1. Introduction
DENV infection is a common arbovirus illness in humans [
1]. DENV has four serotypes (one to four) and belongs to the genus
Flavivirus within the family Flaviviridae. In addition to DENV, other viruses in the genus
Flavivirus, such as the West Nile virus (WNV), Japanese encephalitis virus (JEV), yellow fever virus, and tick-borne encephalitis virus, are pathogens of humans and animals. After acute dengue viral infection, patients often develop fever, rash, headache, muscle and joint pain, and nausea [
2]. The most severe form of the disease is dengue hemorrhagic fever/dengue shock syndrome, which is associated with vascular leakage, hemorrhage, and shock [
3]. More than 3.5 billion people are at risk for DENV infection. It has been estimated that each year, up to 96 million of the 390 million DENV infections in the world are symptomatic [
1].
The
Flavivirus genome is a single-stranded, positive-sense RNA of approximately 11 kb in length. Its genomic RNA consists of a 5′ untranslated region (UTR), an open reading frame (ORF), and a 3′ UTR [
4]. The ORF encodes a polyprotein that is processed into three structural proteins (capsid (C), premembrane (prM), and envelope (E)) and seven nonstructural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) [
5,
6,
7]. The structural proteins are the main components of the virion. The nonstructural proteins are important for viral RNA replication [
8].
Flavivirus NS4B is essential for viral RNA replication [
9,
10,
11] and the evasion of the host immune response [
12,
13]. NS4B, 27 kDa, is an integral membrane protein that has five predicted transmembrane domains (TMD) [
14]. The pTMD4 of the NS4A region (2K) serves as a signal sequence for the translocation of the adjacent NS4B into the endoplasmic reticulum (ER) lumen [
15]. DENV NS4B is one of the membrane-bound viral replication complexes in the ER [
14]. The DENV NS4B expression was shown to modulate innate immunity signaling by blocking the α/β interferon pathway through the inhibition of STAT1 phosphorylation, dimerization, and translocation to the nucleus [
12,
13]. Furthermore, NS4B is able to induce mitochondrial elongation and inhibit the activation of the immune response elicited by the mitochondrial antiviral-signaling protein-dependent interaction of the ER with mitochondria to promote infection [
16]. The DENV-2 NS4B protein expression also modulates the unfolded protein response (UPR) transcriptional activation. The expression of the DENV NS4B protein suppressed the induction of heavy-chain binding protein (BiP/GRP78) and EDEM in the unfolded protein response (UPR) [
17]. Little is known regarding the interaction between NS4B and the host factors to support the virus life-cycle. In this study, we identified the interaction of stress-associated endoplasmic reticulum protein 1 (SERP1) with dengue virus type 2 (DENV-2) NS4B by a membrane-based split-ubiquitin yeast two-hybrid system.
The detailed function of SERP1 in the cells is not clear. SERP1, also known as ribosome-associated membrane protein 4 (RAMP4) [
18], is a tail-anchored protein with an N-terminus exposed on the cytoplasmic (residues 1–115), and a C-terminus inserted on the luminal side of the ER membrane (residues 116–198) [
19]. SERP1 was reported to interact with subunits (Sec 61α and Sec 61β) of the translocon [
18], which acts as a channel and mediates the translocation of polypeptides across membranes to aid protein synthesis [
20]. Thus, SERP1 is implicated in the regulation of membrane protein biogenesis. SERP1 overexpression caused the newly synthesized integral membrane proteins to degrade, and then facilitated protein glycosylation in order to protect cells from ER stress [
21]. The genetic ablation of SERP1 showed that the SERP1
−/− mice had growth retardation, increased mortality, impaired glucose tolerance, and ER stress. In the pituitary, the increased activation of molecules associated with ER stress (P-eIF2α) and apoptosis (C/EBP homologous protein and caspase 3) resulted in a higher mortality in the SERP1
−/− mice than in wild-type (WT) mice [
22].
An increased production of DENV proteins during dengue infection leads to the accumulation of misfolded and unfold proteins in the ER [
23]. This accumulation results in ER stress and activation of the UPR as a host response to alleviate ER stress. Tunicamycin, thapsigargin, and dithiothreitol induce ER stress and trigger three initiation branches of the UPR, inositol-requiring protein 1 (IRE1), protein kinase RNA-like ER kinase (PERK), and activating transcription factor 6 (ATF6) [
24,
25]. Similarly, DENV infection induces these three branches of the UPR [
26,
27] at different infectious stages during ER stress [
28]. The PERK arm is activated early in DENV infection, followed by IRE1-XBP1 mid-infection, and ATF6 later during the infection. DENV has developed a strategy to manipulate the host UPR pathways in order to enhance its survival and viral replication for the successful completion of the virus life-cycle.
In this study, we identified a cellular protein, SERP1, that interacted with DENV-2 NS4B and was involved in the DENV life-cycle. SERP1 is induced in Huh7.5 cells by either DENV-2 infection or the transfection of DENV replicon. Cells stably overexpressing SERP1 showed a significant decrease in DENV-2 yields and RNA replication capacity. In contrast, the overexpression of NS4B alleviated the inhibitory effect of SERP1 overexpression on virus replication. The results of the present study suggest that SERP1 is an antiviral protein, and that the interaction of DENV-2 NS4B with SERP1 may counteract the antagonistic effect of SERP1 on the DENV-2 life-cycle.
2. Materials and Methods
2.1. Cell Lines and Dengue Virus Strain
The HEK-293 cells were maintained in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) at 37 °C in a 5% CO2 incubator. Baby hamster kidney (BHK21) clone 15 cells were kindly provided by Dr. Robert Beatty (Department of Molecular and Cell Biology, University of California at Berkeley, Berkeley, CA, USA), and were cultured in alpha-MEM supplemented with 5% FBS at 37 °C in a 5% CO2 incubator. The human hepatocyte cell line Huh7.5 cells were maintained in DMEM supplemented with 10% FBS at 37 °C in a 5% CO2 incubator. The dengue virus type 2 (DENV-2; strain 16681) was used for the infection of the Huh7.5 and HEK-293 cells.
