Characterization of Port Bolivar Virus, a Novel Entomobirnavirus (Birnaviridae) Isolated from Mosquitoes Collected in East Texas, USA


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Mosquito Collection
2.2. Culture Methods
2.3. Immunofluorescent Studies
2.4. Transmission Electron Microscopy (TEM)
2.5. RNA Extraction and Next-Generation Sequencing
2.6. Phylogenetic Analysis
3. Results
3.1. Virus Isolation
3.2. Immunofluorescent Studies
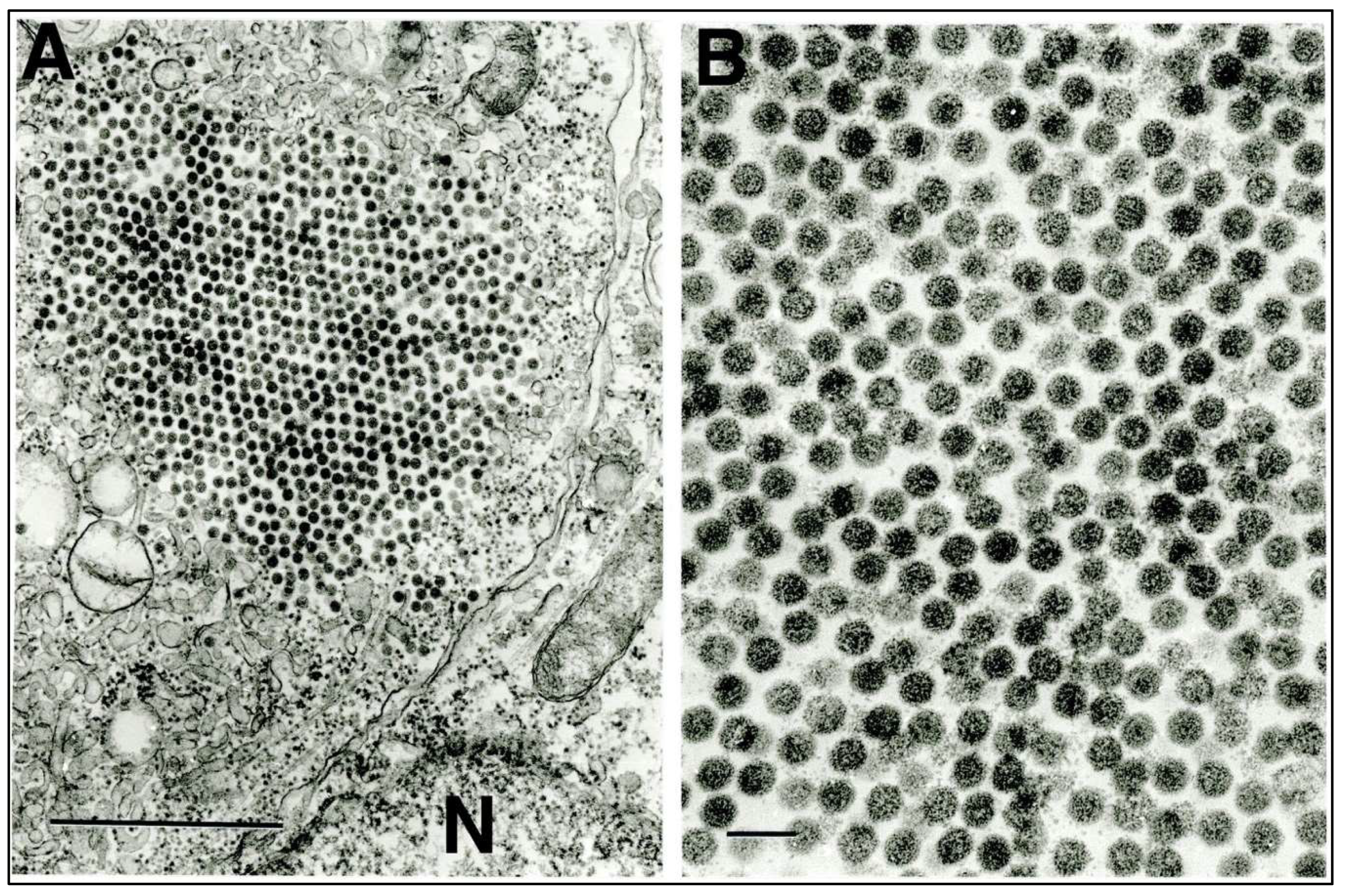
3.3. Transmission Electron Microscopy (TEM)
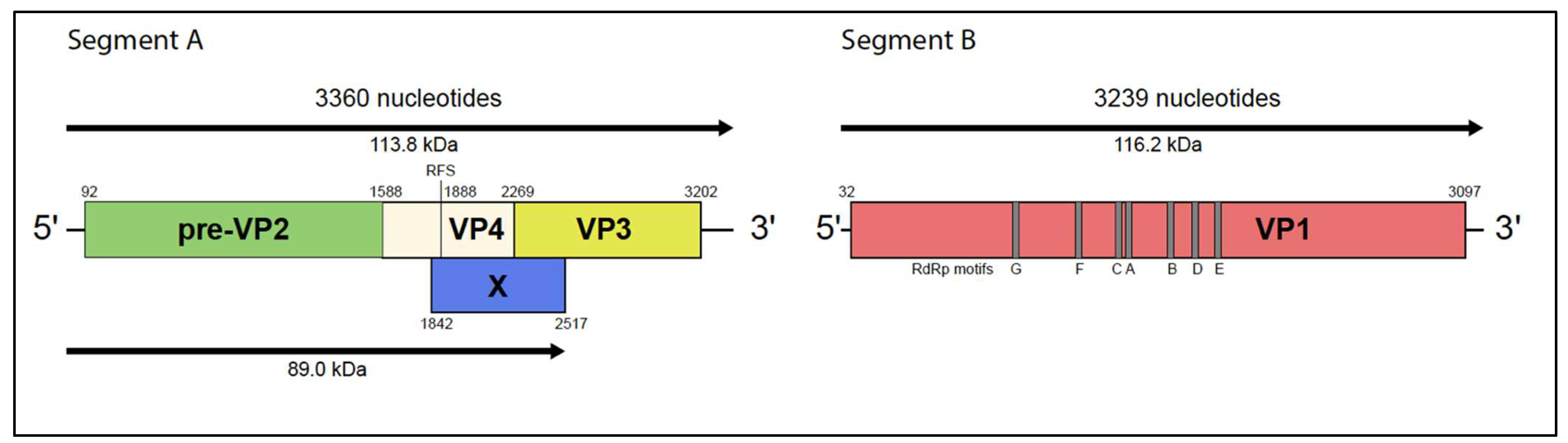
3.4. Genomic Characterization
3.5. Amino Acid Sequence Identities
3.6. Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Delmas:, B.; Attoui, H.; Ghosh, S.; Malik, Y.S.; Mundt, E.; Vakharia, V.N. ICTV Report Comm., ICTV virus taxonomy profile: Birnaviridae. J. Gen. Virol. 2019, 100, 5–6. [Google Scholar] [CrossRef] [PubMed]
- Teninges, D.; Ohanessian, A.; Richard-Molard, C.; Contamine, D. Isolation and Biological Properties of Drosophila X Virus. J. Gen. Virol. 1979, 42, 241–254. [Google Scholar] [CrossRef]
- Huang, Y.; Mi, Z.; Zhuang, L.; Ma, M.; An, X.; Liu, W.; Cao, W.; Tong, Y. Presence of entomobirnaviruses in Chinese mosquitoes in the absence of Dengue virus co-infection. J. Gen. Virol. 2013, 94, 663–667. [Google Scholar] [CrossRef] [PubMed]
- Marklewitz, M.; Gloza-Rausch, F.; Kurth, A.; Kummerer, B.M.; Drosten, C.; Junglen, S. First isolation of an Entomobirnavirus from free-living insects. J. Gen. Virol. 2012, 93, 2431–2435. [Google Scholar] [CrossRef]
- Webster, C.L.; Longdon, B.; Lewis, S.H.; Obbard, D.J. Twenty-Five New Viruses Associated with the Drosophilidae (Diptera). Evol. Bioinform. Online 2016, 12, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Cook, S.; Chung, B.Y.; Bass, D.; Moureau, G.; Tang, S.; McAlister, E.; Culverwell, C.L.; Glucksman, E.; Wang, H.; Brown, T.D.; et al. Novel virus discovery and genome reconstruction from field RNA samples reveals highly divergent viruses in dipteran hosts. PLoS ONE 2013, 8, e80720. [Google Scholar] [CrossRef] [Green Version]
