New Viral Sequences Identified in the Flavescence Dorée Phytoplasma Vector Scaphoideus titanus


 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Collection
2.2. DNA Extraction, RNA Extraction, Library Preparation and Sequencing
2.3. Bioinformatic Analysis
2.4. Phylogenetic Trees
2.5. PCR and RT-PCR
3. Results
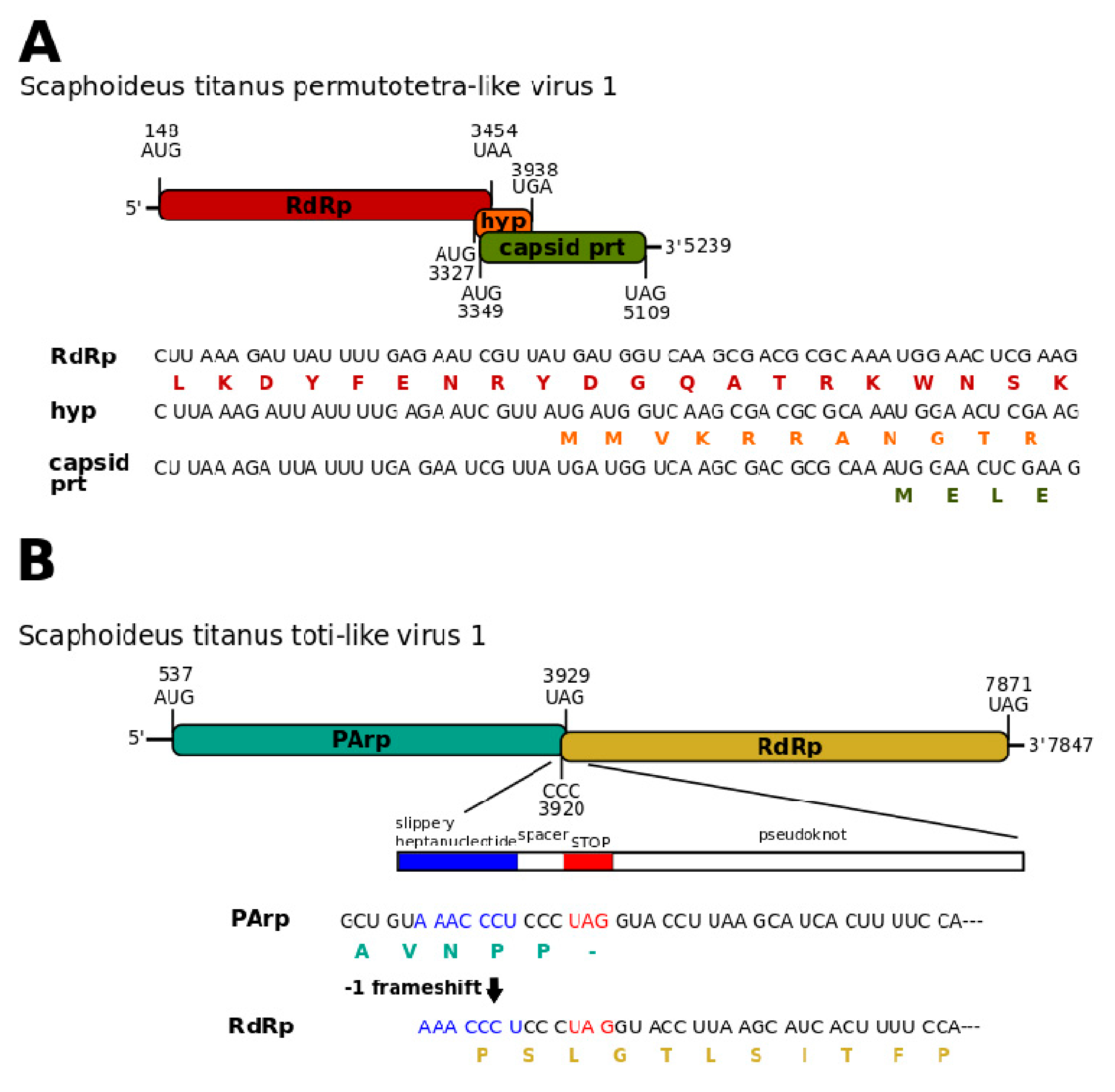
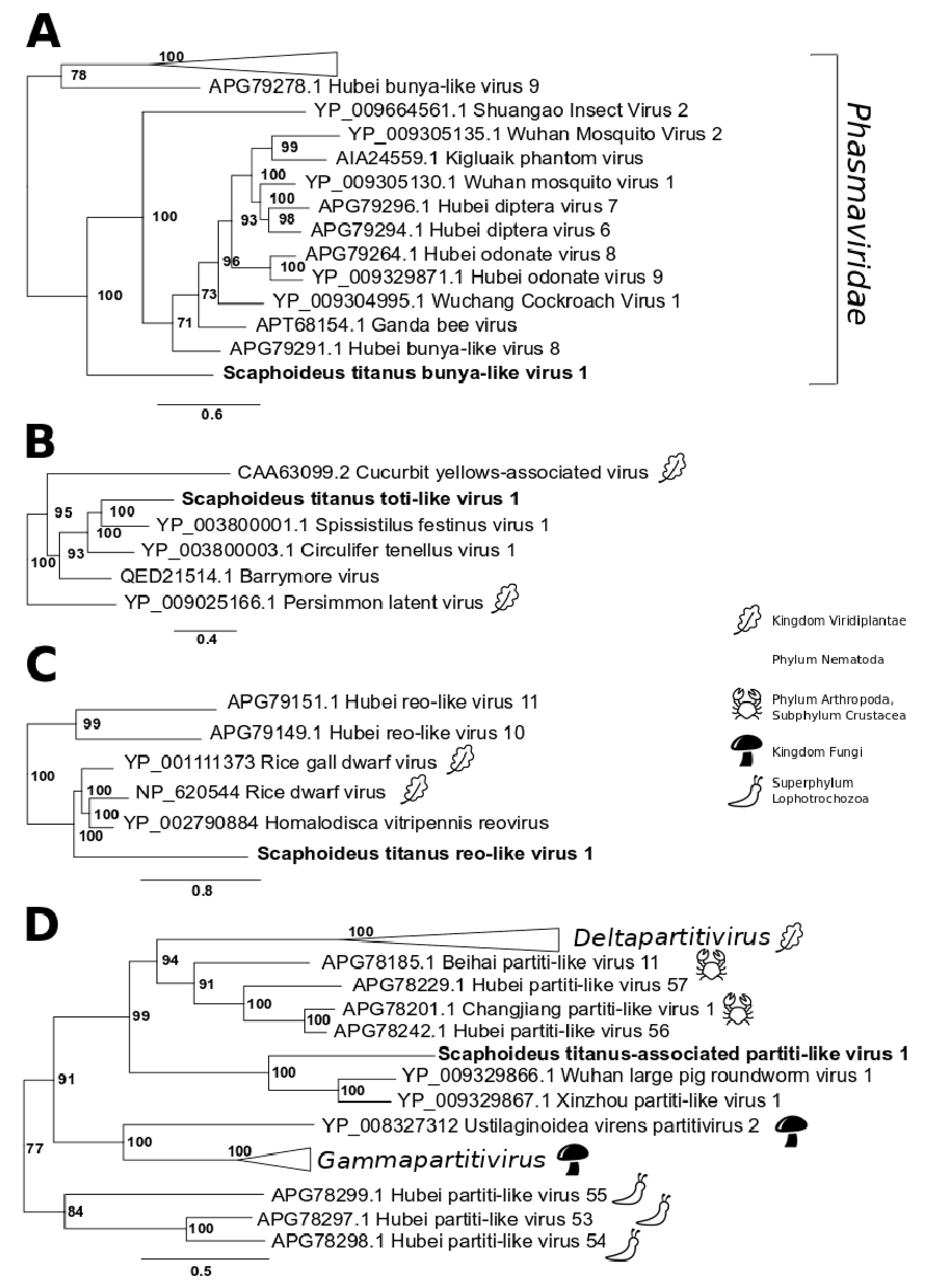
3.1. Single-Stranded Positive-Sense RNA Viruses
3.2. Single-Stranded Negative-Sense RNA Virus
3.3. Double-Stranded RNA Viruses
3.4. Virus Prevalence
3.5. Endogenous Viral Elements
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carton, Y.; Sorensen, C.; Smith, J.; Smith, E. Une coopération exemplaire entre entomologistes enançais et américains pendant la crise du Phylloxera en France (1868–1895). Ann. Société Entomol. Fr. NS 2007, 43, 103–125. [Google Scholar] [CrossRef] [Green Version]
- Papura, D.; Burban, C.; van Helden, M.; Giresse, X.; Nusillard, B.; Guillemaud, T.; Kerdelhué, C. Microsatellite and Mitochondrial Data Provide Evidence for a Single Major Introduction for the Neartic Leafhopper Scaphoideus titanus in Europe. PLoS ONE 2012, 7, e36882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maixner, M.; Pearson, R.; Boudon-Padieu, E.; Caudwell, A. Scaphoideus titanus, a possible vector of grapevine yellows in New York. Plant Dis. 1993, 77, 408–413. [Google Scholar] [CrossRef]
- Schvester, D.; Moutous, G.; Carle, P. Scaphoideus littoralis Ball (Homopt. Jassidae) cicadelle vectrice de la Flavescence dorée de la vigne. Rev. Zool. Agric. Appl. 1962, 61, 118–131. [Google Scholar]
- Krnjajić, S.; Mitrović, M.; Cvrković, T.; Jović, J.; Petrović, A.; Forte, V.; Angelini, E.; Toševski, I. Occurrence and distribution of Scaphoideus titanus in multiple outbreaks of “flavescence dorée” in Serbia. Bull. Insectol. 2012, 60, 197–198. [Google Scholar]
- Lessio, F.; Tota, F.; Alma, A. Tracking the dispersion of Scaphoideus titanus Ball (Hemiptera: Cicadellidae) from wild to cultivated grapevine: Use of a novel mark-capture technique. Bull. Entomol. Res. 2014, 104, 1–12. [Google Scholar] [CrossRef]