2.2. Identification of Host Factors Interacting with DENV-2 Proteins by a Membrane-Based Split-Ubiquitin Yeast Two-Hybrid System
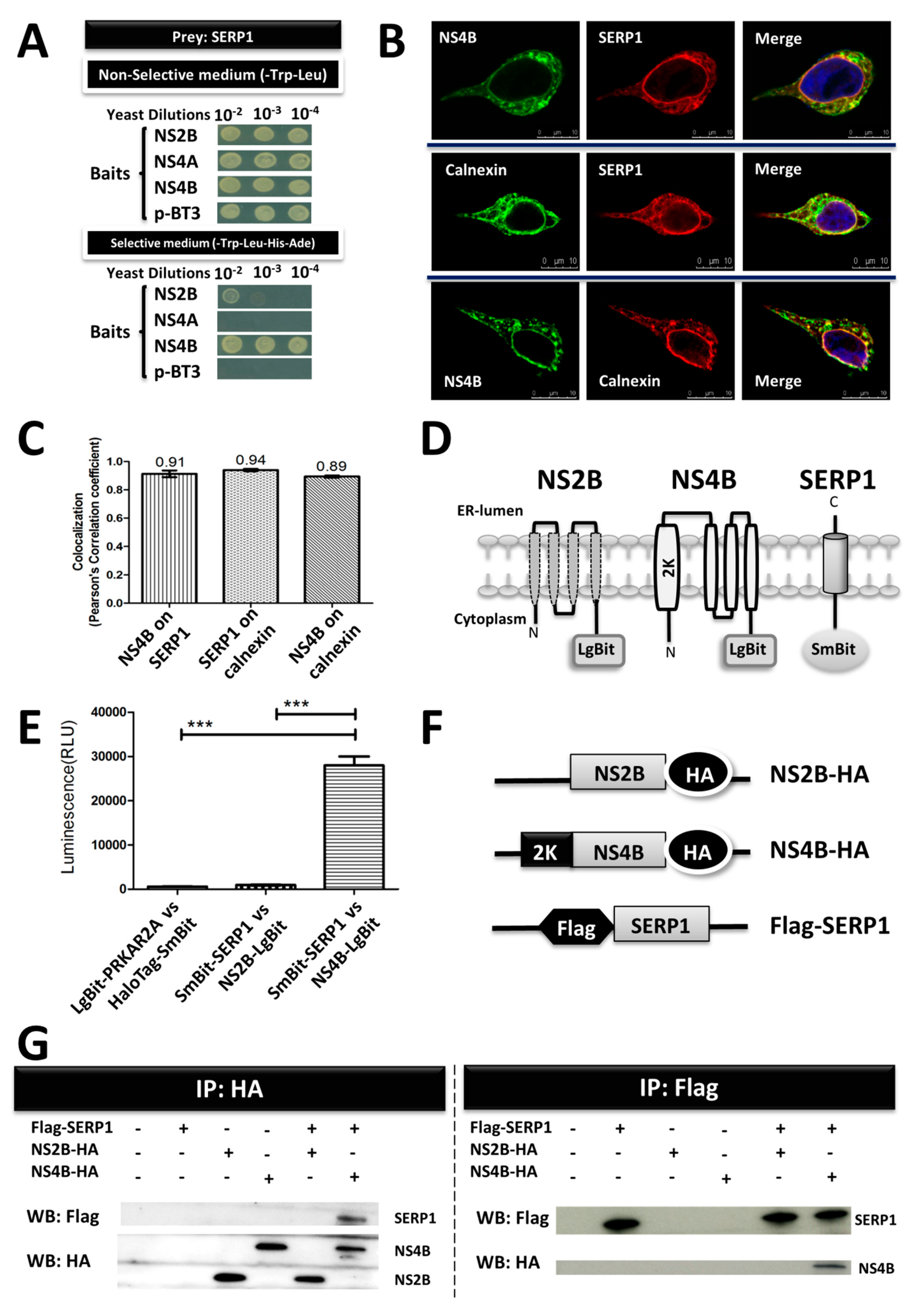
The membrane-based split-ubiquitin yeast two-hybrid system (MYTH), DUAL membrane technology, Dualsystems Biotech, Schlieren, Switzerland, was applied to screen NS2B, NS4A, and NS4B (DENV-2, strain 16681) against a human liver cDNA library. MYTH was performed as previously described [
29]. The cDNAs encoding the NS2B, NS4A, and NS4B proteins were inserted into a yeast expression vector, p-BT3, fused with C-terminal ubiquitin and artificial transcription factor (TF) Lex-VP16 as baits. In addition, the pPR3N prey vector, harboring a human liver cDNA library, was fused with N-terminal ubiquitin as an interactor protein. The
S. cerevisiae strain NMY32 was co-transformed with baits and prey plasmids so as to detect the protein–protein interactions (PPIs) of integral membrane proteins. On the interaction of the bait and prey, the N-terminal ubiquitin and C-terminal ubiquitin reconstituted a form of ubiquitin, which caused the ubiquitin-specific-proteases to be released downstream of the TF. The TF translocated into the nucleus and activated the auxotrophic reporter genes, allowing yeast growth on a selective medium. Ten-fold serial yeast dilutions were spotted onto nonselective plates (lacking tryptophan and leucine (-Trp–Leu)) and selective plates (lacking tryptophan, leucine, histidine, and adenine (-Trp–Leu–His–Ade)) for the detection of protein–protein interactions.
2.3. Measurement of SERP1 Protein Levels in Flag-SERP1-Overexpressing Huh7.5 Cells by Immunoprecipitation Analysis
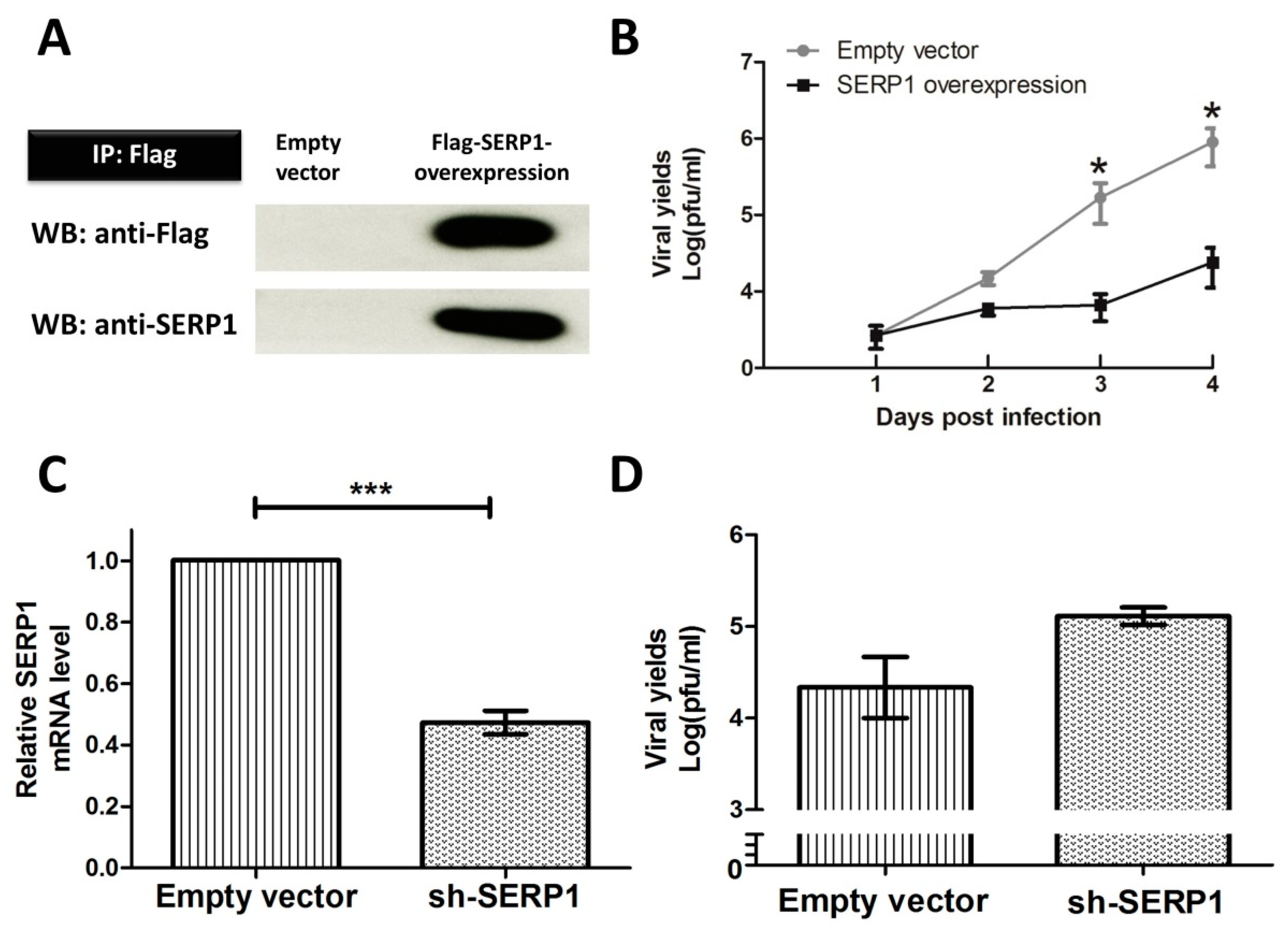
The lentiviral vector pLKO-AS2 was purchased from the National RNAi Core Facility, Academia Sinica, Taipei, Taiwan. The full-length SERP1 protein with an N-terminal Flag tag was cloned into the lentiviral vector (pLKO-AS2-Flag-SERP1) in order to overexpress the protein. The HEK 293T cells were co-transfected with the lentiviral plasmid (pLKO-AS2-Flag-SERP1) and two helper plasmids, pCMV-ΔR 8.91 and pMD.G, by Lipofectamine 2000 (Invitrogen, California, USA). The cell supernatants containing lentiviruses were harvested 24 and 48 h after transfection. The viral titers were assayed with a Lenti-X p24 Rapid Titer Kit (Clontech, Mountain View, CA, USA). The Huh7.5 cells were infected with lentiviruses (multiplicity of infection (MOI) = 10)) in a medium containing polybrene (8 µg/mL). Three days after infection, the cells were treated with puromycin (Invitrogen) so as to derive a pool of resistant clones.
