouse Mx1 to human IRF7. Curr. Opin. Immunol. 2016, 38, 109–120. [Google Scholar] [CrossRef] [PubMed]
- De Groot, A.E.; Zarif, J.C.; Pienta, K.J. Abstract 4005: HDAC inhibitors regulate M2 tumor-associated macrophage function through histone acetylation. Tumor Biol. 2017, 77, 4005. [Google Scholar]
- Yan, B.; Xie, S.; Liu, Z.; Ran, J.; Li, Y.; Wang, J.; Yang, Y.; Zhou, J.; Li, D.; Liu, M. HDAC6 deacetylase activity is critical for lipopolysaccharide-induced activation of macrophages. PLoS ONE 2014, 9, e110718. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Jiang, Y.; He, Z.; Kitazato, K.; Wang, Y. Cellular defence or viral assist: The dilemma of HDAC6. J. Gen. Virol. 2017, 98, 322–337. [Google Scholar] [CrossRef] [PubMed]
- Tayade, C.; Black, G.P.; Fang, Y.; Croy, B.A. Differential Gene Expression in Endometrium, Endometrial Lymphocytes, and Trophoblasts during Successful and Abortive Embryo Implantation. J. Immunol. 2006, 176, 148–156. [Google Scholar] [CrossRef]
- Höglund, J.K.; Buitenhuis, A.J.; Guldbrandtsen, B.; Su, G.; Thomsen, B.; Lund, M.S. Overlapping chromosomal regions for fertility traits and production traits in the Danish Holstein population. J. Dairy Sci. 2009, 92, 5712–5719. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Marker | Gene Acronym | Gene Function | Polymorphism | Gene Location | Chromosomal Location 1 | Reference |
|---|---|---|---|---|---|---|
| rs80800372 | GBP1 | Interferon-induced guanylate binding protein with known antiviral functions | A>G | 3’UTR | SSC4 | [7] |
| rs340943904 | GBP5 | Inflammasome assembly, innate immunity | G>T | Intron 5 | SSC4 | [9] |
| c.3534C>T | CD163 | Macrophage-specific scavenger receptor, mediates PRRSV entry into macrophages | C>T | 3’UTR | SSC5 | [11] |
| rs1107556229 | G>A | Exon 10 | SSC5 | [11] | ||
| -547ins+275 | MX1 | Interferon-induced GTP metabolizing enzyme, antiviral properties | Indel 275 bp | Promoter | SSC13 | [12] |
| -1533G>A | USP18 | Ubiquitin-specific proteases, Downregulation of interferon responses | G>A | Promoter | SSC5 | [16] |
| rs325981825 | HDAC6 | Epigenetic labeling of histones by acetylation/deacetylation | G>A | Exon 3 | SSCX | This study |
| g.2360C>T | C>T | Exon 15 | SSCX | This study |
| HDAC6 Fragment | Polymorphism | Position from ATG * | Location | Change Type |
|---|---|---|---|---|
| Fragment 1 | C/T | −1538 | Exon 1 | 5′UTR |
| Fragment 2 | G/A | +35 | Exon 3 | Missense (Arg12Lys) |
| C/G | +63 | Exon 3 | Synonym (His21) | |
