Virus Shedding of Avian Influenza in Poultry: A Systematic Review and Meta-Analysis

 , , and
, , and 
Abstract
:1. Introduction
2. Methods
2.1. Systematic Review Methodology
2.2. Literature Search
2.3. Screening and Quality Assessment
- The publication is written in English;
- The study described is primary research;
- Influenza A virus is the subject being studied;
- The study was performed in poultry;
- Virus shedding is quantified.
- All criteria of the first screening are met;
- The subtype of the virus being studied is known;
- The pathogenicity classification of the virus (LPAIV or HPAIV) is known;
- The experimental units are confirmed negative for AIV in advance of the study;
- The virus inoculation route and dose are described;
- Virus shedding is quantified by PCR or by virus isolation;
- The sampling interval of virus shedding is described or;
- Peak virus shedding is described or can be determined from the data.
2.4. Data Extraction
2.5. Data Analysis and Summation
2.5.1. Qualitative Analysis
2.5.2. Quantitative Analysis (Meta-Analysis)
3. Results
3.1. Literature Search and Screening
3.2. Data Analysis and Summation
3.2.1. Qualitative Analysis
3.2.2. Quantitative Analysis (Meta-Analysis)
3.2.3. Shedding Levels
3.2.4. Duration of Shedding
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fouchier, R.A.; Munster, V.; Wallensten, A.; Bestebroer, T.M.; Herfst, S.; Smith, D.; Rimmelzwaan, G.F.; Olsen, B.; Osterhaus, A.D. Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. J. Virol. 2005, 79, 2814–2822. [Google Scholar] [CrossRef]
- Suarez, D.L. Influenza A virus. In Animal Influenza, 2nd ed.; Swayne, D.E., Ed.; John Wiley & Sons, Inc.: Ames, IA, USA, 2017; pp. 1–30. [Google Scholar]
- Sims, L.D.; Swayne, D.E. Avian influenza control strategies. In Animal Influenza, 2nd ed.; John Wiley & Sons, Inc.: Ames, IA, USA, 2016; pp. 363–377. [Google Scholar]
- Boni, M.F.; Galvani, A.P.; Wickelgren, A.L.; Malani, A. Economic epidemiology of avian influenza on smallholder poultry farms. Theor. Popul. Biol. 2013, 90, 135–144. [Google Scholar] [CrossRef] [Green Version]
- WHO Cumulative Number of Confirmed Human Cases of Avian Influenza A (H5N1) Reported to WHO. Available online: https://www.who.int/influenza/human_animal_interface/H5N1_cumulative_table_archives/en/ (accessed on 15 December 2018).
- Kalthoff, D.; Globig, A.; Beer, M. (Highly pathogenic) avian influenza as a zoonotic agent. Vet. Microbiol. 2010, 140, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Tanner, W.D.; Toth, D.J.; Gundlapalli, A.V. The pandemic potential of avian influenza A(H7N9) virus: A review. Epidemiol. Infect. 2015, 143, 3359–3374. [Google Scholar] [CrossRef]
- Humberd, J.; Boyd, K.; Webster, R.G. Emergence of influenza A virus variants after prolonged shedding from pheasants. J. Virol. 2007, 81, 4044–4051. [Google Scholar] [CrossRef]
- Bertran, K.; Lee, D.; Balzli, C.; Pantin-Jackwood, M.J.; Spackman, E.; Swayne, D.E. Age is not a determinant factor in susceptibility of broilers to H5N2 clade 2.3.4.4 high pathogenicity avian influenza virus. Vet. Res. 2016, 47. [Google Scholar] [CrossRef]
- Mondal, S.; Xing, Z.; Cardona, C. A comparison of virulence of influenza A virus isolates from mallards in experimentally inoculated turkeys. Avian Dis. 2013, 57, 790–796. [Google Scholar] [CrossRef]