The stable Huh7.5 cells overexpressing Flag-tagged SERP1 and the cells receiving an empty vector were lysed in a radioimmunoprecipitation assay (RIPA) buffer, and the protein concentration was determined using a Pierce BCA Protein Assay Kit (Thermo Scientific, Rockford, IL, USA). Equal amounts of cell lysates were analyzed by immunoprecipitation using anti-Flag M2 affinity gel (A2220, Sigma, Missouri, USA) for 2 h at 4 °C. The beads were washed with a lysis buffer and were eluted. The proteins were fractionated on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and were transferred to the nitrocellulose membranes. An immunoblot analysis was performed using anti-Flag antibody (GTX115043, Genetex, CA, USA) and anti-SERP1 antibody (ab130974, Abcam, Cambridge, MA, USA).
2.4. Measurement of Viral Protein Levels by Immunoprecipitation Analysis
To express the NS2B and NS4B proteins, the full-length NS2B and NS4B proteins with or without an N-terminal 2K signal peptide were cloned into the lentiviral vector pLKO-AS2. All of the constructs (pLKO-AS2-NS2B-HA, pLKO-AS2-△2K-NS4B-HA, and pLKO-AS2-NS4B-HA) were expressed with the HA tag epitope fused at the carboxyl terminals. The pLKO-AS2-△2K-NS4B-HA plasmid contained the full-length NS4B lacking an N-terminal 2K signal peptide of NS4A. The parental Huh7.5 cells and Flag-SERP1-overexpressing Huh7.5 cells were transfected with pLKO-AS2-NS2B-HA, pLKO-AS2-△2K-NS4B-HA, or pLKO-AS2-NS4B-HA plasmids. Equal amounts of cell lysates were analyzed by immunoprecipitation using anti-HA magnetic beads (88836, Thermo Fisher Scientific, Waltham, MA, USA), and the HA-tagged proteins were detected by Western blot (WB) using the rabbit polyclonal anti-HA antibody (SG77, Thermo Fisher Scientific, Invitrogen).
2.5. The Localization of NS4B and SERP1 in the Endoplasmic Reticulum by Confocal Microscopy
The Flag-SERP1-overexpressing Huh7.5 cells were infected with DENV-2 (multiplicity of infection (MOI) of 5). At 3 d.p.i, the Huh7.5 cells were washed once with phosphate-buffered saline (PBS), fixed with 1% paraformaldehyde for 1 h at room temperature, permeabilized with methanol for 15 min at 4 °C, and blocked with 2% horse serum in PBS. Rabbit polyclonal anti-NS4B (GTX103349, GeneTex), Flag-tag mouse antibody (F1804, Sigma), and mouse monoclonal anti-calnexin (M178-3, MBL Life Science, Nagoya, Japan) were used as the primary antibodies. Alexa Fluor 488 anti-mouse IgG antibody and Alexa Fluor® 568 anti-rabbit IgG H&L antibody (Molecular Probes, Invitrogen) were used as the secondary antibodies. Finally, the coverslips were washed extensively and fixed onto slides. The images were captured by a Leica TCS SP5II confocal microscope (CLSM, Heidelberg, Germany). A colocalization analysis was performed using LAS AF software (Leica Microsystems, Germany) in order to analyze the colocalization of the fluorescent signals.
