MicroRNA-7 Inhibits Rotavirus Replication by Targeting Viral NSP5 In Vivo and In Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cells and Virus
2.3. MA104 Cells Transfection, Viral RNA and Protein Detection
2.4. Prediction of Binding Site to miR-7 by miRanda Software
2.5. Luciferase Assay
2.6. Immunofluorescence Staining
2.7. Virus Titer Detection
2.8. Viral Load Detection and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Assay
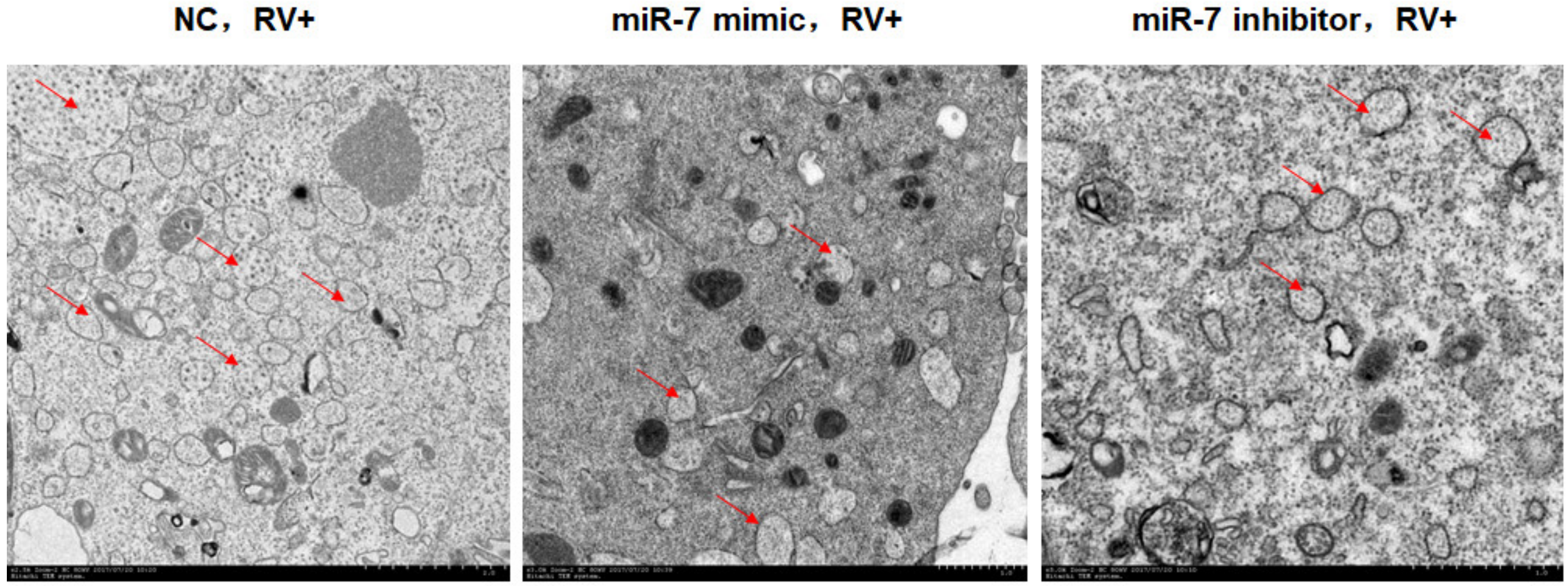
2.9. Observation of Viroplasm by Transmission Electron Microscopy (TEM)
2.10. Animal Inoculation and Virus Challenge
2.11. Histopathological and Jejunum Section Immunofluorescence
2.12. Statistical Analysis
3. Results
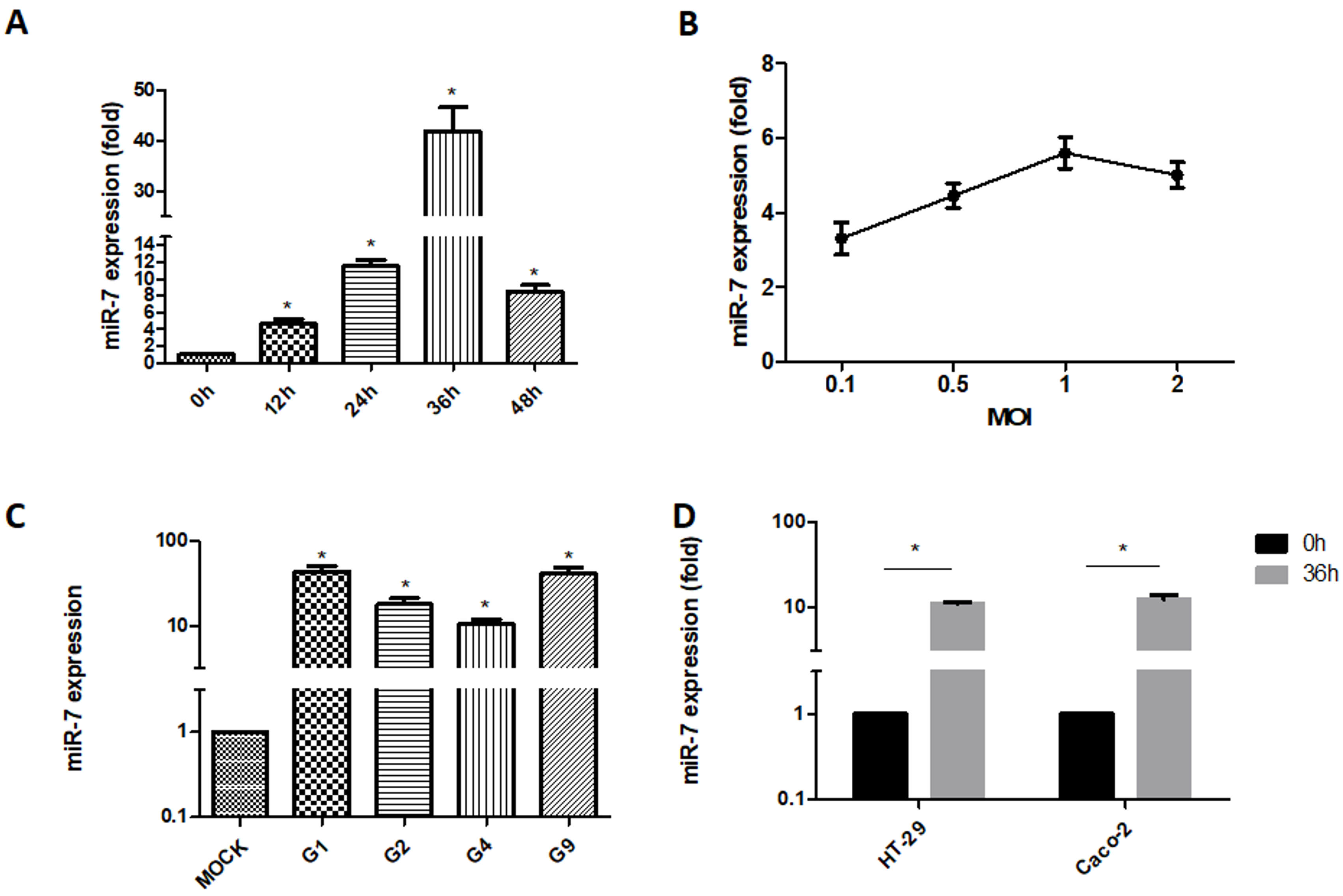
3.1. Detection of MIR-7 Expression during Rotavirus Replication
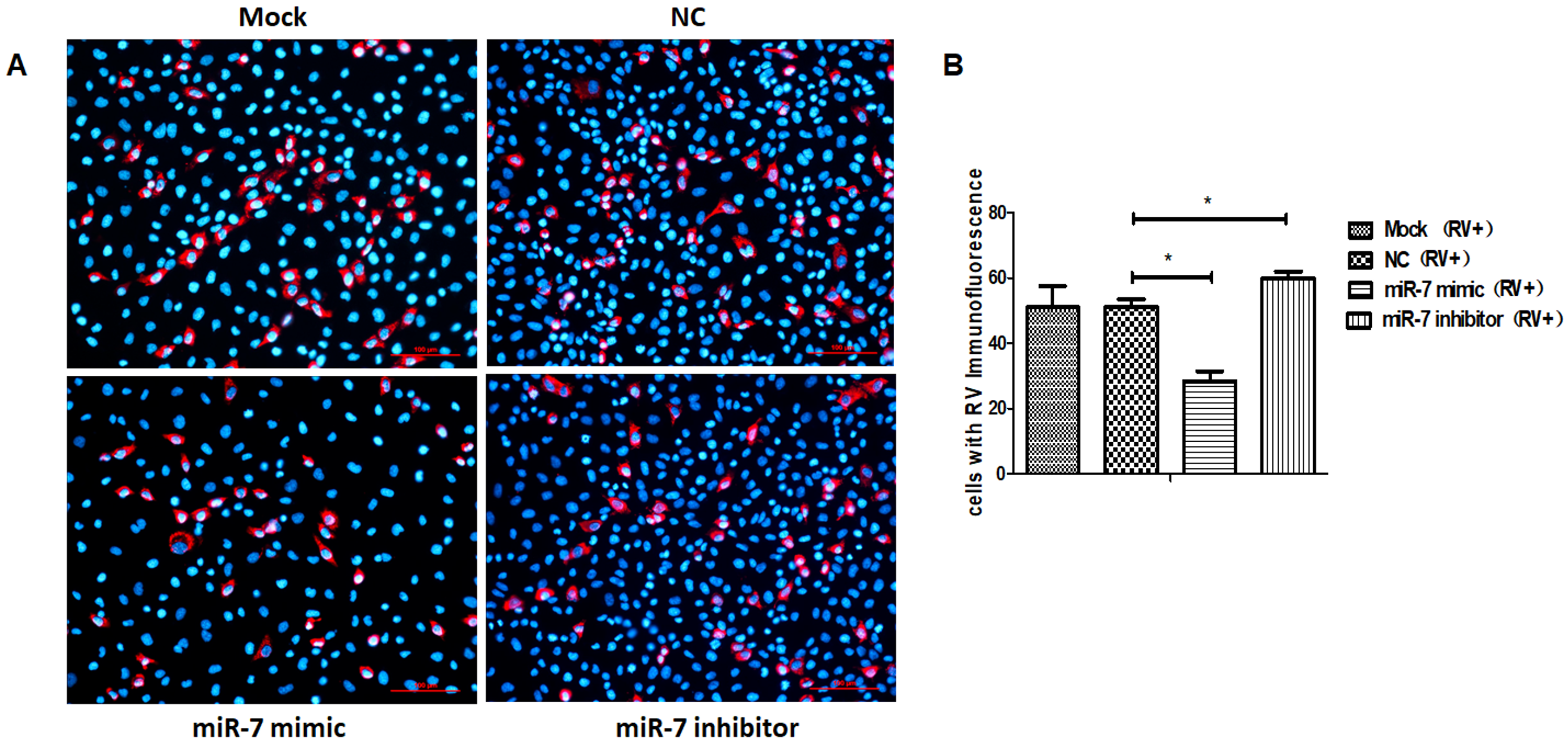
3.2. The Role of MIR-7 in Rotavirus Replication
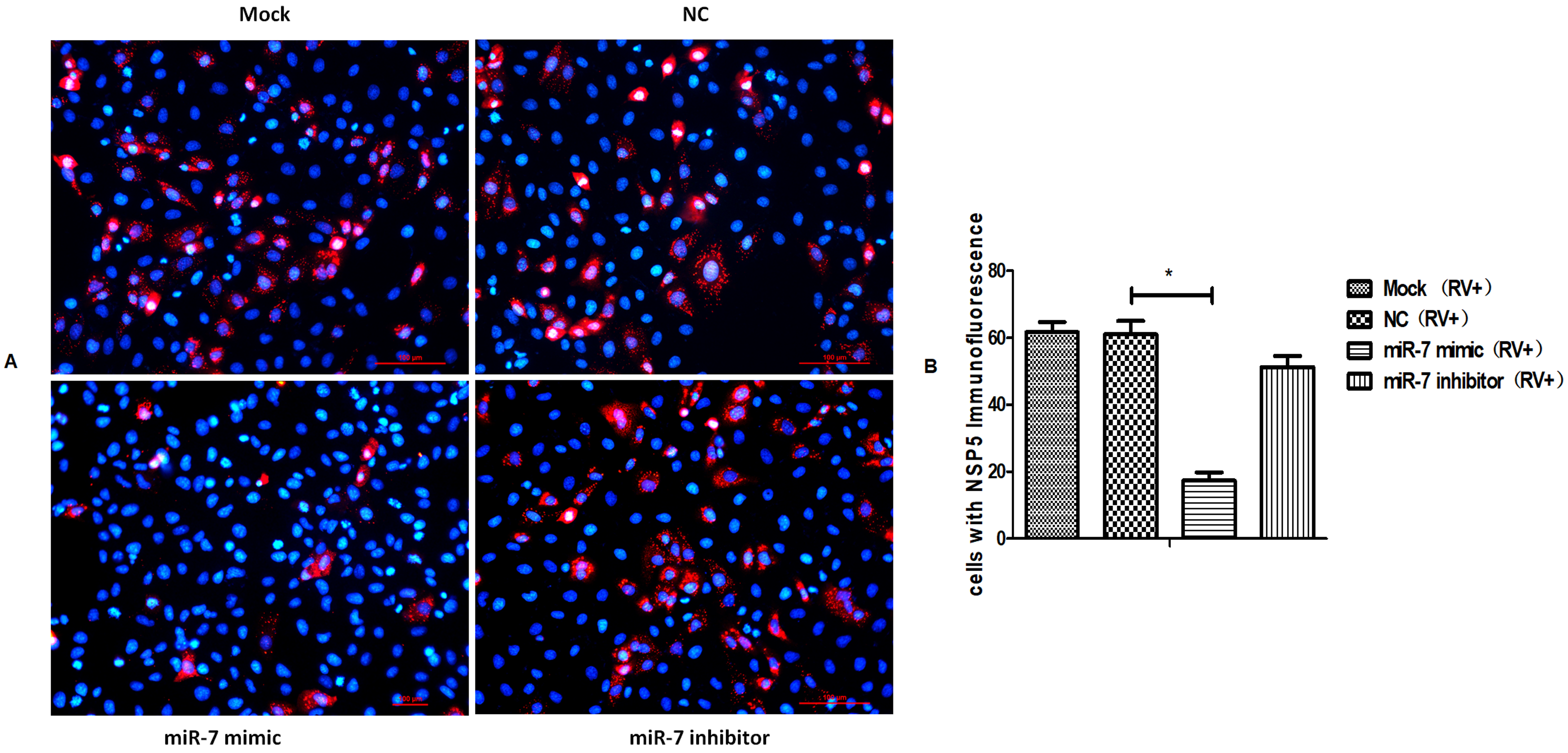
3.3. MIR-7 Regulates Rotavirus Gene Replication by Targeting the NSP5 Gene
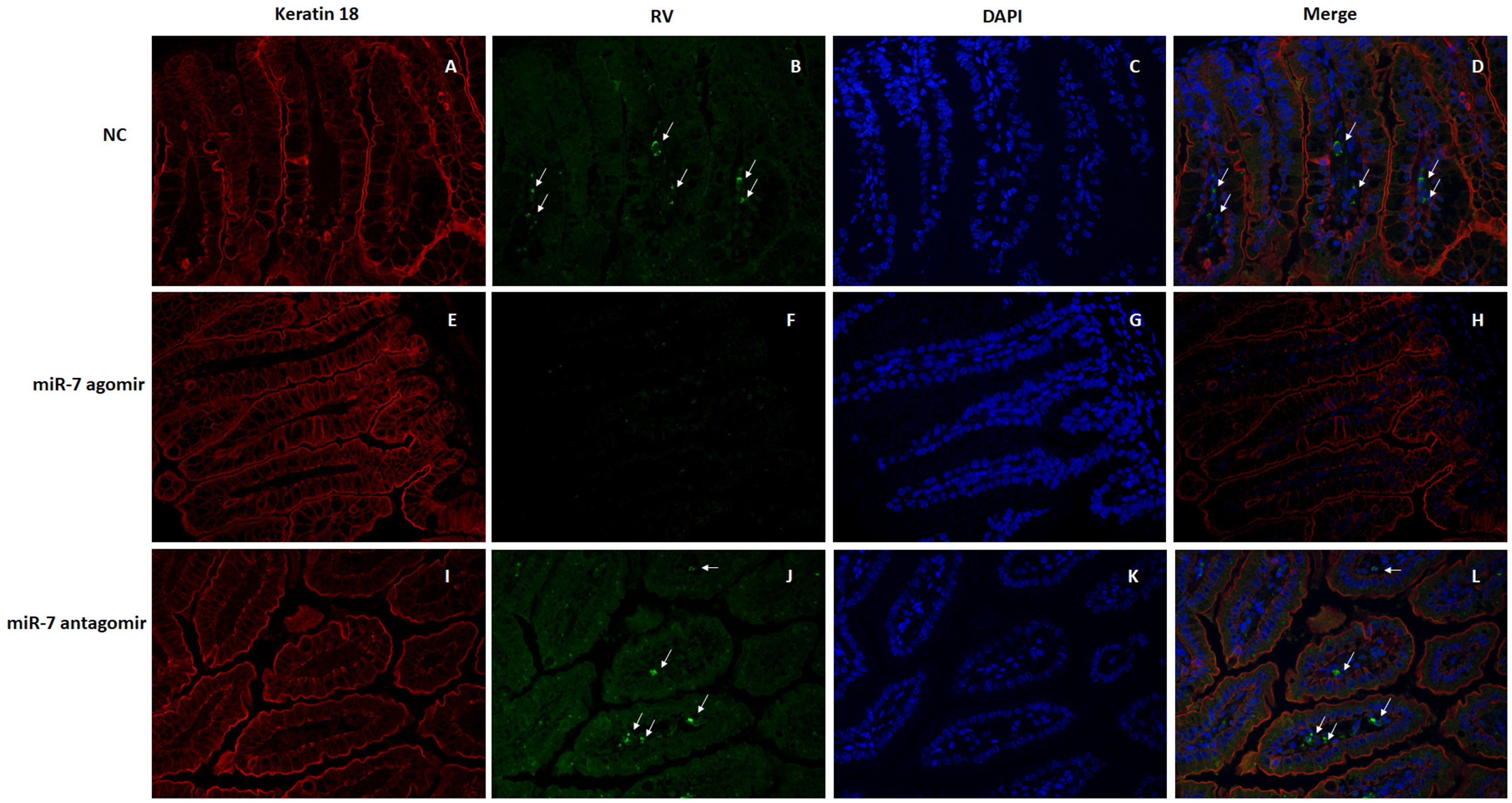
3.4. The Role of MIR-7 in a Rotavirus Diarrhea Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Parashar, U.D.; For the World Health Organization–Coordinated Global Rotavirus Surveillance Network; Agocs, M.; Serhan, F.; de Oliveira, L.; Mwenda, J.M.; Mihigo, R.; et al. Global, Regional, and National Estimates of Rotavirus Mortality in Children <5 Years of Age, 2000–2013. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2016, 62, S96–S105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthijnssens, J.; Bilcke, J.; Ciarlet, M.; Martella, V.; Banyai, K.; Rahman, M.; Zeller, M.; Beutels, P.; Van Damme, P.; Van Ranst, M. Rotavirus disease and vaccination: Impact on genotype diversity. Future Microbiol. 2009, 4, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Estes, M.K.; Howley, P.M. Fields Virology, 6th ed.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 1347–1401. [Google Scholar]