- Pavan, F.; Mori, N.; Bigot, G.; Zandigiacomo, P. Border effect in spatial distribution of Flavescence dorée affected grapevines and outside source of Scaphoideus titanus vectors. Bull. Insectol. 2012, 65, 281–290. [Google Scholar]
- Rossi, M.; Pegoraro, M.; Ripamonti, M.; Abbà, S.; Beal, D.; Giraudo, A.; Veratti, F.; Malembic-Maher, S.; Salar, P.; Bosco, D.; et al. Genetic diversity of Flavescence Dorée phytoplasmas at the vineyard scale. Appl. Environ. Microbiol. 2019, 85, e03123-18. [Google Scholar] [CrossRef] [Green Version]
- Martini, M.; Murari, E.; Mori, N.; Bertaccini, A. Identification and epidemic distribution of two flavescence dorée—related phytoplasmas in Veneto (Italy). Plant Dis. 1999, 83, 925–930. [Google Scholar] [CrossRef] [Green Version]
- Caudwell, A. Deux Années D’études sur la Flavescence Dorée Nouvelle Maladie Grave de la Vigne; Institut National de la Recherche Agronomique: Paris, France, 1957. [Google Scholar]
- Kriston, É.; Krizbai, L.; Szabó, G.; Bujdosó, B.; Orosz, S.; Dancsházy, Z.; Szőnyegi, S. First occurrence of grapevine flavescence doree in Hungary. Növényvédelem 2013, 49, 433–438. [Google Scholar]
- Jermini, M.; Gusberti, M.; Schaub, L.; Linder, C.; Gugerli, P.; Schärer, S.; Emery, S. Flavescence dorée and Scaphoideus titanus: Distribution and control in Switzerland. IOBCWPRS Bull. 2008, 36, 107–111. [Google Scholar]
- Weintraub, P.G.; Beanland, L. Insect vectors of phytoplasmas. Annu. Rev. Entomol. 2006, 51, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Morone, C.; Boveri, M.; Giosuè, S.; Gotta, P.; Rossi, V.; Scapin, I.; Marzachì, C. Epidemiology of Flavescence Dorée in Vineyards in Northwestern Italy. Phytopathology 2007, 97, 1422–1427. [Google Scholar] [CrossRef] [PubMed]
- Belli, G.; Bianco, P.; Conti, M. Grapevine yellows in Italy: Past, present and future. J. Plant Pathol. 2010, 92, 303–326. [Google Scholar]
- Chuche, J.; Thiéry, D. Biology and ecology of the Flavescence dorée vector Scaphoideus titanus: A review. Agron. Sustain. Dev. 2014, 34, 381–403. [Google Scholar] [CrossRef] [Green Version]