- Guionie, O.; Guillou-Cloarec, C.; Courtois, D.; Bougeard, B.S.; Amelot, M.; Jestin, V. Experimental infection of Muscovy ducks with highly pathogenic avian influenza virus (H5N1) belonging to clade 2.2. Avian Dis. 2010, 54, 538–547. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J.; Smith, D.M.; Wasilenko, J.L.; Spackman, E. Low pathogenicity avian influenza viruses infect chicken layers by different routes of inoculation. Avian Dis. 2012, 56, 276–281. [Google Scholar] [CrossRef]
- Chaves, A.J.; Busquets, N.; Campos, N.; Ramis, A.; Dolz, R.; Rivas, R.; Valle, R.; Abad, F.X.; Darji, A.; Majo, N. Pathogenesis of highly pathogenic avian influenza A virus (H7N1) infection in chickens inoculated with three different doses. Avian Pathol. 2011, 40, 163–172. [Google Scholar] [CrossRef]
- Das, B.R.; Kumar, M.; Murugkar, H.V.; Nagarajan, S.; Kumar, D.S.; Kalaiyarasu, S.; Kulkarni, D.D.; Tosh, C. Experimental inoculation of a crow derived influenza A (H5N1) virus in chickens and its pathological and genetic characterization. Indian J. Anim. Sci. 2016, 86, 238–242. [Google Scholar]
- Van der Goot, J.A.; Koch, G.; de Jong, M.C.; van Boven, M. Quantification of the effect of vaccination on transmission of avian influenza (H7N7) in chickens. Proc. Natl. Acad. Sci. USA 2005, 102, 18141–18146. [Google Scholar] [CrossRef] [Green Version]
- Sitaras, I.; Rousou, X.; Kalthoff, D.; Beer, M.; Peeters, B.; de Jong, M.C. Role of vaccination-induced immunity and antigenic distance in the transmission dynamics of highly pathogenic avian influenza H5N1. J. R. Soc. Interface 2016, 13, 20150976. [Google Scholar] [CrossRef]
- Spackman, E.; Pantin-Jackwood, M.J.; Kapczynski, D.R.; Swayne, D.E.; Suarez, D.L. H5N2 Highly Pathogenic Avian Influenza Viruses from the US 2014-2015 outbreak have an unusually long pre-clinical period in turkeys. BMC Vet. Res. 2016, 12, 260. [Google Scholar] [CrossRef]
- Ge, F.; Xin, L.; Houbin, J.; Dequan, Y.; Jian, L.; Xinyong, Q.; Jian, W.; Xianchao, Y.; Yafeng, Q.; Peihong, L.; et al. Genotypic evolution and antigenicity of H9N2 influenza viruses in Shanghai, China. Arch. Virol. 2016, 161, 1437–1445. [Google Scholar] [CrossRef]
- Jiao, P.; Song, H.; Liu, X.; Song, Y.; Cui, J.; Wu, S.; Ye, J.; Qu, N.; Zhang, T.; Liao, M. Pathogenicity, Transmission and Antigenic Variation of H5N1 Highly Pathogenic Avian Influenza Viruses. Front. Microbiol. 2016, 7, 635. [Google Scholar] [CrossRef]
- Kapczynski, D.R.; Tumpey, T.M.; Hidajat, R.; Zsak, A.; Chrzastek, K.; Tretyakova, I.; Pushko, P. Vaccination with virus-like particles containing H5 antigens from three H5N1 clades protects chickens from H5N1 and H5N8 influenza viruses. Vaccine 2016, 34, 1575–1581. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority, Application of systematic review methodology to food and feed safety assessments to support decision making. EFSA J. 2010, 8, 1637–1727. [CrossRef]
- O’Connor, A.M.; Sargeant, J.M. An introduction to systematic reviews in animal health, animal welfare, and food safety. Anim. Health Res. Rev. 2014, 15, 3–13. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Bruggeman PlotReader Tool. 2010. Available online: https://jornbr.home.xs4all.nl/plotreader/ (accessed on 2 June 2017).