- Long, C.P.; McDonald, S.M. Rotavirus genome replication: Some assembly required. PLoS Pathog. 2017, 13, e1006242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichwald, C.; Vascotto, F.; Fabbretti, E.; Burrone, O.R. Rotavirus NSP5: Mapping phosphorylation sites and kinase activation and viroplasm localization domains. J. Virol. 2002, 76, 3461–3470. [Google Scholar] [CrossRef] [Green Version]
- Afrikanova, I.; Fabbretti, E.; Miozzo, M.C.; Burrone, O.R. Rotavirus NSP5 phosphorylation is up-regulated by interaction with NSP2. J. Gen. Virol. 1998, 79, 2679–2686. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.; Ouldali, M.; Menetrey, J.; Poncet, D. Structural organisation of the rotavirus nonstructural protein NSP5. J. Mol. Biol. 2011, 413, 209–221. [Google Scholar] [CrossRef]
- Sen, A.; Sen, N.; Mackow, E.R. The formation of viroplasm-like structures by the rotavirus NSP5 protein is calcium regulated and directed by a C-terminal helical domain. J. Virol. 2007, 81, 11758–11767. [Google Scholar] [CrossRef] [Green Version]
- Sotelo, P.H.; Schumann, M.; Krause, E.; Chnaiderman, J. Analysis of rotavirus non-structural protein NSP5 by mass spectrometry reveals a complex phosphorylation pattern. Virus Res. 2010, 149, 104–108. [Google Scholar] [CrossRef]
- Sen, A.; Agresti, D.; Mackow, E.R. Hyperphosphorylation of the rotavirus NSP5 protein is independent of serine 67, [corrected] NSP2, or [corrected] the intrinsic insolubility of NSP5 is regulated by cellular phosphatases. J. Virol. 2006, 80, 1807–1816. [Google Scholar] [CrossRef] [Green Version]
- Criglar, J.M.; Hu, L.; Crawford, S.E.; Hyser, J.M.; Broughman, J.R.; Prasad, B.V.; Estes, M.K. A novel form of rotavirus NSP2 and phosphorylation-dependent NSP2-NSP5 interactions are associated with viroplasm assembly. J. Virol. 2014, 88, 786–798. [Google Scholar] [CrossRef] [Green Version]