| Fragment 3 | G/A | +2180 | Intron 13 | - |
| G/A | +2222 | Exon 14 | Synonym (Gln337) | |
| G/A | +2340 | Intron 14 | - | |
| C/T | +2360 | Exon 15 | Missense (Pro360Leu) | |
| Fragment 4 | C/A | +3785 | Exon 19 | Missense (Pro503His) |
| Fragment 5 | G/T | +9813 | Exon 25 | Synonym (Gln799) |
| C/A | +10450 | Intron 26 | - |
| Polymorphism | Protein Domain (EMBL-EBI) | SIFT Prediction (Score) | Polyphen-2 Prediction (Score) |
|---|---|---|---|
| Arg12Lys | Not tolerant (0.00) * | Unknown, not enough reference sequences | |
| Pro360Leu | Hist_deacetyl (PF00850) | Not tolerant (0.02) | Probably damaging (1.000) |
| Pro503His | Hist_deacetyl (PF00850) | Not tolerant (0.00) | Probably damaging (1.000) |
| Marker | Gene | MAF (Allele) | AA | AB | BB * |
|---|---|---|---|---|---|
| rs80800372 | GBP1 | 0.19 (G) | 15 | 64 | 171 |
| rs340943904 | GBP5 | 0.25 (T) | 22 | 82 | 144 |
| c.3534C>T | CD163 | 0.33 (C) | 41 | 70 | 119 |
| rs1107556229 | CD163 | 0.29 (A) | 32 | 70 | 127 |
| −547ins+275 | MX1 | 0.25 (insertion) | 18 | 50 | 170 |
| rs325981825 | HDAC6 | 0.37 (A) | 35 | 103 | 90 |
| g.2360C>T | HDAC6 | 0.38 (T) | 32 | 104 | 32 |
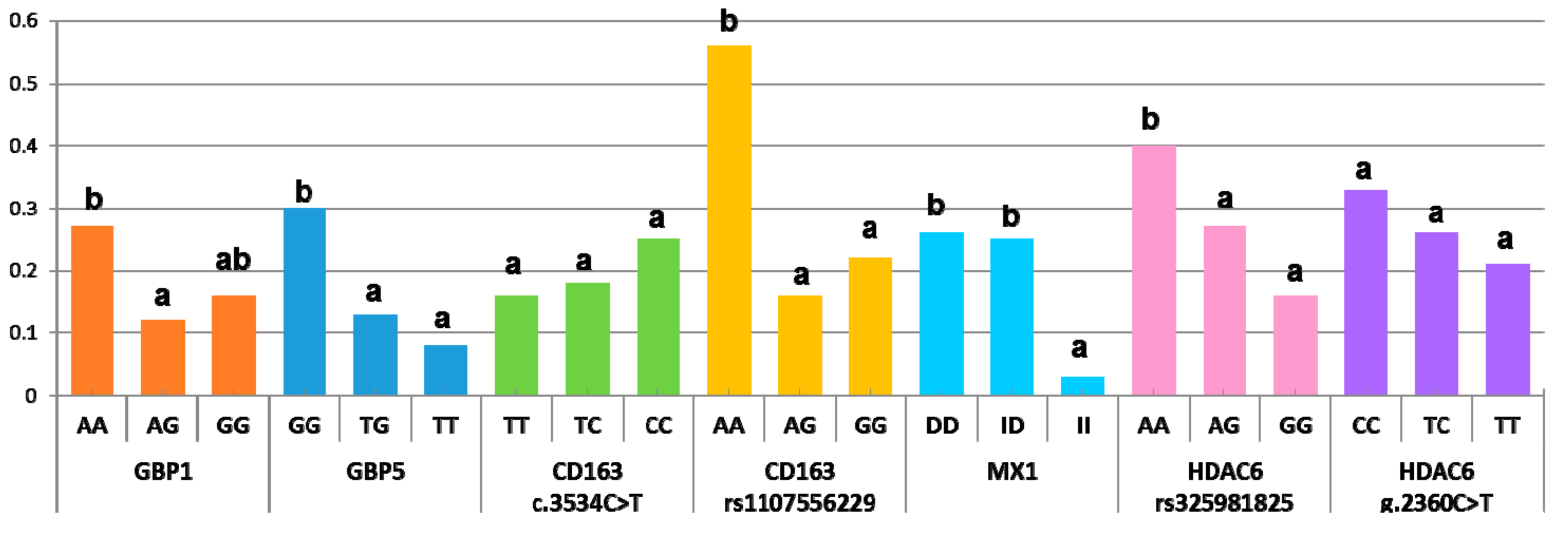
| Marker | Gene | Contrast | Odds Ratio | p |
|---|---|---|---|---|
| rs80800372 | GBP1 | AA/AG | 2.69 | 0.008 |
| rs340943904 | GBP5 | GG/TG | 2.76 | 0.003 |
| GG/TT | 4.49 | 0.02 | ||
| rs1107556229 | CD163 | AA/AG | 2.58 | <0.0001 |
| AA/GG | 1.96 | 0.0004 | ||
| −547ins+275 | MX1 | DD/II | 9.35 | 0.03 |
| ID/II | 8.63 | 0.04 | ||
| rs325981825 | HDAC6 | AA/GG | 4.08 | 0.002 |
| AG/GG | 2.34 | 0.02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pena, R.N.; Fernández, C.; Blasco-Felip, M.; Fraile, L.J.; Estany, J. Genetic Markers Associated with Field PRRSV-Induced Abortion Rates. Viruses 2019, 11, 706. https://doi.org/10.3390/v11080706
Pena RN, Fernández C, Blasco-Felip M, Fraile LJ, Estany J. Genetic Markers Associated with Field PRRSV-Induced Abortion Rates. Viruses. 2019; 11(8):706. https://doi.org/10.3390/v11080706
Chicago/Turabian StylePena, Ramona N., Carlos Fernández, María Blasco-Felip, Lorenzo J. Fraile, and Joan Estany. 2019. "Genetic Markers Associated with Field PRRSV-Induced Abortion Rates" Viruses 11, no. 8: 706. https://doi.org/10.3390/v11080706
APA StylePena, R. N., Fernández, C., Blasco-Felip, M., Fraile, L. J., & Estany, J. (2019). Genetic Markers Associated with Field PRRSV-Induced Abortion Rates. Viruses, 11(8), 706. https://doi.org/10.3390/v11080706