Genetic, Morphological and Antigenic Relationships between Mesonivirus Isolates from Australian Mosquitoes and Evidence for Their Horizontal Transmission

 , , , ,
, , , ,  , , , ,
, , , ,  add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Virus Isolation
2.2. Detection and Isolation of Mesoniviruses from Homogenised Mosquito Pools
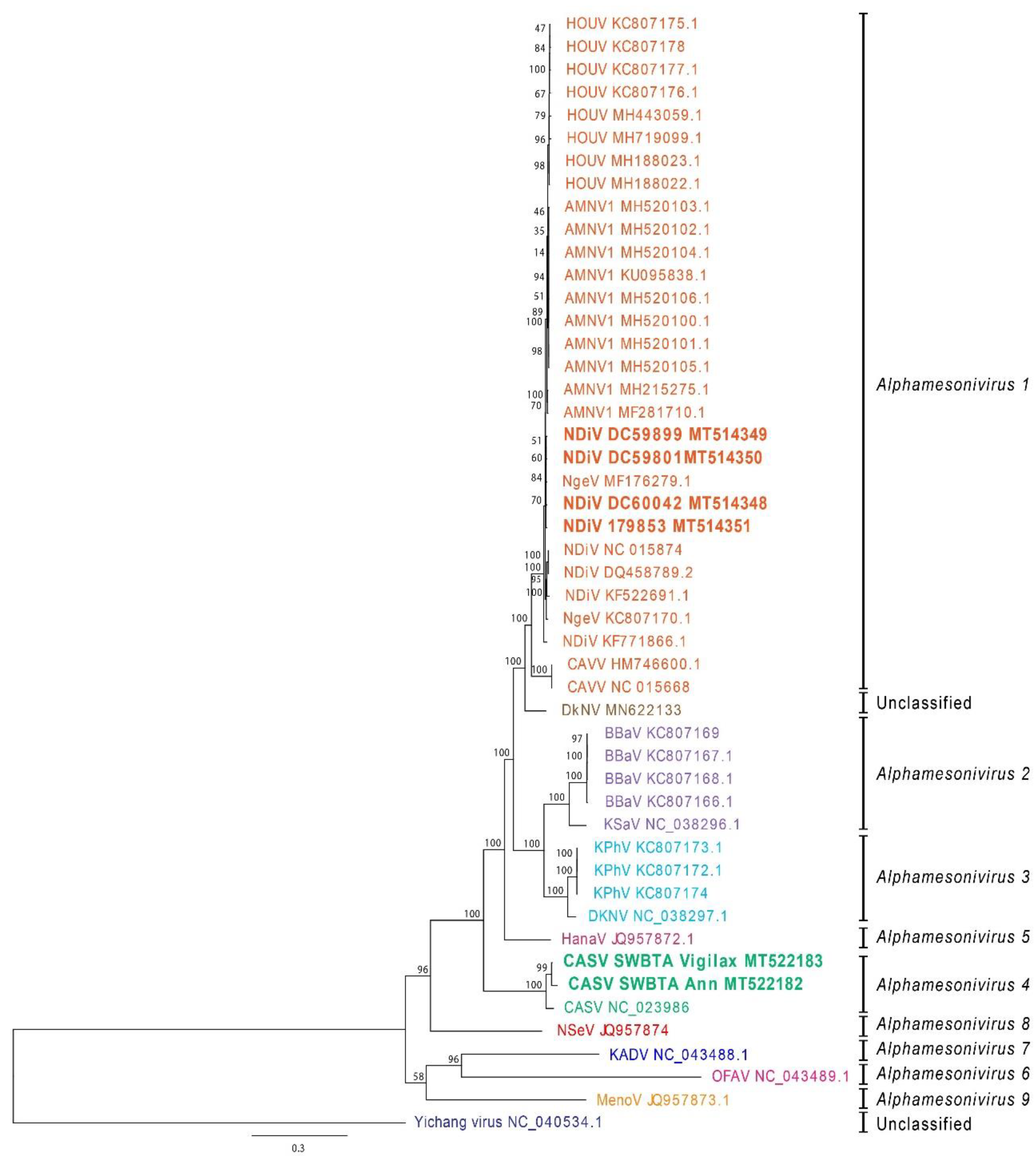
2.3. Genome Sequencing and Phylogenetic Analysis
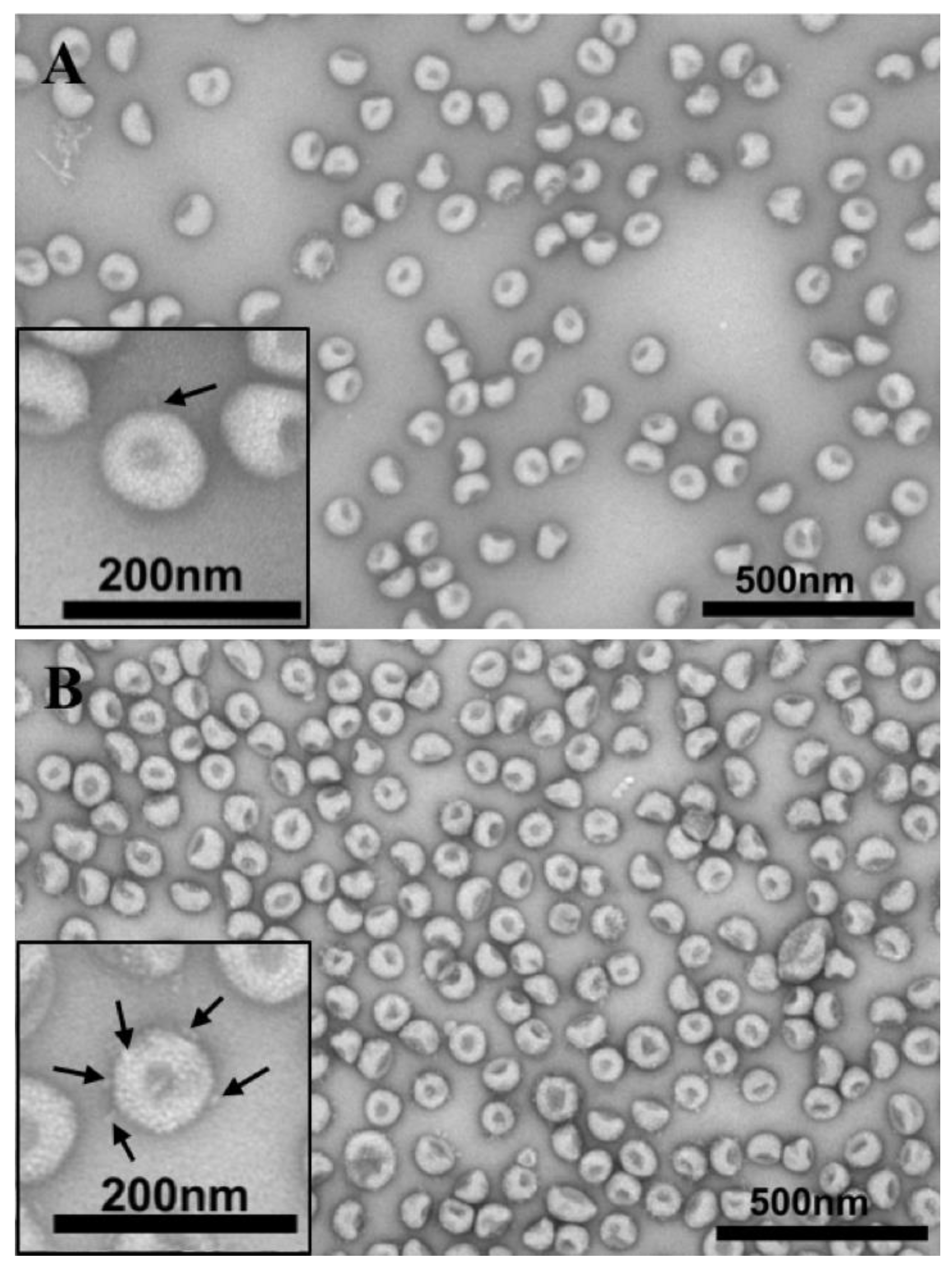
2.4. Electron Microscopy—Purification and TEM
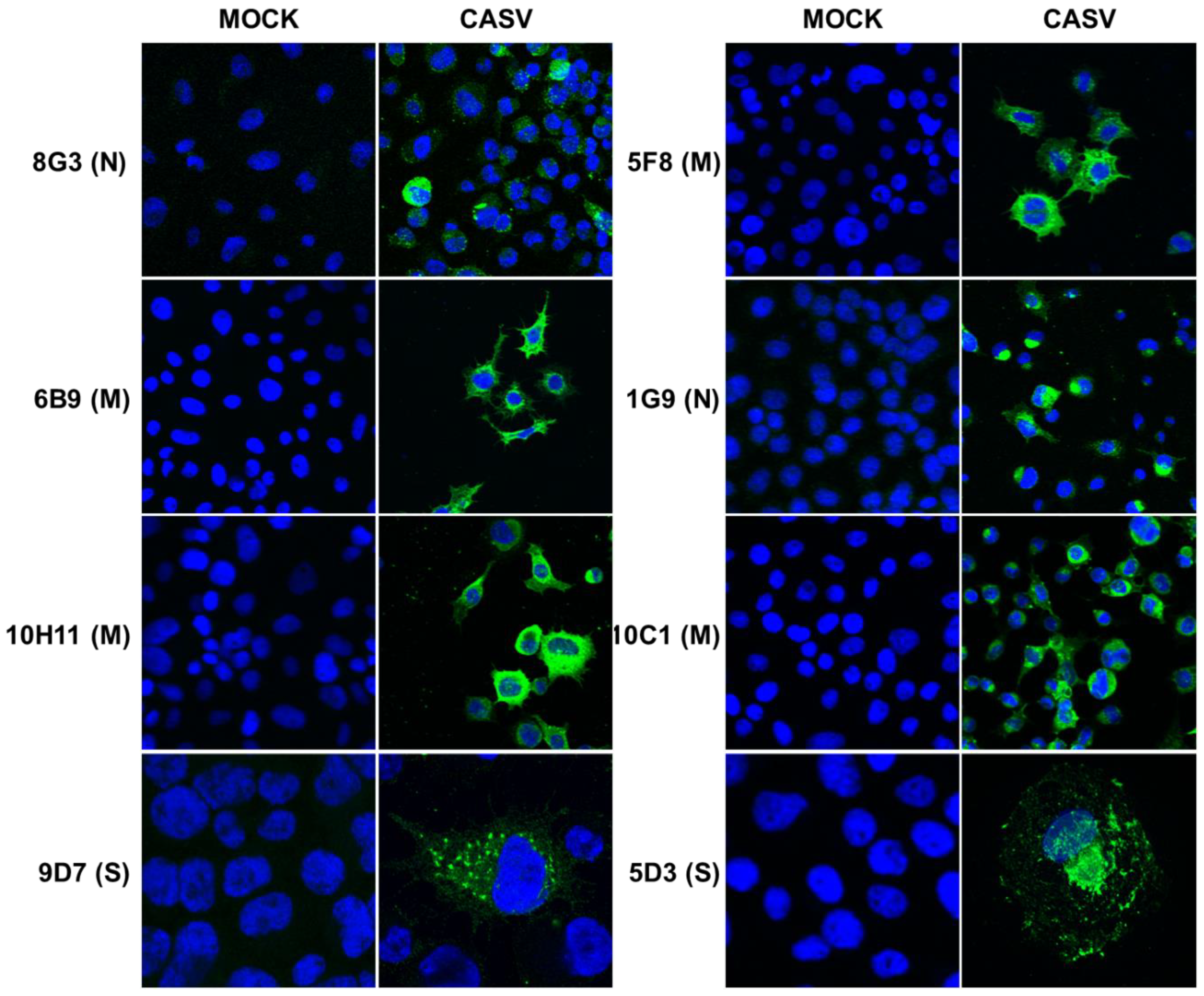
2.5. Preparation of Mouse Antiserum and Monoclonal Antibodies to CASV
2.6. SDS-PAGE, Western Blot and Immunoprecipitation
2.7. Microneutralisation Assay
2.8. RT-PCR
2.9. Viral RNA Extraction from Mosquito Saliva on Flinders Technology Associates (FTA) Cards
2.10. Deep Sequencing of Viral RNA Eluted from FTA Cards
3. Results
3.1. Isolation of Two Mesoniviruses from Multiple Mosquito Species in Australia