- Contin, R.; Arnoldi, F.; Campagna, M.; Burrone, O.R. Rotavirus NSP5 orchestrates recruitment of viroplasmic proteins. J. Gen. Virol. 2010, 91, 1782–1793. [Google Scholar] [CrossRef] [PubMed]
- Eichwald, C.; Rodriguez, J.F.; Burrone, O.R. Characterization of rotavirus NSP2/NSP5 interactions and the dynamics of viroplasm formation. J. Gen. Virol. 2004, 85, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Lopez, T.; Rojas, M.; Ayala-Breton, C.; Lopez, S.; Arias, C.F. Reduced expression of the rotavirus NSP5 gene has a pleiotropic effect on virus replication. J. Gen. Virol. 2005, 86, 1609–1617. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Charpilienne, A.; Parent, A.; Boussac, A.; D’Autreaux, B.; Poupon, J.; Poncet, D. The rotavirus nonstructural protein NSP5 coordinates a [2Fe-2S] iron-sulfur cluster that modulates interaction to RNA. FASEB J. 2013, 27, 1074–1083. [Google Scholar] [CrossRef]
- Campagna, M.; Eichwald, C.; Vascotto, F.; Burrone, O.R. RNA interference of rotavirus segment 11 mRNA reveals the essential role of NSP5 in the virus replicative cycle. J. Gen. Virol. 2005, 86, 1481–1487. [Google Scholar] [CrossRef]
- Chnaiderman, J.; Barro, M.; Spencer, E. NSP5 phosphorylation regulates the fate of viral mRNA in rotavirus infected cells. Arch. Virol. 2002, 147, 1899–1911. [Google Scholar] [CrossRef]
- Schirle, N.T.; Sheu-Gruttadauria, J.; MacRae, I.J. Structural basis for microRNA targeting. Science 2014, 346, 608–613. [Google Scholar] [CrossRef]
- Lim, L.P.; Glasner, M.E.; Yekta, S.; Burge, C.B.; Bartel, D.P. Vertebrate microRNA genes. Science 2003, 299, 1540. [Google Scholar] [CrossRef] [Green Version]
- Seeley, J.J.; Baker, R.G.; Mohamed, G.; Bruns, T.; Hayden, M.S.; Deshmukh, S.D.; Freedberg, D.E.; Ghosh, S. Induction of innate immune memory via microRNA targeting of chromatin remodelling factors. Nature 2018, 559, 114–119. [Google Scholar] [CrossRef]