- Gonzales, J.L. Wageningen Bioveterinary Research, Lelystad, the Netherlands. the assessment of similarity between measured shedding levels by virus isolation and real time PCR. 2018; (unpublished). [Google Scholar]
- Gonzales, J.L.; Elbers, A.R.; Bouma, A.; Koch, G.; de Wit, J.J.; Stegeman, J.A. Transmission characteristics of low pathogenic avian influenza virus of H7N7 and H5N7 subtypes in layer chickens. Vet. Microbiol. 2012, 155, 207–213. [Google Scholar] [CrossRef]
- Bouma, A.; Claassen, I.; Natih, K.; Klinkenberg, D.; Donnelly, C.A.; Koch, G.; van Boven, M. Estimation of transmission parameters of H5N1 avian influenza virus in chickens. PLoS Pathog. 2009, 5, e1000281. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 1 Novermber 2017).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Therneau, T. A Package for Survival Analysis in S version 2.38. 2015. Available online: https://cran.r-project.org/web/packages/survival/index.html (accessed on 1 December 2017).
- Guan, J.; Fu, Q.; Sharif, S. Replication of an H9N2 Avian Influenza Virus and Cytokine Gene Expression in Chickens Exposed by Aerosol or Intranasal Routes. Avian Dis. 2015, 59, 263–268. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.; Swayne, D.E.; Smith, D.; Shepherd, E. Effect of species, breed and route of virus inoculation on the pathogenicity of H5N1 highly pathogenic influenza (HPAI) viruses in domestic ducks. Vet. Res. 2013, 44, 62. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Gonzales, J.L.; Koch, G.; Elbers, A.R.W.; van der Goot, J.A. Similar transmissibility of the Italian H7N1 highly pathogenic avian influenza virus and its low pathogenic avian influenza virus predecessor. Vet. J. 2018, 232, 20–22. [Google Scholar] [CrossRef]
- Spekreijse, D.; Bouma, A.; Stegeman, J.A.; Koch, G.; de Jong, M.C. The effect of inoculation dose of a highly pathogenic avian influenza virus strain H5N1 on the infectiousness of chickens. Vet. Microbiol. 2011, 147, 59–66. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J.; Stephens, C.B.; Bertran, K.; Swayne, D.E.; Spackman, E. The pathogenesis of H7N8 low and highly pathogenic avian influenza viruses from the United States 2016 outbreak in chickens, turkeys and mallards. PLoS ONE 2017, 12, e0177265. [Google Scholar] [CrossRef]
- Bertran, K.; Lee, D.-H.; Criado, M.F.; Smith, D.; Swayne, D.E.; Pantin-Jackwood, M.J. Pathobiology of Tennessee 2017 H7N9 low and high pathogenicity avian influenza viruses in commercial broiler breeders and specific pathogen free layer chickens. Vet. Res. 2018, 49, 82. [Google Scholar] [CrossRef]