- Steffek, R.; Reisenzein, H.; Zeisner, N. Analysis of the pest risk from Grapevine flavescence dorée phytoplasma to Austrian viticulture. EPPO Bull. 2007, 37, 191–203. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health; Jeger, M.; Bragard, C.; Caffier, D.; Candresse, T.; Chatzivassiliou, E.; Dehnen-Schmutz, K.; Gilioli, G.; Jaques Miret, J.A.; MacLeod, A.; et al. Risk to plant health of Flavescence dorée for the EU territory. EFSA J. 2016, 14, e04603. [Google Scholar]
- Ignoffo, C.M. Entomopathogens as insecticides. Environ. Lett. 1975, 8, 23–40. [Google Scholar] [CrossRef]
- Bergoin, M.; Tijssen, P. Biological and molecular properties of densoviruses and their use in protein expression and biological control. In The Insect Viruses; Plenum Publishing, Inc.: New York, NY, USA, 1998; pp. 141–169. [Google Scholar]
- Jackson, T.A.; Crawford, A.M.; Glare, T.R. Oryctes virus—Time for a new look at a useful biocontrol agent. J. Invertebr. Pathol. 2005, 89, 91–94. [Google Scholar] [CrossRef]
- Marzachì, C.; Veratti, F.; Bosco, D. Direct PCR detection of phytoplasmas in experimentally infected insects. Ann. Appl. Biol. 1998, 133, 45–54. [Google Scholar] [CrossRef]
- BBMap. Available online: Sourceforge.net/projects/bbmap/ (accessed on 11 March 2019).
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Madan, A. CAP3: A DNA sequence assembly program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, M.; Lin, X.-D.; Tian, J.-H.; Chen, L.-J.; Chen, X.; Li, C.-X.; Qin, X.-C.; Li, J.; Cao, J.-P.; Eden, J.-S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- King, A.M.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Iflaviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M., Lefkowitz, E., Adams, M.J., Carstens, E.B., Eds.; Elsevier: San Diego, CA, USA, 2012; Volume 9, p. 848. [Google Scholar]
- Spear, A.; Sisterson, M.S.; Yokomi, R.; Stenger, D.C. Plant-feeding insects harbor double-stranded RNA viruses encoding a novel proline-alanine rich protein and a polymerase distantly related to that of fungal viruses. Virology 2010, 404, 304–311. [Google Scholar] [CrossRef] [Green Version]
- Coffin, R.; Coutts, R. Relationships among Trialeurodes vaporariorum—Transmitted Yellowing Viruses from Europe and North America. J. Phytopathol. 1995, 143, 375–380. [Google Scholar] [CrossRef]