2.6. NanoBiT Complementation Assay of SERP1 and NS4B for Protein–Protein Interactions
NanoLuc® Luciferase is divided into two subunits, large BiT (LgBiT; 18 kDa) and small BiT (SmBiT; 1 kDa peptide), which are expressed as fusions in order to target proteins of interest (Promega). The entire regions of NS2B, NS4B, or SERP1 were coupled to Nano-Luc Binary Technology (NanoBiT) vectors (pBiT1.1-C (TK/LgBiT) vector, pBiT2.1-C (TK/SmBiT) vector, pBiT1.1-N (TK/LgBiT) vector, and pBiT2.1-N (TK/SmBiT) vector; Promega, Madison, WI, USA), according to the manufacturer’s instructions. The HEK-293 cells were cultured in DMEM (Sigma-Aldrich) containing 10% FBS at a density of 1 × 104 cells/well in 96-well plates for 24 h. The HEK-293 cells were transfected with a 40 ng LgBiT fusion construct and a 40 ng SmBiT fusion construct using Lipofectamine 2000 (Invitrogen). Twenty-four hours after transfection, the cells were serum-starved for 30 min in 0% FBS in an Opti-MEM I medium, and the relative luminescence unit (RLU) was measured using a Nano-Glo Live Cell Assay System (Promega) and GloMax® Discover System (Promega).
2.7. Interaction of SERP1 with NS4B by Co-Immunoprecipitation (Co-IP)
The Huh7.5 cells were transfected with expression constructs for pLKO-AS2-Flag-SERP1, pLKO-AS2-NS2B-HA, or pLKO-AS2-NS4B-HA. At three days post-transfection, the membrane proteins in the Huh7.5 cells were isolated with Mem-PER™ Plus Membrane Protein Extraction Kit, according to the manufacturer’s protocol (89842, Thermo Fisher Scientific, USA). The protein concentration was determined using a Pierce BCA Protein Assay Kit (23225, Thermo Fisher Scientific, USA). Equal amounts of protein complexes were analyzed by immunoprecipitation using anti-Flag M2 affinity gel (A2220, Sigma) or anti-HA magnetic beads (88836, Thermo Fisher Scientific, USA). The proteins were fractionated on 10% SDS-PAGE and were transferred to nitrocellulose membranes. Immunoblot analyses were performed using anti-Flag antibody (GTX115043, Genetex, CA, USA) and rabbit polyclonal anti-HA antibody (SG77, Thermo Fisher Scientific, USA).
2.8. Determination of Viral Yields through Plaque-Forming Assay
A plaque-forming assay was used to determine the viral titer. The BHK21 clone 15 cells were seeded in 12-well plates (4 × 10
4 cells per well) containing 1 mL of medium, and were incubated overnight. Then, 0.1 mL of aliquots of a serially diluted virus solution was added to ~70%–80% confluent BHK21 cells. After a 6-h adsorption period, the virus solutions were replaced with 0.75% methylcellulose (Sigma) and 2% FBS. The methylcellulose solution was removed from the wells, and the cells were fixed and stained with a naphthol blue-black solution (0.1% naphthol blue-black, 1.36% sodium acetate, and 6% glacial acetic acid) at six days post-infection. The number of plaque forming units (PFU) per milliliter of DENV-2 was then determined [
30].
2.9. Establishment of Stable Cells Expressing shSERP1
