Comparison of the Pathogenicity in Mice of A(H1N1)pdm09 Viruses Isolated between 2009 and 2015 in Japan

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
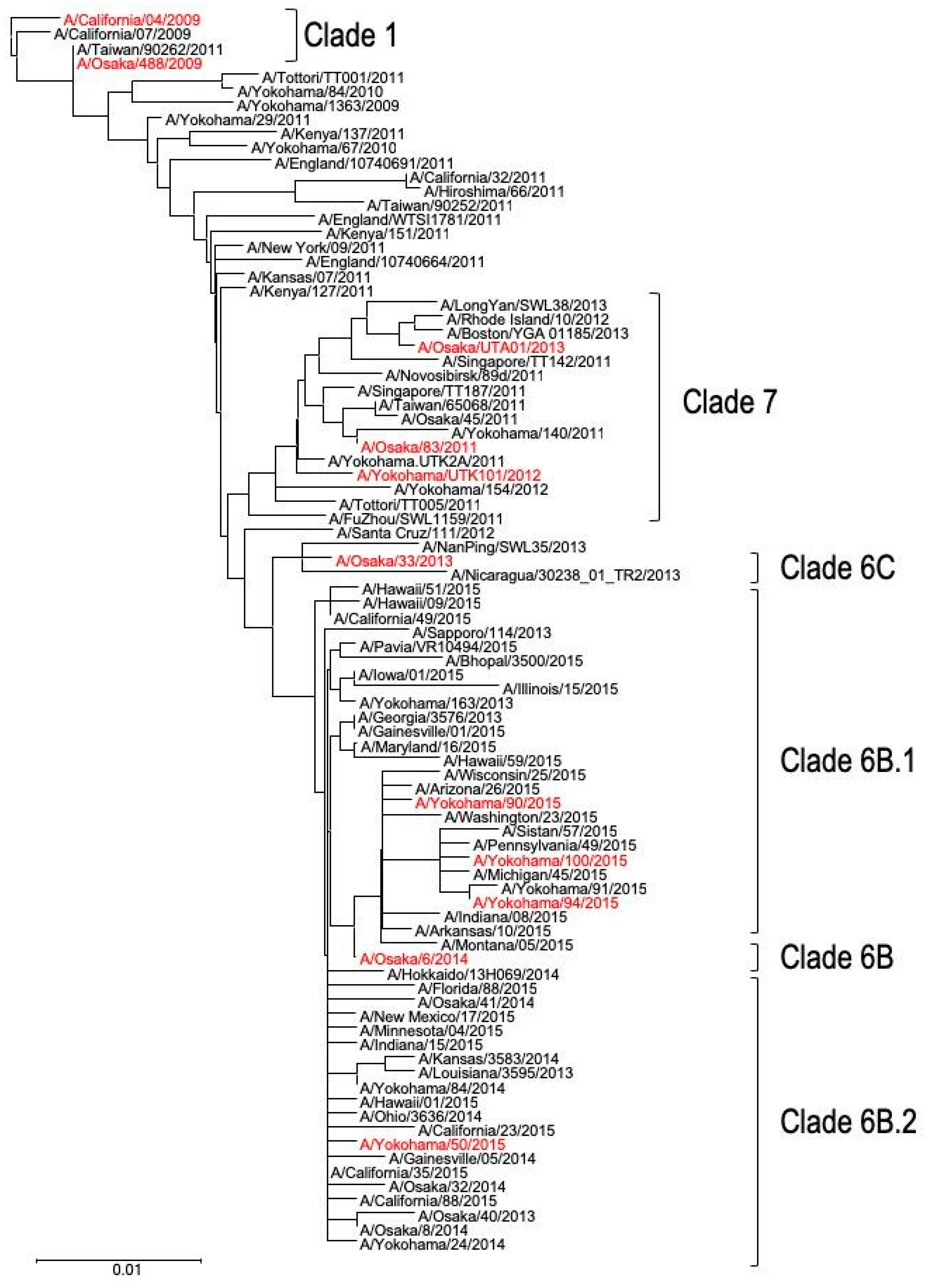
2.2. Phylogenetic Analysis
2.3. Sequencing
2.4. Mouse Experiments
2.5. Statistical Analysis
2.6. Ethics
3. Results
3.1. Genetic Analyses of Japanese A(H1N1)pdm09 Isolates
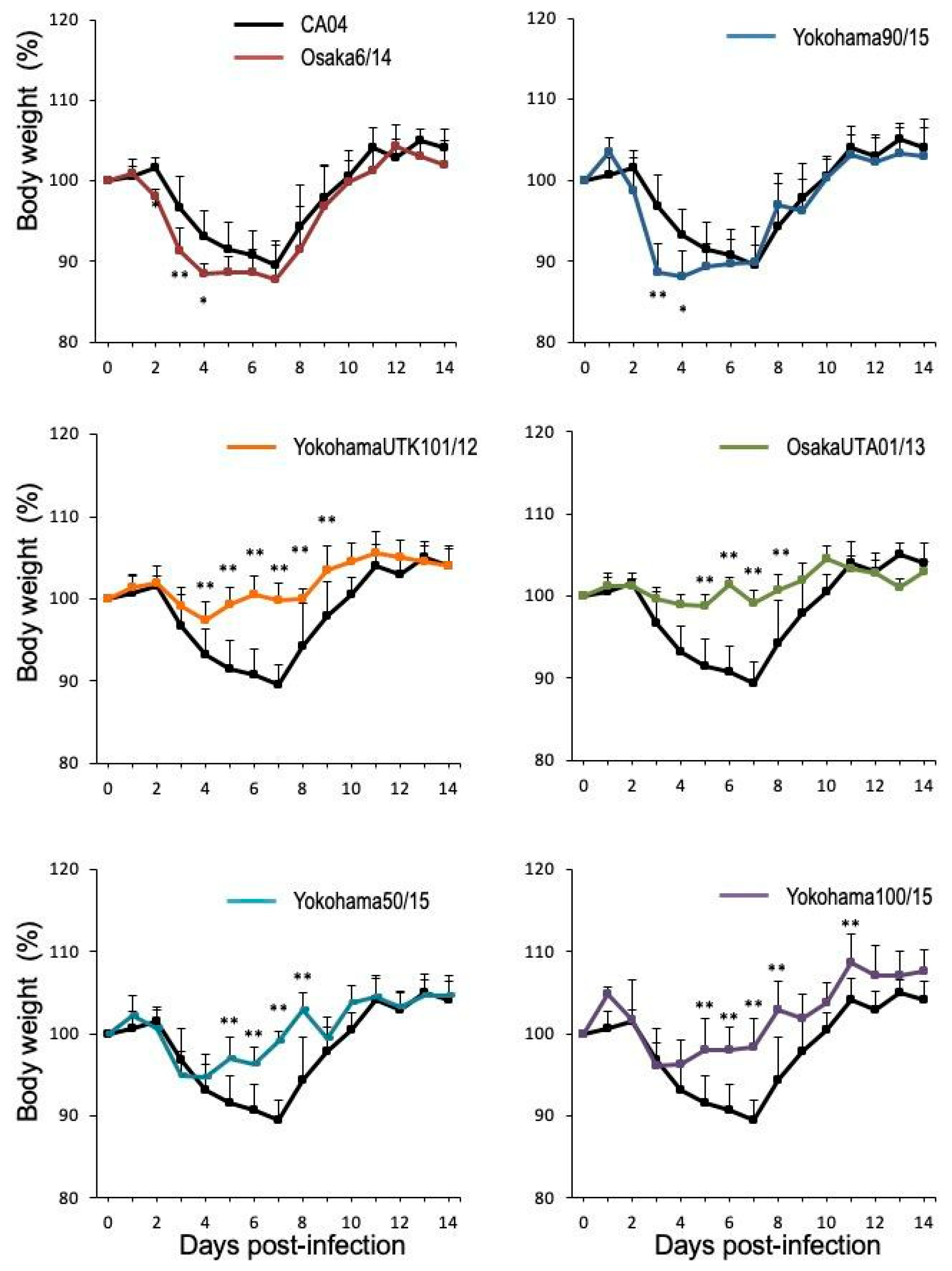
3.2. Pathogenicity of A(H1N1)pdm09 Isolates in Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Garten, R.J.; Davis, C.T.; Russell, C.A.; Shu, B.; Lindstrom, S.; Balish, A.; Sessions, W.M.; Xu, X.; Skepner, E.; Deyde, V.; et al. Antigenic and genetic characteristics of swine-origin 2009 A(H1N1) influenza viruses circulating in humans. Science 2009, 325, 197–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Team Eurosurveillance editorial Pandemic alert level 6: Scientific criteria for an influenza pandemic fulfilled. Eurosurveillance 2009, 14, 1.