- Ito, T.; Suzaki, K.; Nakano, M. Genetic characterization of novel putative rhabdovirus and dsRNA virus from Japanese persimmon. J. Gen. Virol. 2013, 94, 1917–1921. [Google Scholar] [CrossRef] [Green Version]
- Stenger, D.C.; Sisterson, M.S.; Krugner, R.; Backus, E.A.; Hunter, W.B. A new Phytoreovirus infecting the glassy-winged sharpshooter (Homalodisca vitripennis). Virology 2009, 386, 469–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitbart, M.; Salamon, P.; Andresen, B.; Mahaffy, J.M.; Segall, A.M.; Mead, D.; Azam, F.; Rohwer, F. Genomic analysis of uncultured marine viral communities. Proc. Natl. Acad. Sci. USA 2002, 99, 14250–14255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obbard, D.J. Expansion of the metazoan virosphere: Progress, pitfalls, and prospects. Curr. Opin. Virol. 2018, 31, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmonds, P.; Adams, M.J.; Benkő, M.; Breitbart, M.; Brister, J.R.; Carstens, E.B.; Davison, A.J.; Delwart, E.; Gorbalenya, A.E.; Harrach, B.; et al. Consensus statement: Virus taxonomy in the age of metagenomics. Nat. Rev. Microbiol. 2017, 15, 161. [Google Scholar] [CrossRef] [PubMed]
- Bonning, B.C. The Insect Virome: Opportunities and Challenges. Curr. Issues Mol. Biol. 2019, 34, 1–12. [Google Scholar] [PubMed]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M. Ictv Report Consortium ICTV Virus Taxonomy Profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef]
- Lara Pinto, A.Z.; de Santos de Carvalho, M.; de Melo, F.L.; Ribeiro, A.L.M.; Morais Ribeiro, B.; Dezengrini Slhessarenko, R. Novel viruses in salivary glands of mosquitoes from sylvatic Cerrado, Midwestern Brazil. PLoS ONE 2017, 12, e0187429. [Google Scholar] [CrossRef]
- Pettersson, H.-O.J.; Shi, M.; Eden, J.-S.; Holmes, C.E.; Hesson, C.J. Meta-Transcriptomic Comparison of the RNA Viromes of the Mosquito Vectors Culex pipiens and Culex torrentium in Northern Europe. Viruses 2019, 11, 1033. [Google Scholar] [CrossRef] [Green Version]
- Trivellone, V.; Illinois Natural History Survey, Champaign, IL, USA. Personal Communication, 2018.