3.2. Comparison of Virion Morphology of CASV and NDiV by Electron Microscopy
3.3. Production of Monoclonal Antibodies to CASV and NDiV
3.4. Detection of Mesonivirus RNA in Mosquito Saliva during Routine Arbovirus Surveillance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pasternak, A.O.; Spaan, W.J.M.; Snijder, E.J. Nidovirus transcription: How to make sense? J. Gen. Virol. 2006, 87, 1403–1421. [Google Scholar] [CrossRef]
- Lauber, C.; Goeman, J.J.; Parquet, M.D.C.; Nga, P.T.; Snijder, E.J.; Morita, K.; Gorbalenya, A.E. The footprint of genome architecture in the largest genome expansion in RNA viruses. PLOS Pathog. 2013, 9, e1003500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warrilow, D.; Watterson, D.; Hall, R.A.; Davis, S.S.; Weir, R.; Kurucz, N.; Whelan, P.; Allcock, R.; Hall-Mendelin, S.; O’Brien, C.A.; et al. A new species of Mesonivirus from the Northern Territory, Australia. PLoS ONE 2014, 9, e91103. [Google Scholar] [CrossRef]
- Zirkel, F.; Roth, H.; Kurth, A.; Drosten, C.; Ziebuhr, J.; Junglen, S. Identification and characterization of genetically divergent members of the newly established family Mesoniviridae. J. Virol. 2013, 87, 6346–6358. [Google Scholar] [CrossRef] [Green Version]
- Nga, P.T.; Parquet, M.D.C.; Lauber, C.; Parida, M.; Nabeshima, T.; Yu, F.; Thuy, N.T.; Inoue, S.; Ito, T.; Okamoto, K.; et al. Discovery of the first insect Nidovirus, a missing evolutionary link in the emergence of the largest RNA virus genomes. PLOS Pathog. 2011, 7, e1002215. [Google Scholar] [CrossRef] [PubMed]
- Zirkel, F.; Kurth, A.; Quan, P.-L.; Briese, T.; Ellerbrok, H.; Pauli, G.; Leendertz, F.H.; Lipkin, W.I.; Ziebuhr, J.; Drosten, C.; et al. An insect Nidovirus emerging from a primary tropical rainforest. mBio 2011, 2, 00077-11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ICTV. ICTV Master Species List 2018b.v2. 2018. Available online: https://talk.ictvonline.org/files/master-species-lists/m/msl/8266 (accessed on 13 October 2020).