- Swayne, D.S.; Slemons, R.D. Comparative pathology of a chicken-origin and two duck-origin influenza virus isolates in chickens: The effect of route of inoculation. Vet. Pathol. 1994, 21, 237–245. [Google Scholar] [CrossRef]
- Webster, R.G.; Yakhno, M.; Hinshaw, V.S.; Bean, W.J.; Murti, K.G. Intestinal influenza: Replication and characterization of influenza viruses in ducks. Virology 1978, 84, 268–278. [Google Scholar] [CrossRef]
- Carranza-Flores, J.M.; Padilla-Noriega, L.; Loza-Rubio, E.; Garcia-Espinosa, G. Prolonged excretion of a low-pathogenicity H5N2 avian influenza virus strain in the Pekin duck. J. Vet. Sci. 2013, 14, 487–490. [Google Scholar] [CrossRef]
- Slemons, R.D.; Easterday, B.C. The Natural History of Type-A Influenza Viruses and Wild Waterfowl. In Wildlife Diseases; Page, L.A., Ed.; Springer: Boston, MA, USA, 1976; pp. 215–224. [Google Scholar]
- Pantin-Jackwood, M.J.; Swayne, D.E. Pathogenesis and pathobiology of avian influenza virus infection in birds. Rev. Sci. Tech. (Int. Off. Epizoot.) 2009, 28, 113–136. [Google Scholar] [CrossRef]
- Pillai, S.P.; Pantin-Jackwood, M.; Suarez, D.L.; Saif, Y.M.; Lee, C.W. Pathobiological characterization of low-pathogenicity H5 avian influenza viruses of diverse origins in chickens, ducks and turkeys. Arch. Virol. 2010, 155, 1439–1451. [Google Scholar] [CrossRef]
- Spackman, E.; Gelb, J., Jr.; Preskenis, L.A.; Ladman, B.S.; Pope, C.R.; Pantin-Jackwood, M.J.; McKinley, E.T. The pathogenesis of low pathogenicity H7 avian influenza viruses in chickens, ducks and turkeys. Virol. J. 2010, 7, 331. [Google Scholar] [CrossRef]
- Saenz, R.A.; Essen, S.C.; Brookes, S.M.; Iqbal, M.; Wood, J.L.; Grenfell, B.T.; McCauley, J.W.; Brown, I.H.; Gog, J.R. Quantifying transmission of highly pathogenic and low pathogenicity H7N1 avian influenza in turkeys. PLoS ONE 2012, 7, e45059. [Google Scholar] [CrossRef]
- Lavoie, E.T.; Sorrell, E.M.; Perez, D.R.; Ottinger, M.A. Immunosenescence and age-related susceptibility to influenza virus in Japanese quail. Dev. Comp. Immunol. 2007, 31, 407–414. [Google Scholar] [CrossRef]
- VanDalen, K.K.; Franklin, A.B.; Mooers, N.L.; Sullivan, H.J.; Shriner, S.A. Shedding light on avian influenza H4N6 infection in mallards: Modes of transmission and implications for surveillance. PLoS ONE 2010, 5, e12851. [Google Scholar] [CrossRef]
- Costa, T.P.; Brown, J.D.; Howerth, E.W.; Stallknecht, D.E. The effect of age on avian influenza viral shedding in mallards (Anas platyrhynchos). Avian Dis. 2010, 54, 581–585. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
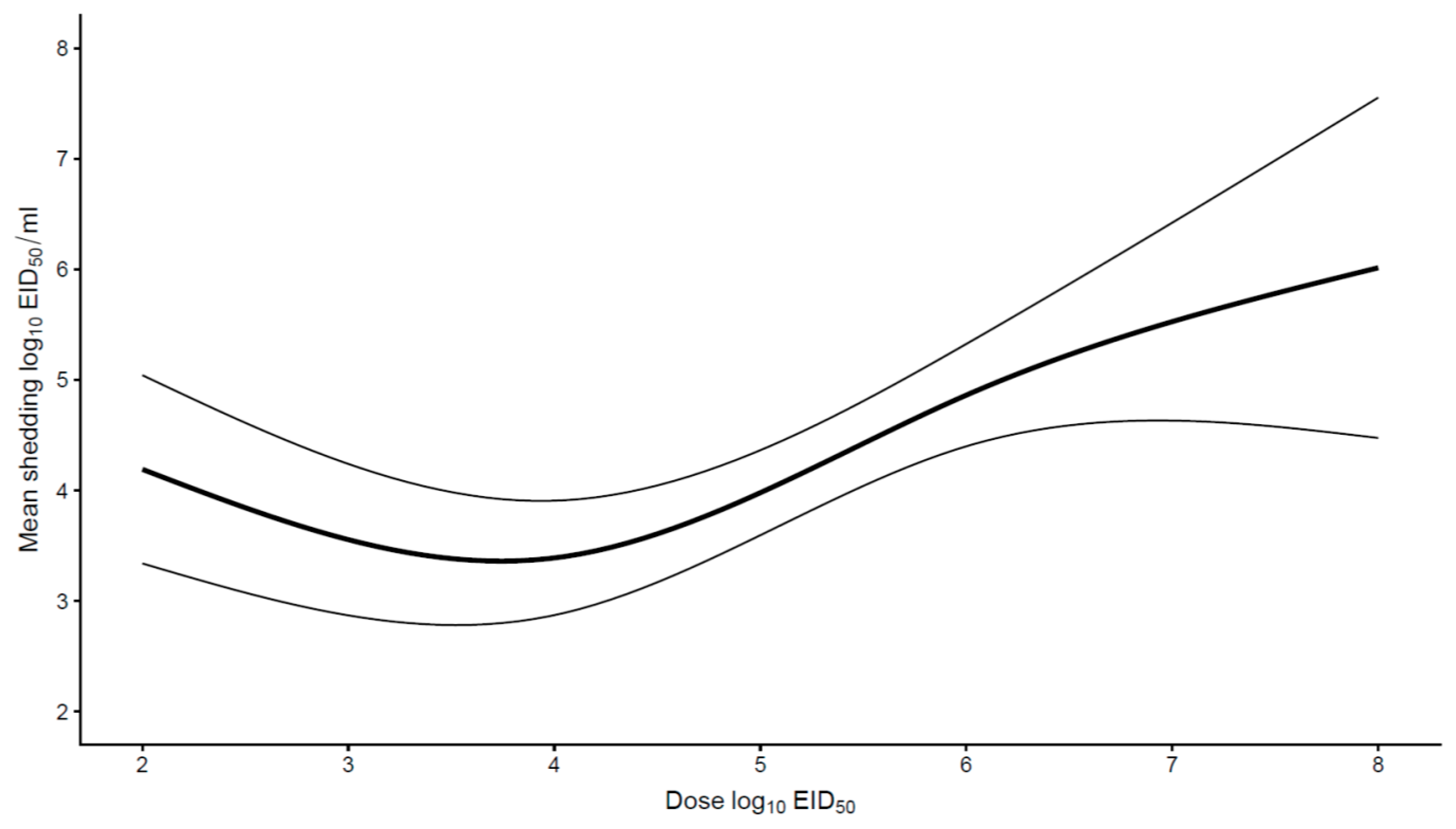{kind=link}