A lentiviral vector expressing short hairpin RNA, pLKO-1 (National RNAi Core Facility, Academia Sinica, Taipei, Taiwan), was applied to knock down the SERP1 expression. The short hairpin RNA (shRNA) targeting SERP1 (5′-CCCGAAGAGAAGGCGTCTGTA-3′) was used to transfect the packaging HEK 293T cells with the helper vectors pCMV-ΔR 8.91 and pMD.G with Lipofectamine 2000 (Invitrogen). The supernatants containing lentiviral particles were harvested, and the viral titers were assayed with a Lenti-X p24 Rapid Titer Kit (Clontech). The HEK-293 cells were infected with lentiviruses (MOI = 10) in medium containing polybrene (8 µg/mL). Three days after infection, the HEK-293 cells were treated with puromycin to derive a pool of puromycin-resistant clones. Knockdown efficiency was assayed using quantitative reverse transcription-polymerase chain reaction (qRT-PCR).
2.10. SERP1 Knockout Cells Generated using the Type II Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) System
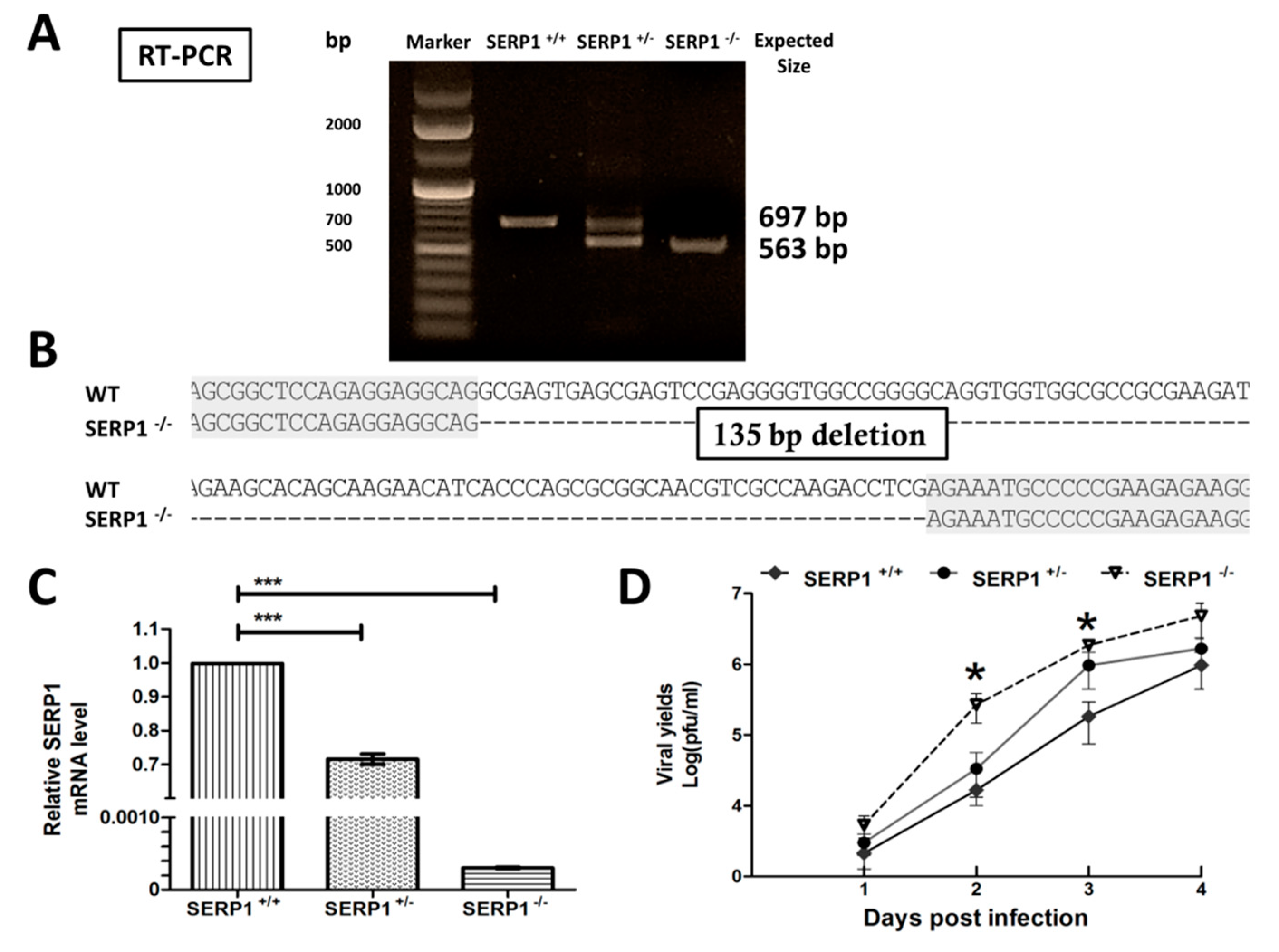
The SERP1
−/− Huh7.5 cells were generated by the CRISPR system. This system consists of a bacterial CRISPR-associated endonuclease (Cas9) and small guide RNAs (sgRNAs). Endonuclease Cas9 paired with the sgRNA target DNA causes double-strand breaks (DSBs) [
31]. Two target sites (5′-GGCGAGTGAGCGAGTCCGAGGG-3′ and 5′-GCCCCTCTGCGTTCCGCAGCCGG-3′) were chosen from SERP1 exon 1, and were then inserted into the sgRNA expression vectors. The co-expression of the Cas9 and sgRNA plasmids, which were introduced into the Huh7.5 cells by electroporation using microporation (Microporator MP100, Digital Bio, Labtech France), induced site-specific DNA cleavage. Consequently, the SERP1 protein was disrupted at the endogenous level. Individual clones were isolated from the Huh7.5 cell populations. Genomic DNA was extracted using a GeneJET Genomic DNA Purification Kit (Thermo Fisher Scientific). The SERP1 region was amplified at exon 1 by PCR, using the forward primer 5′-GCCAGTTCTCTTCCTCCTGC-3′ and reverse primer 5′-TCACAGGTCTCCCCTTCCGT-3′. The PCR products were cloned into the pGEM-T Easy (Promega) vector and were sequenced. The total RNA was isolated from the Huh7.5 knockout sublines (SERP1
+/− and SERP1
−/−) using an RNeasy Mini Kit (Qiagen, Hilden, Germany), and the RNA was treated with an RNase-Free DNase set (QIAGEN; cat. no. 79254) at 30 °C for 30 min to remove potential DNA contamination (Promega, Madison, WI, USA). Reverse transcription was performed using a RevertAid H minus First Strand cDNA Synthesis Kit (Thermo Scientific). The SERP1 region was amplified from exon 1 to exon 3 by PCR, by using the forward primer 5′-AACGCGCACGCGCA-3′ and reverse primer 5′-ATCCTGATACTTTGAATAATC T-3′. The RT-PCR products were sequenced.
2.11. Measurement of SERP1 RNA Levels in the Knockdown Cells and Knockout Sublines by qRT-PCR
The total RNA was isolated from the parental Huh7.5 cells, parental HEK-293 cells, knockdown cells, and knockout sublines using an RNeasy Mini Kit (Qiagen, Hilden, Germany), and the RNA was treated with an RNase-Free DNase set (QIAGEN, cat. no. 79254) at 30 °C for 30 min to remove the contaminating DNA from the RNA preparations. Reverse transcription was performed using a RevertAid H minus First Strand cDNA Synthesis Kit (Thermo Scientific). The specific primers for SERP1 were designed (forward primer: 5′-CGAGCCGAGCCTCGCAGCGGCTC-3′ and reverse primer: 5′-ACCAGGGTCCTACAGACGCCTTC-3′). The expression level of SERP1 mRNA was determined by quantitative real-time PCR (ViiA 7 Real-Time PCR System, Life Technologies) using KAPA™ SYBR