- Lauber, C.; Ziebuhr, J.; Junglen, S.; Drosten, C.; Zirkel, F.; Nga, P.T.; Morita, K.; Snijder, E.J.; Gorbalenya, A.E. Mesoniviridae: A proposed new family in the order Nidovirales formed by a single species of mosquito-borne viruses. Arch. Virol. 2012, 157, 1623–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorbalenya, A.E.; Enjuanes, L.; Ziebuhr, J.; Snijder, E.J. Nidovirales: Evolving the largest RNA virus genome. Virus Res. 2006, 117, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Hon, C.C.; Yip, C.W.; Law, K.M.; Yeung, Y.S.; Chan, K.H.; Peiris, J.S.M.; Leung, F.C. Quantitative comparison of the efficiency of antibodies against S1 and S2 subunit of SARS coronavirus spike protein in virus neutralization and blocking of receptor binding: Implications for the functional roles of S2 subunit. FEBS Lett. 2006, 580, 5612–5620. [Google Scholar] [CrossRef] [Green Version]
- Buchholz, U.J.; Bukreyev, A.; Yang, L.; Lamirande, E.W.; Murphy, B.R.; Subbarao, K.; Collins, P.L. Contributions of the structural proteins of severe acute respiratory syndrome coronavirus to protective immunity. Proc. Natl. Acad. Sci. USA 2004, 101, 9804–9809. [Google Scholar] [CrossRef] [Green Version]
- Bolling, B.G.; Eisen, L.; Moore, C.G.; Blair, C.D. Insect-specific flaviviruses from culex mosquitoes in Colorado, with evidence of vertical transmission. Am. J. Trop. Med. Hyg. 2011, 85, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras-Gutierrez, M.A.; Guzman, H.; Thangamani, S.; Vasilakis, N.; Tesh, R.B. Experimental infection with and maintenance of cell fusing agent virus (Flavivirus) in Aedes aegypti. Am. J. Trop. Med. Hyg. 2017, 97, 299–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prow, N.A.; Mah, M.G.; Deerain, J.M.; Warrilow, D.; Colmant, A.M.G.; O’Brien, C.A.; Harrison, J.J.; McLean, B.J.; Hewlett, E.K.; Piyasena, T.B.H.; et al. New genotypes of Liao ning virus (LNV) in Australia exhibit an insect-specific phenotype. J. Gen. Virol. 2018, 99, 596–609. [Google Scholar] [CrossRef] [PubMed]
- Saiyasombat, R.; Bolling, B.G.; Brault, A.C.; Bartholomay, L.C.; Blitvich, B.J. Evidence of efficient transovarial transmission of Culex flavivirus by Culex pipiens (Diptera: Culicidae). J. Med. Ѐntomol. 2011, 48, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Camp, J.V.; Bakonyi, T.; Soltész, Z.; Zechmeister, T.; Nowotny, N. Uranotaenia unguiculata Edwards, 1913 are attracted to sound, feed on amphibians, and are infected with multiple viruses. Parasites Vectors 2018, 11, 456. [Google Scholar] [CrossRef] [Green Version]
- Diagne, M.M.; Gaye, A.; Ndione, M.H.D.; Faye, M.; Fall, G.; Dieng, I.; Widen, S.G.; Wood, T.G.; Popov, V.; Guzman, H.; et al. Dianke virus: A new mesonivirus species isolated from mosquitoes in Eastern Senegal. Virus Res. 2019, 275, 197802. [Google Scholar] [CrossRef] [PubMed]
- Gaye, A.; Diagne, M.M.; Ndiaye, E.H.; Ndione, M.H.D.; Faye, M.; Talla, C.; Fall, G.; Ba, Y.; Diallo, D.; Dia, I.; et al. Vector competence of anthropophilic mosquitoes for a new mesonivirus in Senegal. Emerg. Microbes Infect. 2020, 9, 496–504. [Google Scholar] [CrossRef] [Green Version]
- Birnberg, L.; Temmam, S.; Aranda, C.; Correa-Fiz, F.; Talavera, S.; Bigot, T.; Eloit, M.; Busquets, N. Viromics on honey-baited FTA Cards as a new tool for the detection of circulating viruses in mosquitoes. Viruses 2020, 12, 274. [Google Scholar] [CrossRef] [Green Version]