| Explanatory Variables | Levels | Number of Observations | Percentage of Observations (%) | ||
|---|---|---|---|---|---|
| HPAIV | LPAIV | HPAIV | LPAIV | ||
| Species The poultry species that were infected | Chicken | 484 | 477 | 86.43 | 65.3 |
| Duck | 66 | 112 | 11.79 | 15.3 | |
| Turkey | 8 | 108 | 1.43 | 14.8 | |
| Goose | 2 | 2 | 0.36 | 0.3 | |
| Pigeon | 8 | 1.1 | |||
| Quail | 16 | 2.2 | |||
| Guinea fowl | 4 | 0.5 | |||
| Pheasant | 3 | 0.4 | |||
| Age a The age at infection | Adult | 70 | 109 | 12.50 | 14.9 |
| Young | 490 | 621 | 87.50 | 85.1 | |
| Inoculation route Site at which the animal is experimentally inoculated with AIV | Aerosol | 4 | 0.5 | ||
| Contact | 17 | 231 | 3.04 | 31.6 | |
| Intrachoanal | 119 | 191 | 21.25 | 26.2 | |
| Intracloacal | 4 | 0.5 | |||
| Intranasal (combi) | 424 | 276 | 75.71 | 37.8 | |
| Intraoviduct | 4 | 0.5 | |||
| Oropharyngeal | 20 | 2.7 | |||
| Subtype Subtype of the inoculated virus | H3N2 | 4 | 0.5 | ||
| H5N1 | 349 | 11 | 62.77 | 1.5 | |
| H5N2 | 165 | 239 | 29.68 | 32.7 | |
| H5N3 | 42 | 5.8 | |||
| H5N5 | 11 | 1.5 | |||
| H5N6 | 4 | 0.8 | |||
| H5N7 | 6 | ||||
| H5N8 | 16 | 9 | 2.88 | 1.2 | |
| H5N9 | 11 | 1.5 | |||
| H6N2 | 38 | 5.2 | |||
| H7N1 | 16 | 104 | 2.88 | 14.2 | |
| H7N2 | 2 | 54 | 0.36 | 7.4 | |
| H7N3 | 6 | 18 | 1.08 | 2.5 | |
| H7N7 | 2 | 72 | 0.36 | 9.9 | |
| H7N8 | 6 | 0.8 | |||
| H7N9 | 64 | 8.8 | |||
| H9N2 | 38 | 5.2 | |||
| H10N9 | 3 | 0.4 | |||
| Virus origin Species from which the virus originates | Same species | 146 | 392 | 26.07 | 53.7 |
| Same phylogenetic order | 333 | 95 | 59.46 | 13.0 | |
| Different phylogenetic order | 81 | 225 | 14.46 | 30.8 | |
| Origin unknown | 18 | 2.5 | |||
| Sample site Body part where a sample was taken for determination of the virus shedding levels | Buccal | 5 | 4 | 0.89 | 0.5 |
| Cloacal | 238 | 357 | 42.50 | 48.9 | |
| Tracheal | 91 | 181 | 16.25 | 24.8 | |
| Oropharyngeal | 183 | 187 | 32.68 | 25.6 | |
| Nasal | 1 | 1 | 0.18 | 0.1 | |
| Oral | 38 | 6.79 | |||
| Laryngopharyngeal | 4 | 0.71 | |||
| Type | Layer | 418 | 177 | 86.36 | 37.11 |
| Production type of commercial chickens | Broiler | 28 | 3 | 5.79 | 0.63 |
| Unknown | 38 | 297 | 7.85 | 62.26 | |
| Explanatory Variables | Poultry Species | Effect | Difference in Mean Shedding (log10 EID50/mL) a | Difference in Peak Shedding (log10 EID50/mL) a | p (Mean|Peak) |
|---|---|---|---|---|---|
| Pathotype (HPAI vs. LPAI) | All | Poultry infected with LPAI shed lower levels of virus than poultry infected with HPAI | −1.82 ± 0.33 | −3.14 ± 0.51 | <0.001|<0.001 |