® FAST qPCR Kit Master Mix (2×) Universal (Kapa Biosystems). The levels of β-actin mRNA were used as an internal control (forward primer: 5′-TGGATCAGCAAGCAGGAGTATG-3′ and reverse primer: 5′-GCATTTGCGGTGGACGAT-3′) [
32]. The amplification program consisted of 40 cycles (each cycle is 95 °C for 3 s, annealing at 60 °C for 30 s) after 95 °C for 3 min. Fluorescent products were detected at the last step of each cycle. The relative quantitative values of SERP1 were normalized to the expression of β-actin. The 2
−△△CT method was used to calculate the relative changes in the SERP1 expression.
2.12. Transient Replication Activity Assay of the DNA-Launched DENV-2 Replicon
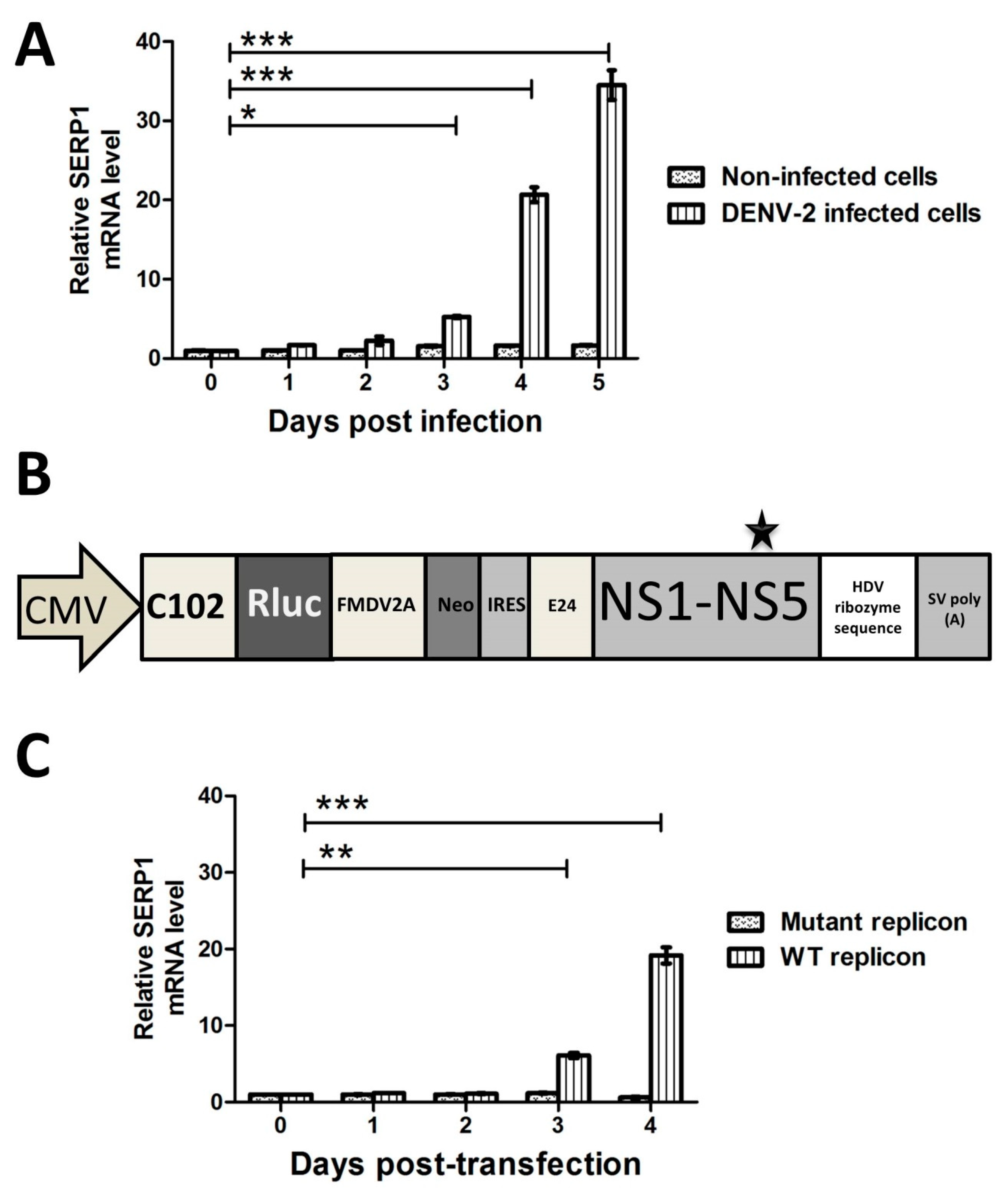
The plasmid pTight-DENV-2 is a DNA-launched infectious clone of DENV2 strain 16681. A DNA launched replicon, pCMV-DV2Rep, derived from the pTight-DENV-2-16681 plasmid, expressed Capsid protein (C), humanized Renilla luciferase (hRluc), the foot-and-mouth disease virus NS2A (FMDV2A) autoprotease, a neomycin resistance gene (Neo), an internal ribosomal entry site (IRES), C-terminal 24 amino acids of Envelope protein (E24), and NS1–NS5. As a negative control, a replicon with an inactivating mutation (GDD to GAA) in the catalytic site of the NS5 RNA-dependent RNA polymerase (RdRp) was used. A transient replication activity assay was performed to monitor the replication efficiencies of the DENV-2 replicons [
33,
34]. The Huh7.5 cells (parental cells, SERP1-overexpressing cells, and SERP1 knockout cells) were plated in 24-well plates (2 × 104 cells per well) and were co-transfected with 0.1 μg pCMV-DV2Rep and 0.1 μg pTET-OFF using Lipofectamine 2000 (Invitrogen), according to the manufacturer’s protocol. At 24 h post-transfection, doxycycline was added to the culture medium at a final concentration of 2 µg/mL to turn off the DENV-2 gene expression driven by the CMVmin promoter. The luciferase activity was measured using a GloMAX 20/20 luminometer (Promega).
4. Discussion
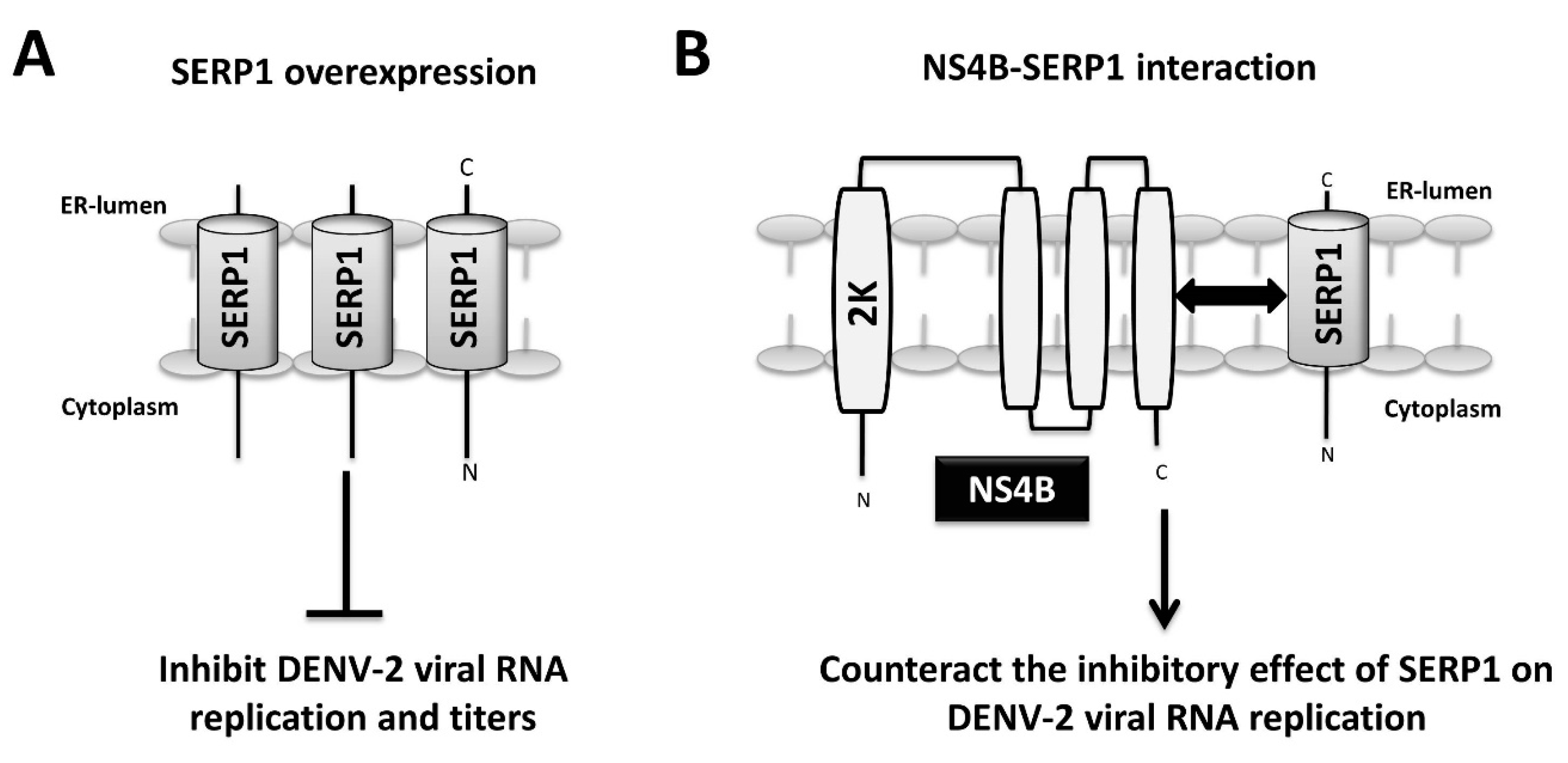
In the present study, the host factor SERP1 was identified as a DENV-2 NS4B-interacting protein. The SERP1 expression was induced upon DENV-2 infection and viral RNA replication in the Huh7.5 cells. The knockdown and knockout of SERP1 cells increased the viral yields in the DENV-2-infected cells. The SERP1 overexpression inhibited the viral yields at the viral replication step of the viral life-cycle in the Huh7.5 cells. The overexpression of NS4B in the SERP1-expressing Huh7.5 cells rescued the viral RNA replication inhibited by SERP1 overexpression. The results indicated that the inhibitory effect of SERP1 on the DENV-2 RNA replication is alleviated by the NS4B protein, possibly through the NS4B–SERP1 interaction. We proposed a hypothetical model in which DENV-2 infection induces ER stress and results in the induction of SERP1 expression. The DENV-2 NS4B synthesis occurs during viral RNA replication, and may counteract the inhibitory effect of SERP1 on DENV-2 viral replication via the interaction of NS4B with SERP1 during DENV-2 infection (
Figure 7A,B).