- McLean, B.J.; Hobson-Peters, J.; Webb, C.E.; Watterson, D.; Prow, N.A.; Nguyen, H.D.; Hall-Mendelin, S.; Warrilow, D.; Johansen, C.A.; Jansen, C.C.; et al. A novel insect-specific flavivirus replicates only in Aedes-derived cells and persists at high prevalence in wild Aedes vigilax populations in Sydney, Australia. Virology 2015, 486, 272–283. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- O’Brien, C.A.; Hobson-Peters, J.; Yam, A.W.Y.; Colmant, A.M.G.; McLean, B.J.; Prow, N.A.; Watterson, D.; Hall-Mendelin, S.; Warrilow, D.; Ng, M.-L.; et al. Viral RNA intermediates as targets for detection and discovery of novel and emerging mosquito-borne viruses. PLOS Negl. Trop. Dis. 2015, 9, e0003629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colmant, A.M.G.; O’Brien, C.A.; Newton, N.D.; Watterson, D.; Hardy, J.M.; Coulibaly, F.; Bielefeldt-Ohmann, H.; Warrilow, D.; Huang, B.; Paramitha, D.; et al. Novel monoclonal antibodies against Australian strains of negeviruses and insights into virus structure, replication and host -restriction. J. Gen. Virol. 2020, 101, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piyasena, T.B.H.; Setoh, Y.X.; Hobson-Peters, J.; Newton, N.D.; Bielefeldt-Ohmann, H.; McLean, B.J.; Vet, L.J.; Khromykh, A.A.; Suhrbier, A. Infectious DNAs derived from insect-specific flavivirus genomes enable identification of pre- and post-entry host restrictions in vertebrate cells. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- O’Brien, C.A.; Pegg, C.L.; Nouwens, A.S.; Bielefeldt-Ohmann, H.; Huang, B.; Warrilow, D.; Harrison, J.J.; Haniotis, J.; Schulz, B.L.; Paramitha, D.; et al. A Unique Relative of Rotifer Birnavirus Isolated from Australian Mosquitoes. Viruses 2020, 12, 1056. [Google Scholar] [CrossRef]
- Hall-Mendelin, S.; Ritchie, S.A.; Johansen, C.A.; Zborowski, P.; Cortis, G.; Dandridge, S.; Hall, R.A.; Hurk, A.F.V.D. Exploiting mosquito sugar feeding to detect mosquito-borne pathogens. Proc. Natl. Acad. Sci. USA 2010, 107, 11255–11259. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, S.A.; Cortis, G.; Paton, C.; Townsend, M.; Shroyer, D.; Zborowski, P.; Hall-Mendelin, S.; Hurk, A.F.V.D. A simple non-powered passive trap for the collection of mosquitoes for arbovirus surveillance. J. Med. Ѐntomol. 2013, 50, 185–194. [Google Scholar] [CrossRef]
- Huang, B.; Pyke, A.T.; McMahon, J.L.; Warrilow, D. Complete coding sequence of a case of Chikungunya virus imported into Australia. Genome Announc. 2017, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Auch, A.F.; Qi, J.; Schuster, S.C. MEGAN analysis of metagenomic data. Genome Res. 2007, 17, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Zhou, J.; Jin, Y.; Chen, Y.; Li, J.; Zhang, Q.; Xie, X.; Gan, L.; Liu, Q. Complete genomic and ultrastructural analysis of a Nam Dinh virus isolated from Culex pipiens quinquefasciatus in China. Sci. Rep. 2017, 7, 271. [Google Scholar] [CrossRef] [Green Version]
- Hall, R.A.; Bielefeldt-Ohmann, H.; McLean, B.J.; O’Brien, C.A.; Colmant, A.M.; Piyasena, T.B.; Harrison, J.J.; Newton, N.D.; Barnard, R.T.; Prow, N.A.; et al. Commensal viruses of mosquitoes: Host restriction, transmission, and interaction with arboviral pathogens. Evol. Bioinform. 2016, 12, EBO.S40740. [Google Scholar] [CrossRef]