| Shedding route | All | Respiratory shedding is higher than cloacal shedding for all species | 0.43 ± 0.12 | 0.42 ± 0.15 | <0.001|0.005 |
| Turkeyand goose | NS differences between turkeys and geese with chickens | NS | NS | >0.05|>0.05 | |
| Duck | Higher respiratory shedding than chickens | 0.58 ± 0.35 | 0.75 ± 0.39 | <0.001|<0.001 | |
| Lower cloacal shedding than chickens | −1.02 ± 0.38 | −0.93 ± 0.43 | 0.008|0.03 | ||
| Higher respiratory shedding than cloacal | 1.96 ± 0.17 | 2.10 ± 0.31 | <0.001|<0.001 | ||
| Virus origin | Chicken | Shedding is lower when virus originates from birds from the same order than when the virus originates from the same species | −0.89 ± 0.25 | −0.62 ± 0.30 | <0.001|0.04 |
| Shedding is lower when virus originates from birds from different orders than when the virus originates from the same species | −0.92 ± 0.29 | −0.93 ± 0.34 | <0.002|0.007 | ||
| Duck | NS differences in shedding between the origin of the virus | NS | NS | >0.05|>0.05 | |
| Inoculation route | All | NS differences in shedding between intranasal and intrachoanal routes versus contact infected poultry b | NS | NS | >0.05|>0.05 |
| Age | All | Young poultry shed lower than adults c,d | −1.07 ± 0.21 | −1.03 ± 0.25 | <0.001|<0.001 |
| Type | Chicken | NS differences in shedding between broilers and layers | NS | NS | >0.05|>0.05 |
| Explanatory Variables | Poultry Species | Effect | Difference in Mean Shedding (log10 EID50/mL) | Difference in Peak Shedding (log10 EID50/mL) | p (Mean|Peak) |
|---|---|---|---|---|---|
| Shedding route | All | Respiratory shedding is higher than cloacal shedding with the exception of ducks and pigeons | 0.70 ± 0.11 | 2.15 ± 0.14 | <0.001|<0.001 |
| Duck | Higher cloacal shedding than chickens | 0.65 ± 0.17 | 1.19 ± 0.27 | <0.001|<0.001 | |
| NS difference between cloacal and respiratory shedding | NS | NS | >0.05|>0.05 | ||
| Turkey | Higher cloacal shedding than chickens | 0.36 ± 0.17 | 1.16 ± 0.27 | <0.001|<0.001 | |
| Pigeon | Lower respiratory and cloacal shedding than chickens | −1.21 ± 0.59 | −2.42 ± 0.83 | 0.042|0.003 | |
| Quail | Higher respiratory shedding than chickens | 1.19 ± 0.44 | 2.42 ± 0.65 | 0.007|<0.001 | |
| Guinea fowl | NS differences in shedding between chickens and guinea fowl | NS | NS | >0.05|>0.05 | |
| Virus origin | Chicken | Shedding is lower when the virus originates from birds from the same order than when the virus originates from the same species | −1.17 ± 0.28 | −1.35 ± 0.78 | <0.001|0.01 |
| Shedding is lower when the virus originates from birds from different orders than when the virus originates from the same species | −0.73 ± 0.27 | −2.08 ± 0.67 | 0.007|0.005 | ||