- Zhang, X.; Zuo, X.; Yang, B.; Li, Z.; Xue, Y.; Zhou, Y.; Huang, J.; Zhao, X.; Zhou, J.; Yan, Y.; et al. MicroRNA directly enhances mitochondrial translation during muscle differentiation. Cell 2014, 158, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Rajewsky, K. MicroRNA control in the immune system: Basic principles. Cell 2009, 136, 26–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Rooij, E.; Sutherland, L.B.; Qi, X.; Richardson, J.A.; Hill, J.; Olson, E.N. Control of stress-dependent cardiac growth and gene expression by a microRNA. Science 2007, 316, 575–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Careem, M.F.; Hunter, B.D.; Sarson, A.J.; Parvizi, P.; Haghighi, H.R.; Read, L.; Heidari, M.; Sharif, S. Host responses are induced in feathers of chickens infected with Marek’s disease virus. Virology 2008, 370, 323–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, L.; Yue, W.; Du, S.; Xin, S.; Zhang, J.; Liu, L.; Li, G.; Lu, J. An update: Epstein-Barr virus and immune evasion via microRNA regulation. Virol. Sin. 2017, 32, 175–187. [Google Scholar] [CrossRef]
- Huang, T.; Zhang, X. Functional analysis of a crustacean microRNA in host-virus interactions. J. Virol. 2012, 86, 12997–13004. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.; Wang, K.; Chen, H.; Liu, X.; Lv, J.; Hou, S.; Zhang, Y.; Sun, Y. Host microRNA miR-1307 suppresses foot-and-mouth disease virus replication by promoting VP3 degradation and enhancing innate immune response. Virology 2019, 535, 162–170. [Google Scholar] [CrossRef]
- Cazalla, D.; Yario, T.; Steitz, J.A. Down-regulation of a host microRNA by a Herpesvirus saimiri noncoding RNA. Science 2010, 328, 1563–1566. [Google Scholar] [CrossRef] [Green Version]
- Aparicio, R.; Simoes Da Silva, C.J.; Busturia, A. MicroRNA miR-7 contributes to the control of Drosophila wing growth. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2015, 244, 21–30. [Google Scholar] [CrossRef]
- Correa-Medina, M.; Bravo-Egana, V.; Rosero, S.; Ricordi, C.; Edlund, H.; Diez, J.; Pastori, R.L. MicroRNA miR-7 is preferentially expressed in endocrine cells of the developing and adult human pancreas. Gene Expr. Patterns GEP 2009, 9, 193–199. [Google Scholar] [CrossRef]