- Shi, M.; Neville, P.; Nicholson, J.; Eden, J.-S.; Imrie, A.; Holmes, E.C. High-resolution metatranscriptomics reveals the ecological dynamics of mosquito-associated RNA viruses in Western Australia. J. Virol. 2017, 91, e00680-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thuy, N.T.; Huy, T.Q.; Nga, P.T.; Morita, K.; Dunia, I.; Benedetti, L. A new nidovirus (NamDinh virus NDiV): Its ultrastructural characterization in the C6/36 mosquito cell line. Virolology 2013, 444, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.K.; Gui, L. Dissecting virus infectious cycles by cryo-electron microscopy. PLOS Pathog. 2016, 12, e1005625. [Google Scholar] [CrossRef] [PubMed]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.; Baksh, M.F.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G.; et al. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Ehrenstein, M.R.; Notley, C.A. The importance of natural IgM: Scavenger, protector and regulator. Nat. Rev. Immunol. 2010, 10, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Klimovich, V.B. IgM and its receptors: Structural and functional aspects. Biochem. Mosc. 2011, 76, 534–549. [Google Scholar] [CrossRef] [PubMed]
- Bosch, B.J.; Van Der Zee, R.; De Haan, C.A.M.; Rottier, P.J.M. The coronavirus spike protein is a class I virus fusion protein: Structural and functional characterization of the fusion core complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, J.D.; Hay, K.; Rini, J.M.; Yu, M.; Wang, L.; Plummer, F.A.; Corbett, C.R.; Andonov, A. Neutralizing epitopes of the SARS-CoV S-protein cluster independent of repertoire, antigen structure or mAb technology. MAbs 2010, 2, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Follis, K.E.; York, J.; Nunberg, J.H. Furin cleavage of the SARS coronavirus spike glycoprotein enhances cell–cell fusion but does not affect virion entry. Virology 2006, 350, 358–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Hurk, A.F.; Johnson, P.; Hall-Mendelin, S.; Northill, J.A.; Simmons, R.J.; Jansen, C.C.; Frances, S.P.; Smith, G.A.; Ritchie, S.A. Expectoration of Flaviviruses during sugar feeding by mosquitoes (Diptera: Culicidae). J. Med. Ѐntomol. 2007, 44, 845–850. [Google Scholar] [CrossRef]
- Ramírez, A.L.; Hall-Mendelin, S.; Doggett, S.L.; Hewitson, G.R.; McMahon, J.L.; Ritchie, S.A.; van den Hurk, A.F. Mosquito excreta: A sample type with many potential applications for the investigation of Ross River virus and West Nile virus ecology. PLOS Negl. Trop. Dis. 2018, 12, e0006771. [Google Scholar] [CrossRef] [PubMed]
- Higgs, S.; Schneider, B.S.; VanLandingham, D.L.; Klingler, K.A.; Gould, E.A. Nonviremic transmission of West Nile virus. Proc. Natl. Acad. Sci. USA 2005, 102, 8871–8874. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Species | Region | Collection Date | Mosquito Species | Pool Size | Mesonivirus Detections/Total Pools Screened |
|---|---|---|---|---|---|
| Alphamesonivirus 1 (NDiV) | Peel Region, WA | 2014 | Aedes hesperonotius | 2 | 1/2 (incl. isolate DC59899) |
| Aedes camptorhynchus | 5–20 | 28/54 (incl. isolate DC60042) | |||
| Aedes alboannulatus | 2–13 | 3/4 | |||
| Aedes notoscriptus | 1–4 | 2/2 | |||
| Ae. vigilax | 2–7 | 2/2 | |||
| Anopheles annulipes | 1–20 | 4/6 | |||
| Cx. annulirostris | 2–5 | 2/2 | |||
| Culex australicus | 6–20 | 4/5 | |||
| Culex globocoxitus | 16–21 | 4/7 | |||
| Culex quinquefasciatus | 8–20 | 3/3 | |||
| Aedes turneri | 1 | 1/1 | |||