- Keane, R.M.; Crawley, M.J. Exotic plant invasions and the enemy release hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Colautti, R.I.; Ricciardi, A.; Grigorovich, I.A.; MacIsaac, H.J. Is invasion success explained by the enemy release hypothesis? Ecol. Lett. 2004, 7, 721–733. [Google Scholar] [CrossRef]
- Medd, N.C.; Fellous, S.; Waldron, F.M.; Xuéreb, A.; Nakai, M.; Cross, J.V.; Obbard, D.J. The virome of Drosophila suzukii, an invasive pest of soft fruit. Virus Evol. 2018, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Souza-Neto, J.; Cosme, R.T.; Rovira, J.; Ortiz, A.; Pascale, J.M.; Dimopoulos, G. Reciprocal tripartite interactions between the Aedes aegypti midgut microbiota, innate immune system and dengue virus influences vector competence. PLoS Negl. Trop. Dis. 2012, 6, e1561. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef] [PubMed]
- Hunter, W.; Glick, E.; Paldi, N.; Bextine, B. Advances in RNA interference: dsRNA Treatment in Trees and Grapevines for Insect Pest Suppression. Southwest. Entomol. 2012, 37, 85–87. [Google Scholar] [CrossRef]
- Killiny, N.; Hajeri, S.; Tiwari, S.; Gowda, S.; Stelinski, L.L. Double-Stranded RNA Uptake through Topical Application, Mediates Silencing of Five CYP4 Genes and Suppresses Insecticide Resistance in Diaphorina citri. PLoS ONE 2014, 9, 1–8. [Google Scholar]
- Cook, S.; Chung, B.Y.-W.; Bass, D.; Moureau, G.; Tang, S.; McAlister, E.; Culverwell, C.L.; Glücksman, E.; Wang, H.; Brown, T.D.K.; et al. Novel Virus Discovery and Genome Reconstruction from Field RNA Samples Reveals Highly Divergent Viruses in Dipteran Hosts. PLoS ONE 2013, 8, e80720. [Google Scholar] [CrossRef] [Green Version]
- Fauver, J.R.; Grubaugh, N.D.; Krajacich, B.J.; Weger-Lucarelli, J.; Lakin, S.M.; Fakoli, L.S., III; Bolay, F.K.; Diclaro, J.W., II; Dabiré, K.R.; Foy, B.D.; et al. West African Anopheles gambiae mosquitoes harbor a taxonomically diverse virome including new insect-specific flaviviruses, mononegaviruses, and totiviruses. Virology 2016, 498, 288–299. [Google Scholar] [CrossRef] [Green Version]
- Zakrzewski, M.; Rašić, G.; Darbro, J.; Krause, L.; Poo, Y.S.; Filipović, I.; Parry, R.; Asgari, S.; Devine, G.; Suhrbier, A. Mapping the virome in wild-caught Aedes aegypti from Cairns and Bangkok. Sci. Rep. 2018, 8, 4690. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Asad, S.; Khromykh, A.A.; Asgari, S. Cell fusing agent virus and dengue virus mutually interact in Aedes aegypti cell lines. Sci. Rep. 2017, 7, 6935. [Google Scholar] [CrossRef] [Green Version]
- Sadeghi, M.; Altan, E.; Deng, X.; Barker, C.M.; Fang, Y.; Coffey, L.L.; Delwart, E. Virome of > 12 thousand Culex mosquitoes from throughout California. Virology 2018, 523, 74–88. [Google Scholar] [CrossRef]
- Rosario, K.; Capobianco, H.; Ng, T.F.F.; Breitbart, M.; Polston, J.E. RNA viral metagenome of whiteflies leads to the discovery and characterization of a whitefly-transmitted carlavirus in North America. PLoS ONE 2014, 9, e86748. [Google Scholar] [CrossRef] [PubMed]