- Fraser, C.; Donnelly, C.A.; Cauchemez, S.; Hanage, W.P.; Van Kerkhove, M.D.; Hollingsworth, T.D.; Griffin, J.; Baggaley, R.F.; Jenkins, H.E.; Lyons, E.J.; et al. Pandemic Potential of a Strain of Influenza A (H1N1): Early Findings. Science 2009, 324, 1557–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiris, J.S.M.; Poon, L.L.M.; Guan, Y. Emergence of a novel swine-origin influenza A virus (S-OIV) H1N1 virus in humans. J. Clin. Virol. 2009, 45, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Bautista, E.; Chotpitayasunondh, T.; Gao, Z.; Harper, S.; Shaw, M.; Uyeki, T.M.; Zaki, S.R.; Hayden, F.; Hui, D.; Kettner, J.; et al. Clinical Aspects of Pandemic 2009 Influenza A (H1N1) Virus Infection. N. Engl. J. Med. 2010, 362, 1708–1719. [Google Scholar]
- Jain, S.; Kamimoto, L.; Bramley, A.M.; Schmitz, A.M.; Benoit, S.R.; Louie, J.; Sugerman, D.E.; Druckenmiller, J.K.; Ritger, K.A.; Chugh, R.; et al. Hospitalized Patients with 2009 H1N1 Influenza in the United States, April–June 2009. N. Engl. J. Med. 2009, 361, 1935–1944. [Google Scholar] [CrossRef] [Green Version]
- Shieh, W.-J.; Blau, D.M.; Denison, A.M.; DeLeon-Carnes, M.; Adem, P.; Bhatnagar, J.; Sumner, J.; Liu, L.; Patel, M.; Batten, B.; et al. 2009 Pandemic Influenza A (H1N1): Pathology and Pathogenesis of 100 Fatal Cases in the United States. Am. J. Pathol. 2010, 177, 166–175. [Google Scholar] [CrossRef]
- Bal, A.; Suri, V.; Mishra, B.; Bhalla, A.; Agarwal, R.; Abrol, A.; Kanta Ratho, R.; Joshi, K. Pathology and virology findings in cases of fatal influenza A H1N1 virus infection in 2009–2010. Histopathology 2012, 60, 326–335. [Google Scholar] [CrossRef]
- Nakajima, N.; Hata, S.; Sato, Y.; Tobiume, M.; Katano, H.; Kaneko, K.; Nagata, N.; Kataoka, M.; Ainai, A.; Hasegawa, H.; et al. Short Communication The First Autopsy Case of Pandemic Influenza (A/H1N1pdm) Virus Infection in Japan: Detection of a High Copy Number of the Virus in Type II Alveolar Epithelial Cells by Pathological and Virological Examination. Jpn. J. Infect. Dis. 2010, 63, 67–71. [Google Scholar]
- Virgilia Soto-Abraham, M.; Soriano-Rosas, J.; Diaz-Quiñonez, J.A.; Silva-Pereyra, J.; Vazquez-Hernandez, P.; Torres-López, O.; Roldán, A.; Cruz-Gordillo, A.; Alonso-Viveros, P.; Navarro-Reynoso, F. Pathological Changes Associated with the 2009 H1N1 Virus. N. Engl. J. Med. 2009, 361, 2001–2003. [Google Scholar] [CrossRef]
- Itoh, Y.; Shinya, K.; Kiso, M.; Watanabe, T.; Sakoda, Y.; Hatta, M.; Muramoto, Y.; Tamura, D.; Sakai-Tagawa, Y.; Noda, T.; et al. In vitro and in vivo characterization of new swine-origin H1N1 influenza viruses. Nature 2009, 460, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Maines, T.R.; Jayaraman, A.; Belser, J.A.; Wadford, D.A.; Pappas, C.; Zeng, H.; Gustin, K.M.; Pearce, M.B.; Viswanathan, K.; Shriver, Z.H.; et al. Transmission and Pathogenesis of Swine-Origin 2009 A(H1N1) Influenza Viruses in Ferrets and Mice. Science 2009, 325, 484–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwok, K.; Riley, S.; Perera, R.A.P.M.; Wei, V.W.; Wu, P.; Wei, L.; Chu, D.K.; Barr, I.G.; Peiris, J.M.; Cowling, B.J. Relative incidence and individual-level severity of seasonal influenza A H3N2 compared with 2009 pandemic H1N1. BMC Infect. Dis. 2017, 17, 337. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Torok, M.R.; Bagdure, D.; Cunningham, M.A.; Williams, J.T.B.; Curtis, D.J.; Wilson, K.; Dominguez, S.R. A comparison of H1N1 influenza among pediatric inpatients in the pandemic and post pandemic era. J. Clin. Virol. 2015, 71, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Neumann, G.; Watanabe, T.; Ito, H.; Watanabe, S.; Goto, H.; Gao, P.; Hughes, M.; Perez, D.R.; Donis, R.; Hoffmann, E.; et al. Generation of influenza A viruses entirely from cloned cDNAs. Proc. Natl. Acad. Sci. USA 1999, 96, 9345–9350. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, E.; Stech, J.; Guan, Y.; Webster, R.G.; Perez, D. Universal primer set for the full-length amplification of all Influenza A viruses. Arch. Virol. 2002, 146, 2275–2289. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoint. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.; Spiro, D.; Wentworth, D.; Beck, E.; Fan, J.; Ghedin, E.; Halpin, R.; Bera, J.; Hine, E.; Proudfoot, K.; et al. The early diversification of influenza A/H1N1pdm. PLoS Curr. 2009, 1, RRN1126. [Google Scholar] [CrossRef]
- Goka, E.; Vallely, P.; Mutton, K.; Klapper, P.E. Mutations associated with severity of the pandemic influenza A(H1N1)pdm09 in humans: A systematic review and meta-analysis of epidemiological evidence. Arch. Virol. 2014, 159, 3167–3183. [Google Scholar] [CrossRef]
- Zhao, Z.; Yi, C.; Zhao, L.; Wang, S.; Zhou, L.; Hu, Y.; Zou, W.; Chen, H.; Jin, M. PB2-588I Enhances 2009 H1N1 Pandemic Influenza Virus Virulence by Increasing Viral Replication and Exacerbating PB2 Inhibition of Beta Interferon Expression. J. Virol. 2014, 88, 2260–2267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedde, M.; Wählisch, S.; Wolff, T.; Schweiger, B. Predominance of HA-222D/G Polymorphism in Influenza A(H1N1)pdm09 Viruses Associated with Fatal and Severe Outcomes Recently Circulating in Germany. PLoS ONE 2013, 8, e57059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barde, P.; Sahu, M.; Shukla, M.K.; Bharti, P.; Sharma, R.K.; Sahare, L.; Ukey, M.J.; Singh, N. The high frequency of non-aspartic acid residues at HA222 in influenza A(H1N1) 2009 pandemic viruses is associated with mortality during the upsurge of 2015: A molecular and epidemiological study from central India. Epidemiol. Infect. 2017, 145, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Shinya, K.; Watanabe, S.; Imai, M.; Hatta, M.; Li, C.; Wolter, B.F.; Neumann, G.; Hanson, A.; Ozawa, M.; et al. Avian-type receptor-binding ability can increase influenza virus pathogenicity in macaques. J. Virol. 2011, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooqui, A.; Leon, A.J.; Lei, Y.; Wang, P.; Huang, J.; Tenorio, R.; Dong, W.; Rubino, S.; Lin, J.; Li, G.; et al. Heterogeneous virulence of pandemic 2009 influenza H1N1 virus in mice. Virol. J. 2012, 9, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uraki, R.; Kiso, M.; Shinya, K.; Goto, H.; Takano, R.; Iwatsuki-Horimoto, K.; Takahashi, K.; Daniels, R.S.; Hungnes, O.; Watanabe, T.; et al. Virulence determinants of pandemic A(H1N1)2009 influenza virus in a mouse model. J. Virol. 2013, 87. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Imai, M.; Watanabe, S.; Shinya, K.; Hatta, M.; Li, C.; Neumann, G.; Ozawa, M.; Hanson, A.; Zhong, G.; et al. Characterization in vitro and in vivo of pandemic (H1N1) 2009 influenza viruses isolated from patients. J. Virol. 2012, 86, 9361–9368. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Viruses | Isolation Year | Nucleotide Identity with CA04/09 (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| PB2 | PB1 | PA | HA | NP | NA | M | NS | ||