- Lopez-Beas, J.; Capilla-Gonzalez, V.; Aguilera, Y.; Mellado, N.; Lachaud, C.C.; Martin, F.; Smani, T.; Soria, B.; Hmadcha, A. miR-7 Modulates hESC Differentiation into Insulin-Producing Beta-like Cells and Contributes to Cell Maturation. Mol. Ther. Nucleic Acids 2018, 12, 463–477. [Google Scholar] [CrossRef]
- Erratum: CircRNA CDR1as/miR-7 signals promote tumor growth of osteosarcoma with a potential therapeutic and diagnostic value [Corrigendum]. Cancer Manag. Res. 2018, 10, 5867. [CrossRef] [PubMed] [Green Version]
- Akalay, I.; Tan, T.Z.; Kumar, P.; Janji, B.; Mami-Chouaib, F.; Charpy, C.; Vielh, P.; Larsen, A.K.; Thiery, J.P.; Sabbah, M.; et al. Targeting WNT1-inducible signaling pathway protein 2 alters human breast cancer cell susceptibility to specific lysis through regulation of KLF-4 and miR-7 expression. Oncogene 2015, 34, 2261–2271. [Google Scholar] [CrossRef] [PubMed]
- Bhere, D.; Tamura, K.; Wakimoto, H.; Choi, S.H.; Purow, B.; Debatisse, J.; Shah, K. microRNA-7 upregulates death receptor 5 and primes resistant brain tumors to caspase-mediated apoptosis. Neuro-Oncology 2018, 20, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, S.; Wang, J.; Wang, J.; Wu, J.; Song, J.; Zhang, C.Y.; Zhang, C.; Wang, C.; Wang, J.J. Increased serum miR-7 is a promising biomarker for type 2 diabetes mellitus and its microvascular complications. Diabetes Res. Clin. Pract. 2017, 130, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, D.; Zhang, S.; Wei, X.; Song, J.; Zhang, Y.; Jin, M.; Shen, Z.; Wang, X.; Feng, Z.; et al. Host-virus interaction: The antiviral defense function of small interfering RNAs can be enhanced by host microRNA-7 in vitro. Sci. Rep. 2015, 5, 9722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, M.C.; Van Rij, R.P.; Andino, R. RNA silencing in viral infections: Insights from poliovirus. Virus Res. 2004, 102, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, W.; Xu, Z.; Pan, J.; Zhao, Z.; Ren, Q. Eriocheir sinensis microRNA-7 targets crab Myd88 to enhance white spot syndrome virus replication. Fish. Shellfish Immunol. 