- Nouri, S.; Salem, N.; Nigg, J.C.; Falk, B.W. Diverse Array of New Viral Sequences Identified in Worldwide Populations of the Asian Citrus Psyllid (Diaphorina citri) Using Viral Metagenomics. J. Virol. 2016, 90, 2434–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wamonje, F.O.; Michuki, G.N.; Braidwood, L.A.; Njuguna, J.N.; Musembi Mutuku, J.; Djikeng, A.; Harvey, J.J.W.; Carr, J.P. Viral metagenomics of aphids present in bean and maize plots on mixed-use farms in Kenya reveals the presence of three dicistroviruses including a novel Big Sioux River virus-like dicistrovirus. Virol. J. 2017, 14, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atoni, E.; Wang, Y.; Karungu, S.; Waruhiu, C.; Zohaib, A.; Obanda, V.; Agwanda, B.; Mutua, M.; Xia, H.; Yuan, Z. Metagenomic virome analysis of Culex mosquitoes from Kenya and China. Viruses 2018, 10, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanfack-Minkeu, F.; Mitri, C.; Bischoff, E.; Belda, E.; Casademont, I.; Vernick, K.D. Interaction of RNA viruses of the natural virome with the African malaria vector, Anopheles coluzzii. Sci. Rep. 2019, 9, 6319. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RNA-seq LibrariesID | Country | State/Region | Sampling Sites | Latitude (Decimal Degrees) | Longitude (Decimal Degrees) | Sampling Date (yyyy/mm/dd) | Number of Sampled Individuals | Positive to Scaphoideus titanus Iflavirus 1 | Positive to Scaphoideus titanustoti-Like Virus 1 | |
|---|---|---|---|---|---|---|---|---|---|---|
| St_HU | Hungary | Pest | Monor | 47.354861 | 19.472177 | 2018-07-12 | 20 | 0 | 0 | |
| St_HU | Hungary | Pest | Gomba | 47.349622 | 19.505863 | 2018-07-12 | 18 | 0 | 0 | |
| St_FR | France | Burgundy | Corgoloin | 47.095667 | 4.908233 | 2018-07-06 | 20 | 0 | 8 | |
| St_FR | France | Dordogne | Saint-Nexans | 44.472132 | 0.345279 | 2018-07-10 | 18 | 1 | 1 | |
| St_IT1 | Italy | Piedmont | Asti | 44.921854 | 8.195758 | 2018-07-31 | 4 | 0 | 0 | |
| St_IT1 | Italy | Piedmont | Montà | 44.807434 | 7.965787 | 2018-07-31 | 12 | 1 | 0 | |
| St_IT1 | Italy | Piedmont | Portacomaro | 44.962898 | 8.26072 | 2018-08-03 | 16 | 0 | 0 | |
| St_IT1 | Italy | Piedmont | Cisterna | 44.82591 | 8.008371 | 2018-08-03 | 4 | 0 | 0 | |
| St_IT1 | Italy | Lombardy | Ome | 45.6302972 | 10.129867 | 2018-07-11 | 4 | 0 | 0 | |
| St_IT1 | Italy | Veneto | Verona | 45.4227833 | 11.025431 | 2018-08-08 | 8 | 0 | 0 | |
| St_IT2 | Italy | Friuli-Venezia Giulia | Togliano | 46.1129556 | 13.411311 | 2018-07-30 | 22 | 0 | 2 | |
| St_IT2 | Italy | Friuli-Venezia Giulia | Savorgnano al Torre | 46.161786 | 13.286864 | 2018-07-31 | 24 | 0 | 1 | |
| St_IT2 | Italy | Abruzzo | Vacri | 42.290333 | 14.223861 | 2018-07-16 | 12 | 0 | 2 | |
| St_CH | Switzerland | Canton Vaud | Gland | 46.42616 | 6.28217 | 2018-08-09 | 16 | 0 | 4 | |
| St_CH | Switzerland | Canton Valais | Pramagnon | 46.24987 | 7.45371 | 2018-08-09 | 16 | 0 | 0 | |
| 214 | 2 | 18 | Total | |||||||
| St_USA | USA | Illinois | Muncie | 40.0893 | -87.8323 | 2018-06-11 | 3 | |||
| Insects for DNA extraction1 | Country | State/Region | Sampling Sites | Latitude (Decimal Degrees) | Longitude (Decimal Degrees) | Sampling Date (yyyy/mm/dd) | Number of Sampled Individuals | |||
| Italy | Piedmont | Montà | 44.807434 | 7.965787 | 2018-07-31 | 2 | ||||