| Cule spp. | 1 | 1/2 | |||
| Aedes spp. | 21 | 1/1 (incl. isolate DC59801) | |||
| Darwin, N | 2018 | Culex quinquefasciatus | 1–37 | 2/10 | |
| Culex pullus | 1–47 | 1/14 | |||
| Ballina, NSW | 2013 | Aedes notoscriptus | 25 | 1/3 (incl. isolate 179853) | |
| Brisbane, QLD | 2005 | Cx. annulirostris | 1/59 | ||
| Pooled Cx. annulirostris and Cq. xanthogaster | 7/100 | ||||
| Alphamesonivirus 4 (CASV) | Cairns, QLD | 2005–2007 | Cx. annulirostris | 1–188 | 8/106 |
| Shoalwater Bay | 2019 | Cx. annulirostris | 1/61 | ||
| Ae. vigilax | 1/82 | ||||
| Ae. procax | 1/42 |
| HOUV | DC59899 | DC59801 | DC60042 | 179853 | NDiV | CavV | DkNV | BBaV | KSaV | KPhV | DKNV | HanaV | SWBTA-V | SWBTA-A | CASV | NSeV | KADV | OFAV | MenoV | Yichang | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HOUV | 99 | 99 | 99 | 99 | 99 | 93 | 93 | 78 | 79 | 80 | 81 | 84 | 78 | 78 | 78 | 67 | 59 | 60 | 56 | 37 | |
| DC59899 | 98 | 100 | 100 | 99 | 99 | 93 | 92 | 78 | 79 | 80 | 81 | 84 | 78 | 78 | 78 | 67 | 59 | 60 | 56 | 37 | |
| DC59801 | 98 | 100 | 100 | 100 | 99 | 93 | 92 | 78 | 79 | 80 | 81 | 84 | 78 | 78 | 78 | 67 | 59 | 60 | 56 | 37 | |
| DC60042 | 98 | 99 | 100 | 100 | 99 | 93 | 92 | 78 | 79 | 80 | 81 | 84 | 78 | 78 | 78 | 67 | 59 | 60 | 56 | 37 | |
| 179853 | 98 | 99 | 99 | 99 | 99 | 93 | 92 | 78 | 79 | 80 | 81 | 84 | 78 | 78 | 78 | 67 | 59 | 60 | 56 | 37 | |
| NDiV | 98 | 99 | 99 | 99 | 99 | 93 | 92 | 78 | 79 | 80 | 81 | 84 | 78 | 78 | 78 | 67 | 59 | 60 | 56 | 37 | |
| CavV | 91 | 91 | 91 | 91 | 91 | 91 | 91 | 77 | 78 | 80 | 80 | 83 | 78 | 78 | 78 | 67 | 59 | 60 | 55 | 37 | |
| DkNV | 90 | 90 | 90 | 90 | 90 | 90 | 89 | 77 | 78 | 80 | 80 | 83 | 78 | 78 | 78 | 67 | 59 | 60 | 56 | 37 | |
| BBaV | 79 | 79 | 79 | 79 | 79 | 79 | 78 | 78 | 91 | 84 | 84 | 75 | 71 | 71 | 72 | 63 | 56 | 57 | 53 | 37 | |
| KSaV | 79 | 79 | 79 | 79 | 79 | 79 | 78 | 78 | 91 | 85 | 84 | 75 | 71 | 71 | 72 | 63 | 56 | 57 | 53 | 37 | |
| KPhV | 81 | 81 | 81 | 81 | 81 | 81 | 80 | 81 | 82 | 82 | 96 | 77 | 74 | 74 | 74 | 65 | 58 | 58 | 54 | 37 | |
| DKNV | 82 | 82 | 81 | 81 | 82 | 82 | 81 | 81 | 82 | 82 | 95 | 77 | 74 | 74 | 74 | 65 | 58 | 59 | 54 | 37 | |
| HanaV | 83 | 83 | 83 | 83 | 83 | 83 | 82 | 83 | 75 | 76 | 78 | 78 | 76 | 76 | 85 | 66 | 58 | 59 | 55 | 37 | |
| SWBTA-V | 77 | 77 | 77 | 77 | 77 | 77 | 77 | 77 | 71 | 72 | 74 | 74 | 76 | 99 | 91 | 66 | 58 | 60 | 55 | 37 | |
| SWBTA-A | 77 | 77 | 77 | 77 | 77 | 77 | 77 | 77 | 71 | 72 | 74 | 74 | 76 | 98 | 90 | 66 | 58 | 60 | 55 | 37 | |
| CASV | 77 | 77 | 77 | 77 | 77 | 77 | 77 | 77 | 71 | 71 | 74 | 74 | 76 | 96 | 95 | 65 | 58 | 59 | 55 | 37 | |
| NSeV | 69 | 69 | 69 | 69 | 69 | 69 | 69 | 69 | 65 | 66 | 67 | 68 | 68 | 67 | 67 | 68 | 59 | 59 | 55 | 37 | |
| KADV | 61 | 61 | 61 | 61 | 61 | 61 | 61 | 62 | 59 | 59 | 61 | 61 | 61 | 61 | 61 | 61 | 62 | 60 | 57 | 37 | |
| OFAV | 58 | 58 | 58 | 58 | 58 | 58 | 58 | 59 | 56 | 56 | 58 | 58 | 59 | 57 | 58 | 58 | 58 | 59 | 56 | 37 | |
| MenoV | 63 | 63 | 63 | 63 | 63 | 63 | 63 | 63 | 60 | 60 | 62 | 62 | 62 | 62 | 62 | 62 | 62 | 61 | 57 | 36 | |
| Yichang | 46 | 46 | 46 | 46 | 46 | 47 | 46 | 47 | 46 | 47 | 47 | 47 | 46 | 46 | 46 | 46 | 46 | 45 | 45 | 45 |
| Name | Made to | Isotype | Cross-Reactivity * | Protein Target # | Microneutralisation § |
|---|---|---|---|---|---|
| C.1G9 | CASV | IgG1 | − | N | <1 |
| C.5D3 | CASV | IgG2a | − | S/S1 | >128 |
| C.5F8 | CASV | IgM | + | M | <1 |