2018, 79, 274–283. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, J.; Geng, P.; Kui, X.; Xie, Y.; Zhang, L.; Liu, Y.; Yin, N.; Zhang, G.; Yi, S.; et al. MicroRNA profile analysis of host cells before and after wild human rotavirus infection. J. Med. Virol. 2016, 88, 1497–1510. [Google Scholar] [CrossRef]
- Li, S.; Zhang, G.; Wu, J.; Yin, N.; Yi, S.; Mi, K.; Cao, Y.; Li, Y.; Sun, M.; Li, H. Isolation human rotavirus strains and adaptive culture. China Biotechnol. 2013, 33, 9–14. [Google Scholar]
- Enright, A.J.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D.S. MicroRNA targets in Drosophila. Genome Biol. 2003, 5, R1. [Google Scholar] [CrossRef] [Green Version]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human MicroRNA targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Geng, P.; Liu, Y.; Wu, J.; Qiao, H.; Xie, Y.; Yin, N.; Chen, L.; Lin, X.; Liu, Y.; et al. Rotavirus-encoded virus-like small RNA triggers autophagy by targeting IGF1R via the PI3K/Akt/mTOR pathway. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Qiao, H.; Yin, N.; Chen, L.; Xie, Y.; Wu, J.; Du, J.; Lin, X.; Wang, Y.; Liu, Y.; et al. Immune and cytokine/chemokine responses of PBMCs in rotavirus-infected rhesus infants and their significance in viral pathogenesis. J. Med. Virol. 2019, 91, 1448–1469. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.T.; de Souza, C.O.; Asensi, M.; Buesa, J. Homotypic protection against rotavirus-induced diarrhea in infant mice breast-fed by dams immunized with the recombinant VP8* subunit of the VP4 capsid protein. Viral Immunol. 2000, 13, 187–200. [Google Scholar] [CrossRef]