| Italy | Piedmont | Dogliani | 44.5362837 | 7.9312913 | 2018-08-20 | 2 |
| Library ID | Accession Numbers | Sequence Length (bp) | Best Blastx Hit Description | % Identity | % Query Coverage | RPKM | Tentative Name | Genome |
|---|---|---|---|---|---|---|---|---|
| St_IT1 | MN982380 | 10729 | YP_009129265.1 polyprotein [Graminella nigrifrons virus 1] | 51 | 83 | 0.01 | Scaphoideus titanus iflavirus 1 | +ssRNA (monopartite) |
| St_IT2 | MN982382 | 7847 | YP_003800001.1 RNA-directed RNA polymerase partial [Spissistilus festinus virus 1] | 42 | 37 | 0.002 | Scaphoideus titanus toti-like virus 1 | dsRNA (monopartite) |
| St_CH | MN982383 | 7839 | YP_003800001.1 RNA-directed RNA polymerase partial [Spissistilus festinus virus 1] | 42 | 37 | 0.032 | Scaphoideus titanus toti-like virus 1 | dsRNA (monopartite) |
| St_FR | MN982379 | 10693 | YP_009129265.1 polyprotein [Graminella nigrifrons virus 1] | 51 | 83 | 0.004 | Scaphoideus titanus iflavirus 1 | +ssRNA (monopartite) |
| St_FR | MN982381 | 7871 | YP_003800001.1 RNA-directed RNA polymerase partial [Spissistilus festinus virus 1] | 42 | 37 | 0.023 | Scaphoideus titanus toti-like virus 1 | dsRNA (monopartite) |
| St_USA | MN982384 | 4361 | API65464.1 glycoprotein precursor [Sanxia Water Strider Virus 2] | 25 | 12 | 1.016 | Scaphoideus titanus bunya-like virus 1 | −ssRNA (tripartite): segment M |
| St_USA | MN982385 | 1458 | APG79297.1 putative nucleoprotein [Hubei diptera virus 7] | 27 | 43 | 0.9 | Scaphoideus titanus bunya-like virus 1 | −ssRNA (tripartite): segment S |
| St_USA | MN982386 | 7740 | APT68154.1 RNA-dependent RNA polymerase [Ganda bee virus] | 33 | 74 | 0.359 | Scaphoideus titanus bunya-like virus 1 | −ssRNA (tripartite): segment L |
| St_USA | MN982387 | 10519 | YP_009129265.1 polyprotein [Graminella nigrifrons virus 1] | 51 | 85 | 0.045 | Scaphoideus titanus iflavirus 2 | +ssRNA (monopartite) |
| St_USA | MN982388 | 1455 | AWA82259.1 hypothetical protein, partial [Naganuma virus] | 38 | 91 | 0.145 | Scaphoideus titanus sobemo-like virus 1 | +ssRNA (bipartite) |
| St_USA | MN982389 | 3213 | YP_009329970.1 hypothetical protein [Hubei sobemo-like virus 26] | 60 | 99 | 0.103 | Scaphoideus titanus sobemo-like virus 1 | +ssRNA (bipartite) |
| St_USA | MN982390 | 1550 | AWA82259.1 hypothetical protein partial [Naganuma virus] | 36 | 77 | 0.231 | Scaphoideus titanus sobemo-like virus 2 | +ssRNA (bipartite) |
| St_USA | MN982391 | 3100 | YP_009330073.1 hypothetical protein 2 [Hubei sobemo-like virus 24] | 46 | 97 | 0.172 | Scaphoideus titanus sobemo-like virus 2 | +ssRNA (bipartite) |
| St_USA | MN982393 | 1303 | YP_009329884.1 hypothetical protein [Wuhan large pig roundworm virus 1] | 35 | 61 | 0.227 | Scaphoideus titanus-associated partiti-like virus 1 | dsRNA (bipartite) |
| St_USA | MN982392 | 1558 | AVV63192.1 RNA-directed RNA polymerase [Wuhan large pig roundworm virus] | 44 | 87 | 0.272 | Scaphoideus titanus-associated partiti-like virus 1 | dsRNA (bipartite) |
| St_USA | MN982394 | 5239 | YP_009337778.1 RdRp [Hubei permutotetra-like virus 9] | 39 | 39 | 0.764 | Scaphoideus titanus permutotetra-like virus 1 | +ssRNA (monopartite) |