| C.6B9 | CASV | IgM | + | M | <1 |
| C.8G3 | CASV | IgG1 | − | N | <1 |
| C.9D7 | CASV | IgG2a | − | S/S2 | >128 |
| C.10C1 | CASV | IgG2b | − | M | <1 |
| C.10H11 | CASV | IgM/IgG2a | + | M | <1 |
| N.1A10 | NDiV | IgM | − | − | <8 |
| N.1E6 | NDiV | IgM | − | − | 16 |
| N.2A11 | NDiV | IgG2a | + | M | <8 |
| N.2B2 | NDiV | IgM | + | M | <8 |
| N.2D10 | NDiV | IgG2a | + | − | <8 |
| N.2E12 | NDiV | IgG2b | − | S1 | >1024 |
| N.2G8 | NDiV | IgM | − | − | 8 |
| N.3A10 | NDiV | IgG2a | − | − | 16 |
| N.3B2 | NDiV | NT | − | ND | 8 |
| N.3B11 | NDiV | IgG3 | + | M | <8 |
| N.3C10 | NDiV | IgM | − | N | 8 |
| N.3D9 | NDiV | IgG1 | + | − | 128 |
| N.3G4 | NDiV | IgG3 | + | M | <8 |
| N.4C11 | NDiV | IgM | + | ND | <8 |
| N.4D3 | NDiV | IgM | + | M | <8 |
| N.4F9 | NDiV | IgM | − | − | <2 |
| N.4H1 | NDiV | IgG3 | − | N | <2 |
| N.4H7 | NDiV | IgG1 | − | N | <2 |
| N.5E10 | NDiV | IgM | + | ND | <2 |
| N.6A3 | NDiV | IgG1 | − | − | <8 |
| N.6B3 | NDiV | IgG1 | − | − | <8 |
| N.6C3 | NDiV | IgG1 | − | − | <2 |
| N.6C4 | NDiV | IgG2a | − | S | >256 |
| N.6F2 | NDiV | IgG2a | + | − | 16 |
| N.6G5 | NDiV | IgM | − | − | <2 |
| N.7C7 | NDiV | IgG1 | − | − | <8 |
| Location | Collection Year | Samples Positive/Tested for Mesonivirus RNA |
|---|---|---|
| Emerald | 2012/13 | 1/8 |
| Cape York | 2012/14 | 0/8 |
| Mt Isa | 2012 | 0/1 |
| Darwin | 2012 | 2/8 |
| Rockhampton | 2013 | 0/8 |
| Townsville | 2013 | 0/10 |
| St George | 2013 | 0/1 |
| Charleville | 2013 | 0/1 |
| Mareeba | 2013 | 0/1 |
| Badu Island | 2013 | 0/2 |
| Seisia | 2013 | 0/1 |
| Longreach | 2013 | 4/8 |
| 7/57 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Newton, N.D.; Colmant, A.M.G.; O’Brien, C.A.; Ledger, E.; Paramitha, D.; Bielefeldt-Ohmann, H.; Watterson, D.; McLean, B.J.; Hall-Mendelin, S.; Warrilow, D.; et al. Genetic, Morphological and Antigenic Relationships between Mesonivirus Isolates from Australian Mosquitoes and Evidence for Their Horizontal Transmission. Viruses 2020, 12, 1159. https://doi.org/10.3390/v12101159
Newton ND, Colmant AMG, O’Brien CA, Ledger E, Paramitha D, Bielefeldt-Ohmann H, Watterson D, McLean BJ, Hall-Mendelin S, Warrilow D, et al. Genetic, Morphological and Antigenic Relationships between Mesonivirus Isolates from Australian Mosquitoes and Evidence for Their Horizontal Transmission. Viruses. 2020; 12(10):1159. https://doi.org/10.3390/v12101159
Chicago/Turabian StyleNewton, Natalee D., Agathe M. G. Colmant, Caitlin A. O’Brien, Emma Ledger, Devina Paramitha, Helle Bielefeldt-Ohmann, Daniel Watterson, Breeanna J. McLean, Sonja Hall-Mendelin, David Warrilow, and et al. 2020. "Genetic, Morphological and Antigenic Relationships between Mesonivirus Isolates from Australian Mosquitoes and Evidence for Their Horizontal Transmission" Viruses 12, no. 10: 1159. https://doi.org/10.3390/v12101159
APA StyleNewton, N. D., Colmant, A. M. G., O’Brien, C. A., Ledger, E., Paramitha, D., Bielefeldt-Ohmann, H., Watterson, D., McLean, B. J., Hall-Mendelin, S., Warrilow, D., van den Hurk, A. F., Liu, W., Hoare, C., Kizu, J. R., Gauci, P. J., Haniotis, J., Doggett, S. L., Shaban, B., Johansen, C. A., ... Hobson-Peters, J. (2020). Genetic, Morphological and Antigenic Relationships between Mesonivirus Isolates from Australian Mosquitoes and Evidence for Their Horizontal Transmission. Viruses, 12(10), 1159. https://doi.org/10.3390/v12101159