The SERP1 expression was greatly induced in the Huh7.5 cells either infected with DENV-2 or transfected with DENV-2 replicon at 4–5 d.p.i, whereas the SERP1 expression was very low in the cells without DENV-2 infection (
Figure 2A). The SERP1 induction phenomenon was previously shown to occur in response to various stresses (e.g., hypoxia, ischemic, high glucose, and ER stress response). For example, astrocytes exposed to hypoxia show an increase in SERP1 transcripts in a time-dependent manner, and an approximately 5.7-fold induction of SERP1 antigen in a rat ischemic brain [
21]. The SERP1 expression is also induced in cells by treatment with tunicamycin, an endoplasmic reticulum stress inducer [
35]. Both the SERP1 transcript and antigen levels are enhanced in the cells under high-glucose conditions [
22]. The results suggest that SERP1 may serve as a general stress-induced protein upregulated by various stresses.
The molecular mechanism by which the overexpression of SERP1 inhibits DENV-2 viral RNA replication in Huh7.5 cells is not clear. Little is known regarding the upstream ER stress signaling responsible for SERP1 induction. Our observation supports an important role for SERP1 expression during DENV-2 infection. For example, a drastic reduction in viral yields was found in the DENV-2-infected cells overexpressing SERP1. In contrast, viral yields significantly increased in the SERP1
−/− cells infected with DENV-2 (
Figure 4D). ER stress induction in the cells infected by DENV has been suggested to modulate virus replication during the virus life-cycle [
36]. Previous studies have indicated that the XBP1 knockout in MEFs results in a reduction in several UPR genes, SERP1, IRE1, ERdj4, p58IPK, EDEM, PDI-P5, and HEDJ, confirming their XBP1 dependency [
35]. These results suggest that XBP1 is able to regulate the SERP1 expression, implying that SERP1 may belong to the IRE1-XBP1 UPR signaling pathway. Interestingly, the IRE1
−/− and congenic MEF cell lines displayed 10-fold lower DENV viral titers, whereas the XBP1
−/− congenic MEFs result in no apparent differences in the DENV titers. These results suggest a redundancy in this part of the UPR pathway, and imply that SERP1 is not solely regulated by XBP1. Further studies are needed to illustrate the role and regulation of SERP1 in the UPR signaling pathway.
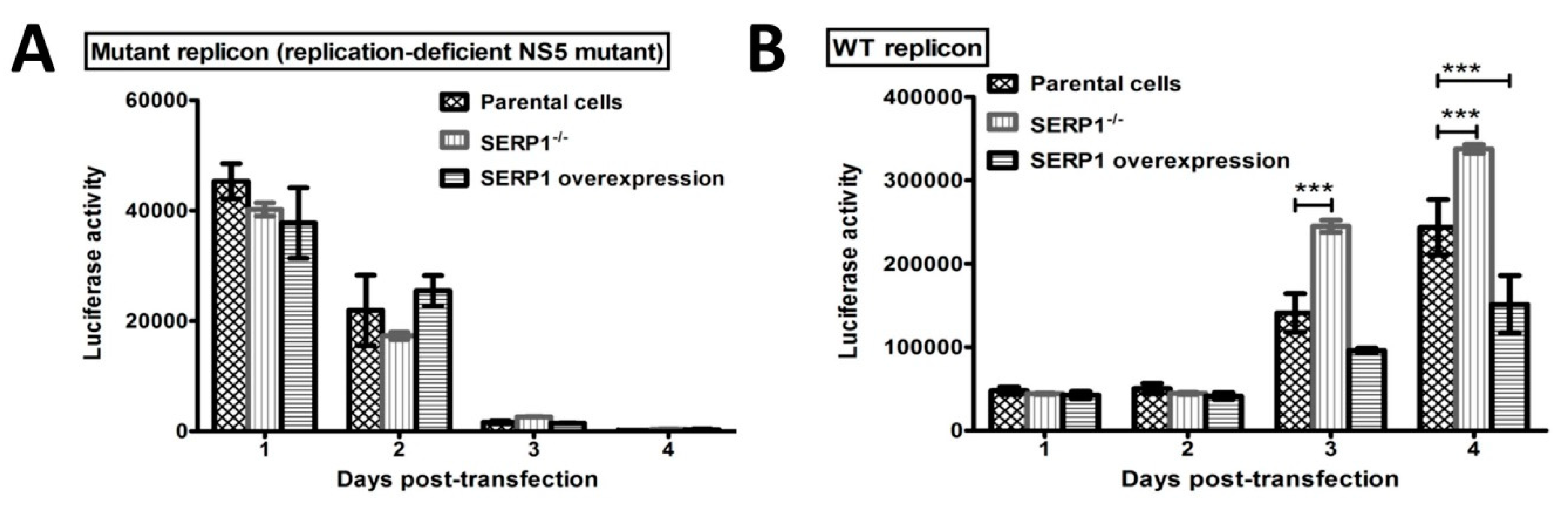
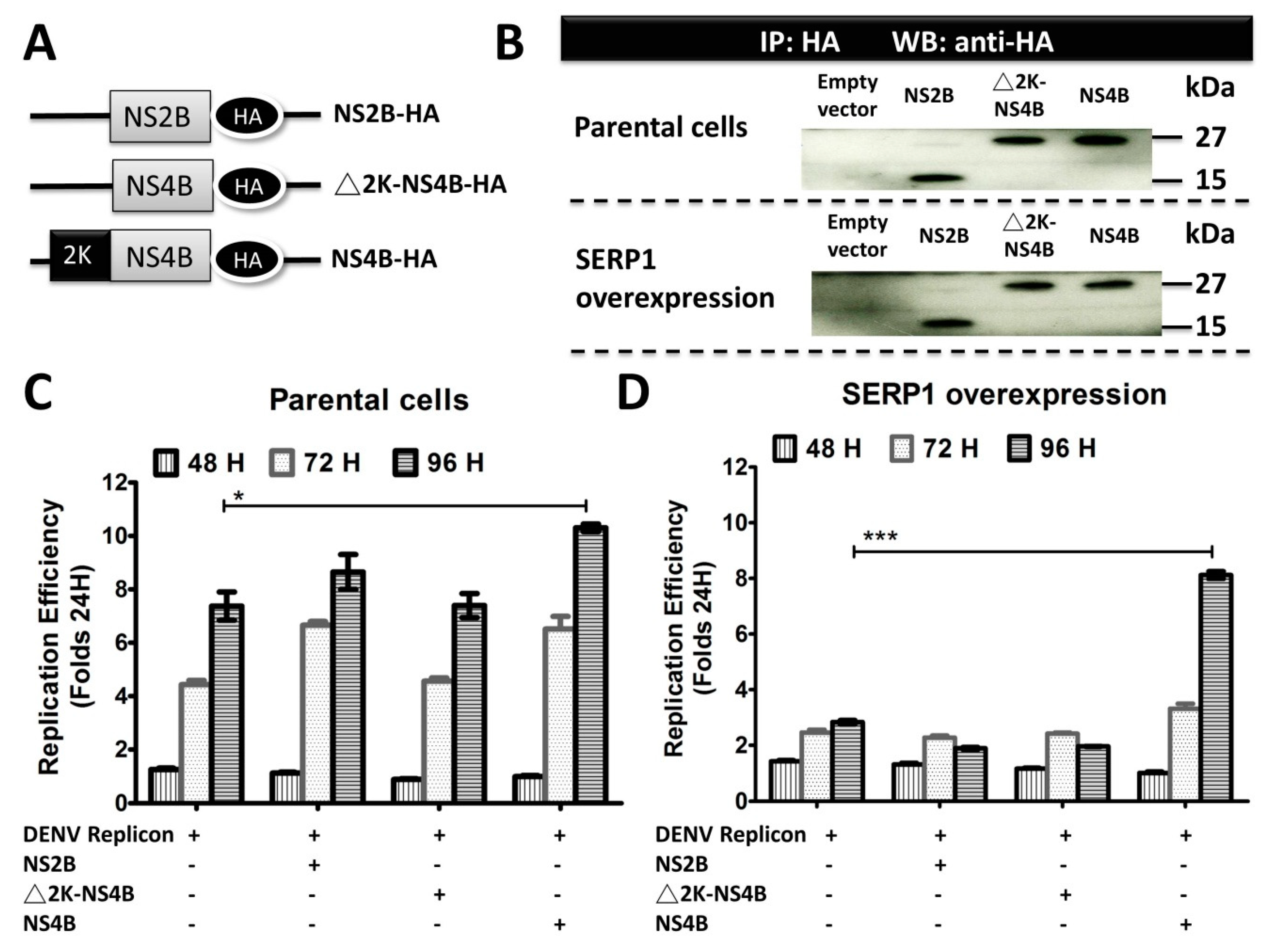
The DENV-2 replicon activity at 96 h post transfection was severely reduced in the SERP1-overexpressing Huh7.5 cells (
Figure 6D). Interestingly, the inhibitory effect of SERP1 on DENV-2 RNA replication in the Huh7.5 cells overexpressing SERP1 was alleviated by the overexpression of the NS4B protein. In contrast, the DENV-2 replicon activity in the Huh7.5 cells gradually increased from 48–96 h post-transfection in the Huh7.5 cells either co-transfected without or with the NS2B or NS4B protein (