| Osaka488/09 | 2009 | 99.8 | 99.8 | 99.7 | 99.4 | 99.8 | 99.5 | 99.8 | 100.0 |
| Osaka83/11 | 2011 | 99.1 | 99.4 | 99.1 | 98.8 | 99.4 | 99.1 | 99.2 | 99.0 |
| YokohamaUTK101/12 | 2012 | 99.2 | 99.4 | 99.2 | 98.9 | 99.4 | 99.2 | 99.4 | 99.2 |
| Osaka33/13 | 2013 | 98.6 | 98.9 | 98.6 | 98.4 | 98.7 | 98.9 | 98.5 | 97.8 |
| OsakaUTA01/13 | 2013 | 98.8 | 99.1 | 98.6 | 98.3 | 98.8 | 98.9 | 98.5 | 98.4 |
| Osaka6/14 | 2014 | 98.3 | 98.7 | 98.7 | 98.1 | 98.5 | 98.8 | 99.2 | 97.2 |
| Yokohama50/15 | 2015 | 98.2 | 98.2 | 98.7 | 97.7 | 98.0 | 98.0 | 98.5 | 97.5 |
| Yokohama90/15 | 2015 | 98.1 | 98.2 | 98.1 | 97.8 | 98.3 | 98.0 | 98.8 | 97.2 |
| Yokohama94/15 | 2015 | 98.1 | 98.3 | 98.1 | 97.7 | 98.2 | 98.0 | 98.9 | 97.2 |
| Yokohama100/15 | 2015 | 98.2 | 98.3 | 98.3 | 97.5 | 98.2 | 97.8 | 98.6 | 97.2 |
| Virus | MLD50 | % of Mean Maximum Body Weight Loss (mean ± SD) b |
|---|---|---|
| CA04/09 | >106.5 | 10.5 ± 2.5 |
| Osaka488/09 | >106.5 | 11.4 ± 4.2 |
| Osaka83/11 | >106.5 | 9.1 ± 3.1 |
| YokohamaUTK101/12 | >106.5 | 2.6 ± 2.3**c |
| Osaka33/13 | >106.5 | 7.6 ± 2.9 |
| Osaka UTA01/13 | >106.5 | 1.2 ± 1.3** |
| Osaka 6/14 | 105.8 | 12.3 ± 4.8 |
| Yokohama50/15 | >106.5 | 5.4 ± 2.9* |
| Yokohama90/15 | 105.8 | 11.8 ± 3.1 |
| Yokohama94/15 | >106.5 | 8.6 ± 5.0 |
| Yokohama100/15 | >106.5 | 3.9 ± 2.8* |
| Virus | Virus Titers (mean log10 PFU/g ±SD)in the Lung | |
|---|---|---|
| Day 3 | Day 6 | |
| CA04/09 | 7.2 ± 0.1 | 5.9 ± 0.7 |
| Osaka488/09 | 7.2 ± 0.2 | 6.4 ± 0.1 |
| Osaka83/11 | 7.1 ± 0.1 | 6.2 ± 0.2 |
| YokohamaUT-K101/12 | 6.3 ± 0.2 ** b | 5.3 ± 0.1 |
| Osaka33/13 | 7.2 ± 0.2 | 6.1 ± 0.2 |
| OsakaUT-A01/13 | 6.9 ± 0.1 * | 5.9 ± 0.3 |
| Osaka6/14 | 7.8 ± 0.2 ** | 6.2 ± 0.4 |
| Yokohama50/15 | 7.4 ± 0.1 | 6.0 ± 0.8 |
| Yokohama90/15 | 7.8 ± 0.2 ** | 6.0 ± 0.2 |
| Yokohama94/15 | 7.7 ± 0.1 ** | 6.2 ± 0.2 |
| Yokohama100/15 | 7.1 ± 0.1 | 5.9 ± 0.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mitake, H.; Yasuhara, A.; Lopes, T.J.S.; Tagawa-Sakai, Y.; Shimizu, K.; Ozawa, H.; Kawakami, C.; Morikawa, S.; Sugaya, N.; Watanabe, T.; et al. Comparison of the Pathogenicity in Mice of A(H1N1)pdm09 Viruses Isolated between 2009 and 2015 in Japan. Viruses 2020, 12, 155. https://doi.org/10.3390/v12020155
Mitake H, Yasuhara A, Lopes TJS, Tagawa-Sakai Y, Shimizu K, Ozawa H, Kawakami C, Morikawa S, Sugaya N, Watanabe T, et al. Comparison of the Pathogenicity in Mice of A(H1N1)pdm09 Viruses Isolated between 2009 and 2015 in Japan. Viruses. 2020; 12(2):155. https://doi.org/10.3390/v12020155
Chicago/Turabian StyleMitake, Hiromichi, Atsuhiro Yasuhara, Tiago J. S. Lopes, Yuko Tagawa-Sakai, Kohei Shimizu, Hiroki Ozawa, Chiharu Kawakami, Saeko Morikawa, Norio Sugaya, Tokiko Watanabe, and et al. 2020. "Comparison of the Pathogenicity in Mice of A(H1N1)pdm09 Viruses Isolated between 2009 and 2015 in Japan" Viruses 12, no. 2: 155. https://doi.org/10.3390/v12020155
APA StyleMitake, H., Yasuhara, A., Lopes, T. J. S., Tagawa-Sakai, Y., Shimizu, K., Ozawa, H., Kawakami, C., Morikawa, S., Sugaya, N., Watanabe, T., & Kawaoka, Y. (2020). Comparison of the Pathogenicity in Mice of A(H1N1)pdm09 Viruses Isolated between 2009 and 2015 in Japan. Viruses, 12(2), 155. https://doi.org/10.3390/v12020155