- Offit, P.A.; Clark, H.F.; Kornstein, M.J.; Plotkin, S.A. A murine model for oral infection with a primate rotavirus (simian SA11). J. Virol. 1984, 51, 233–236. [Google Scholar] [CrossRef] [Green Version]
- Li, J.T.; Wei, J.; Guo, H.X.; Han, J.B.; Ye, N.; He, H.Y.; Yu, T.T.; Wu, Y.Z. Development of a human rotavirus induced diarrhea model in Chinese mini-pigs. World J. Gastroenterol. 2016, 22, 7135–7145. [Google Scholar] [CrossRef]
- Boshuizen, J.A.; Reimerink, J.H.; Korteland-van Male, A.M.; van Ham, V.J.; Koopmans, M.P.; Buller, H.A.; Dekker, J.; Einerhand, A.W. Changes in small intestinal homeostasis, morphology, and gene expression during rotavirus infection of infant mice. J. Virol. 2003, 77, 13005–13016. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhu, F.; Chen, P. miR-7 and miR-218 epigenetically control tumor suppressor genes RASSF1A and Claudin-6 by targeting HoxB3 in breast cancer. Biochem. Biophys. Res. Commun. 2012, 424, 28–33. [Google Scholar] [CrossRef]
- Chakrabarti, M.; Ai, W.; Banik, N.L.; Ray, S.K. Overexpression of miR-7-1 increases efficacy of green tea polyphenols for induction of apoptosis in human malignant neuroblastoma SH-SY5Y and SK-N-DZ cells. Neurochem. Res. 2013, 38, 420–432. [Google Scholar] [CrossRef]
- Hansen, T.B.; Kjems, J.; Damgaard, C.K. Circular RNA and miR-7 in cancer. Cancer Res. 2013, 73, 5609–5612. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Chen, M.; Zhou, J.; Mo, M.S.; Zhu, L.H.; Liu, Y.P.; Gui, Q.J.; Zhang, L.; Li, G.Q. miR-7 inhibits the invasion and metastasis of gastric cancer cells by suppressing epidermal growth factor receptor expression. Oncol. Rep. 2014, 31, 1715–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Cai, K.; Wang, J.; Wang, X.; Cheng, K.; Shi, F.; Jiang, L.; Zhang, Y.; Dou, J. MiR-7, inhibited indirectly by lincRNA HOTAIR, directly inhibits SETDB1 and reverses the EMT of breast cancer stem cells by downregulating the STAT3 pathway. Stem. Cells 2014, 32, 2858–2868. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.Y.; Kong, W.; Jiang, J.; Xu, H.; Zhao, W. miR-7-5p inhibits cell migration and invasion in glioblastoma through targeting SATB1. Oncol. Lett. 2019, 17, 1819–1825. [Google Scholar] [CrossRef] [Green Version]