| Duck | NS differences in shedding between the origin of the virus | NS | NS | >0.05|>0.05 | |
| Turkey | NS differences in shedding between the origin of the virus | NS | NS | >0.05|>0.05 | |
| Inoculation route | Chicken | Aerosol inoculated chickens shed higher than contact infected chickens | 2.04 ± 0.66 | 2.72 ± 1.04 | 0.002|0.009 |
| All | Intranasal inoculated poultry shed higher than contact infected poultry | 0.61 ± 0.14 | 0.77 ± 0.71 | <0.001|<0.001 | |
| All | Oropharyngeal inoculated poultry shed higher than contact infected poultry | 2.44 ± 1.13 | 2.66 ± 1.13 | 0.042|0.03 | |
| All | NS differences in shedding between intrachoanal, intracloacal and intra-oviduct inoculation routes and contact infected poultry | NS | NS | >0.05|>0.05 | |
| Age | Chicken | NS differences in shedding between ages a | NS | NS | >0.05|>0.05 |
| Type | Chicken | NS differences in shedding between broilers and layers b | NS | NS | >0.05|>0.05 |
| Pathotype | Poultry Species | Length of Respiratory Virus Shedding (Days) | Length of Cloacal Virus Shedding (Days) | ||
|---|---|---|---|---|---|
| HPAI | Chicken | 2.6 | (1.1–6.5) | 2.5 | (1.0–6.2) |
| Duck | 6.9 | (2.8–17.1) | 6.6 | (2.7–16.3) | |
| LPAI | Chicken | 6.2 | (0.8–17.8) | 5.5 | (0.7–15.7) |
| Duck | 5.3 | (0.7–15.3) | 8.2 | (1.0–23.3) | |
| Turkey | 10.0 | (1.3–28.7) | 14.1 | (1.8–40.2) | |
| Guinea fowl | 3.3 | (0.4–9.4) | 3.3 | (0.4–9.4) | |
| Pigeon | 3.6 | (0.4–10.2) | 2.8 | (0.3–8.0) | |
| Quail | NA | 6.9 | (0.9–19.8) | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germeraad, E.A.; Sanders, P.; Hagenaars, T.J.; de Jong, M.C.M.; Beerens, N.; Gonzales, J.L. Virus Shedding of Avian Influenza in Poultry: A Systematic Review and Meta-Analysis. Viruses 2019, 11, 812. https://doi.org/10.3390/v11090812
Germeraad EA, Sanders P, Hagenaars TJ, de Jong MCM, Beerens N, Gonzales JL. Virus Shedding of Avian Influenza in Poultry: A Systematic Review and Meta-Analysis. Viruses. 2019; 11(9):812. https://doi.org/10.3390/v11090812
Chicago/Turabian StyleGermeraad, Evelien A., Pim Sanders, Thomas J. Hagenaars, Mart C.M. de Jong, Nancy Beerens, and Jose L. Gonzales. 2019. "Virus Shedding of Avian Influenza in Poultry: A Systematic Review and Meta-Analysis" Viruses 11, no. 9: 812. https://doi.org/10.3390/v11090812
APA StyleGermeraad, E. A., Sanders, P., Hagenaars, T. J., de Jong, M. C. M., Beerens, N., & Gonzales, J. L. (2019). Virus Shedding of Avian Influenza in Poultry: A Systematic Review and Meta-Analysis. Viruses, 11(9), 812. https://doi.org/10.3390/v11090812