Figure 6C). Specifically, overexpressing the NS4B protein in the parental Huh7.5 cells moderately increased the RNA replication compared with the Huh7.5 cells transfected only with DENV-2 replicon. We envisioned that it is likely that the amounts of endogenous SERP1 derived from the induction by the DENV-2 replicon replication are less than those of the SERP1 from stable cells overexpressing SERP1. The expression level of SERP1 determines the degree of inhibitory effect on the DENV-2 RNA replication. This phenomenon implied that NS4B may have different roles, in addition to being a component of the DENV-2 replication complex at the ER during DENV-2 replication.
What is the molecular mechanism by which NS4B counteracts the inhibitory effect of SERP1 on DENV-2 RNA replication? NS4B overexpression may induce cellular responses to prevent the antiviral activity derived from SERP1. A previous study indicated that NS4B is involved in interferon inhibition in DENV-infected cells, in order to prevent the establishment of a cellular antiviral state to enhance virus yields and replication [
13]. Huh7.5 cells exhibit a defect in IFN induction by dsRNA, because of a single point mutation in the dsRNA sensor retinoic acid-inducible gene-I (RIG-I) [
37]. Therefore, it is unlikely that IFN is responsible for the inhibitory effect of the SERP1 expression on DENV replication. DENV NS4B has also been identified as an important organizer of the membranous DENV replication complex, and as an inducer of intracellular membrane rearrangements [
14], which may result in cellular signaling to counteract the inhibitory effect of SERP1 expression on DENV replication. Alternatively, it is anticipated that the role of NS4B in facilitating replication may be as a SERP1 binder to counteract the antiviral role of SERP1. This scenario is supported by the observation that the DENV-2 NS4B–SERP1 interaction was suggested by the results of the MYTH system analysis, subcellular colocalization experiments, NanoBiT complementation assay, and co-immunoprecipitation assay (
Figure 1). However, the other possibility cannot be excluded that SERP1 impairs DENV-2 RNA replication via interrupting the interaction between NS4B and NS2B-NS3, as DENV-2 NS4B was reported to interact with NS2B-NS3, and the interaction is crucial for viral RNA replication [
38]. The overexpression of NS4B alleviates the inhibitory effect of SERP1 on DENV-2 RNA replication, possibly by restoring the binding of NS4B to NS2B-NS3 within the replication complex. Our previous attempt to show the interaction between SERP1 and NS2B-NS3 failed, because of the toxicity of the NS2B-NS3 overexpression in yeast, Huh7.5, and HEK-293 cells. Thus, whether SERP1 overexpression affects the interaction between DENV-2 NS4B and NS2B-NS3 cannot be evaluated in the cells overexpressing NS2B-NS3. Further studies are needed in order to evaluate the possibility. Given the fact that NS4B is a key component of the DENV-2 replication complex located in the ER, NS4B may play multiple roles in counteracting antiviral activity during DENV-2 replication, by interacting with various viral proteins and host factors.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