| St_USA | MN982397 | 1240 | ADN64742.1 non-structural protein [Homalodisca vitripennis reo-like virus] | 79 | 58 | 2.378 | Scaphoideus titanus reo-like virus 1 | dsRNA (multipartite): segment 10 |
| St_USA | MN982398 | 1867 | ADN64768.1 minor core protein [Homalodisca vitripennis reo-like virus] | 83 | 42 | 1.146 | Scaphoideus titanus reo-like virus 1 | dsRNA (multipartite): segment 7 |
| St_USA | MN982399 | 2937 | ADN64783.1 minor core protein [Homalodisca vitripennis reo-like virus] | 52 | 49 | 2.661 | Scaphoideus titanus reo-like virus 1 | dsRNA (multipartite): segment 5 |
| St_USA | MN982395 | 4957 | ADN64816.1 RNA-directed RNA polymerase [Homalodisca vitripennis reo-like virus] | 87 | 61 | 5.053 | Scaphoideus titanus reo-like virus 1 | dsRNA (multipartite): segment 1 |
| St_USA | MN982400 | 3560 | YP_002790885.1 RNA-binding protein [Homalodisca vitripennis reo-like virus] | 96 | 49 | 5.444 | Scaphoideus titanus reo-like virus 1 | dsRNA (multipartite): segment 2 |
| St_USA | MN982401 | 1416 | Q85451.1 RecName: Full=Outer capsid protein P8 | 62 | 88 | 1.679 | Scaphoideus titanus reo-like virus 1 | dsRNA (multipartite): segment 8 |
| St_USA | MN982402 | 3179 | ADN64799.1 major core protein [Homalodisca vitripennis reo-like virus] | 95 | 55 | 1.901 | Scaphoideus titanus reo-like virus 1 | dsRNA (multipartite): segment 3 |
| St_USA | MN982403 | 2648 | YP_002790887.1 zinc-finger protein [Homalodisca vitripennis reo-like virus] | 73 | 30 | 3.571 | Scaphoideus titanus reo-like virus 1 | dsRNA (multipartite): segment 4 |
| St_USA | MN982404 | 1778 | YP_002790889.1 non-structural protein [Homalodisca vitripennis reo-like virus] | 78 | 33 | 1.196 | Scaphoideus titanus reo-like virus 1 | dsRNA (multipartite): segment 6 |
| St_USA | MN982396 | 1179 | YP_002790892.1 non-structural protein [Homalodisca vitripennis reo-like virus] | 68 | 47 | 1.802 | Scaphoideus titanus reo-like virus 1 | dsRNA (multipartite): segment 9 |
| St_USA | MN982405 | 798 | YP_002790894.1 non-structural protein [Homalodisca vitripennis reo-like virus] | 48 | 45 | 4.192 | Scaphoideus titanus reo-like virus 1 | dsRNA (multipartite): segment 11 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ottati, S.; Chiapello, M.; Galetto, L.; Bosco, D.; Marzachì, C.; Abbà, S. New Viral Sequences Identified in the Flavescence Dorée Phytoplasma Vector Scaphoideus titanus. Viruses 2020, 12, 287. https://doi.org/10.3390/v12030287
Ottati S, Chiapello M, Galetto L, Bosco D, Marzachì C, Abbà S. New Viral Sequences Identified in the Flavescence Dorée Phytoplasma Vector Scaphoideus titanus. Viruses. 2020; 12(3):287. https://doi.org/10.3390/v12030287
Chicago/Turabian StyleOttati, Sara, Marco Chiapello, Luciana Galetto, Domenico Bosco, Cristina Marzachì, and Simona Abbà. 2020. "New Viral Sequences Identified in the Flavescence Dorée Phytoplasma Vector Scaphoideus titanus" Viruses 12, no. 3: 287. https://doi.org/10.3390/v12030287
APA StyleOttati, S., Chiapello, M., Galetto, L., Bosco, D., Marzachì, C., & Abbà, S. (2020). New Viral Sequences Identified in the Flavescence Dorée Phytoplasma Vector Scaphoideus titanus. Viruses, 12(3), 287. https://doi.org/10.3390/v12030287