- Tazawa, H.; Yano, S.; Yoshida, R.; Yamasaki, Y.; Sasaki, T.; Hashimoto, Y.; Kuroda, S.; Ouchi, M.; Onishi, T.; Uno, F.; et al. Genetically engineered oncolytic adenovirus induces autophagic cell death through an E2F1-microRNA-7-epidermal growth factor receptor axis. Int. J. Cancer 2012, 131, 2939–2950. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Chien, P.H.; Chen, W.S.; Chien, Y.F.; Hsu, Y.Y.; Wang, L.Y.; Chen, J.Y.; Lin, C.W.; Huang, T.C.; Yu, Y.L.; et al. Hepatitis B Virus-Encoded X Protein Downregulates EGFR Expression via Inducing MicroRNA-7 in Hepatocellular Carcinoma Cells. Evid. Based Complement. Altern. Med. eCAM 2013, 2013, 682380. [Google Scholar] [CrossRef] [Green Version]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Chen, L.; Du, J.; Hu, X.; Xie, Y.; Wu, J.; Lin, X.; Yin, N.; Sun, M.; Li, H. MicroRNA-7 Inhibits Rotavirus Replication by Targeting Viral NSP5 In Vivo and In Vitro. Viruses 2020, 12, 209. https://doi.org/10.3390/v12020209
Zhou Y, Chen L, Du J, Hu X, Xie Y, Wu J, Lin X, Yin N, Sun M, Li H. MicroRNA-7 Inhibits Rotavirus Replication by Targeting Viral NSP5 In Vivo and In Vitro. Viruses. 2020; 12(2):209. https://doi.org/10.3390/v12020209
Chicago/Turabian StyleZhou, Yan, Linlin Chen, Jing Du, Xiaoqing Hu, Yuping Xie, Jinyuan Wu, Xiaochen Lin, Na Yin, Maosheng Sun, and Hongjun Li. 2020. "MicroRNA-7 Inhibits Rotavirus Replication by Targeting Viral NSP5 In Vivo and In Vitro" Viruses 12, no. 2: 209. https://doi.org/10.3390/v12020209
APA StyleZhou, Y., Chen, L., Du, J., Hu, X., Xie, Y., Wu, J., Lin, X., Yin, N., Sun, M., & Li, H. (2020). MicroRNA-7 Inhibits Rotavirus Replication by Targeting Viral NSP5 In Vivo and In Vitro. Viruses, 12(2), 209. https://doi.org/10.3390/v12020209