- Vancini, R.; Paredes, A.; Ribeiro, M.; Blackburn, K.; Ferreira, D.; Kononchik, J.P., Jr.; Hernandez, R.; Brown, D. Espirito Santo virus: A new birnavirus that replicates in insect cells. J. Virol. 2012, 86, 2390–2399. [Google Scholar] [CrossRef] [Green Version]
- Kelly, D.C.; Ayres, M.D.; Howard, S.C.; Lescott, T.; Arnold, M.K.; Seeley, N.D.; Primrose, S.B. Isolation of a bisegmented double-stranded RNA virus from Thirlmere reservoir. J. Gen. Virol. 1982, 62, 313–322. [Google Scholar] [CrossRef]
- Tesh, R.B. A method for the isolation and identification of dengue viruses, using mosquito cell cultures. Am. J. Trop. Med. Hyg. 1979, 28, 1053–1059. [Google Scholar] [CrossRef]
- Popov, V.L.; Tesh, R.B.; Weaver, S.C.; Vasilakis, N. Electron Microscopy in Discovery of Novel and Emerging Viruses from the Collection of the World Reference Center for Emerging Viruses and Arboviruses (WRCEVA). Viruses 2019, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- Lohse, M.; Bolger, A.M.; Nagel, A.; Fernie, A.R.; Lunn, J.E.; Stitt, M.; Usadel, B. RobiNA: A user-friendly, integrated software solution for RNA-Seq-based transcriptomics. Nucleic Acids Res. 2012, 40, W622–W627. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.T.; Wong, K.; Jackman, S.D.; Schein, J.E.; Jones, S.J.; Birol, I. ABySS: A parallel assembler for short read sequence data. Genome Res. 2009, 19, 1117–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, J.T.; Thorvaldsdottir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotech. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Whelan, S.; Goldman, N. A general empirical model of protein evolution derived from multiple protein families using a maximum-likelihood approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Chung, H.K.; Kordyban, S.; Cameron, L.; Dobos, P. Sequence analysis of the bicistronic Drosophila X virus genome segment A and its encoded polypeptides. Virology 1996, 225, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Vakharia, V.N.; Tao, Y.J. The structure of a birnavirus polymerase reveals a distinct active site topology. Proc. Natl. Acad. Sci. USA 2007, 104, 7385–7390. [Google Scholar] [CrossRef] [Green Version]
- Shwed, P.S.; Dobos, P.; Cameron, L.A.; Vakharia, V.N.; Duncan, R. Birnavirus VP1 proteins form a distinct subgroup of RNA-dependent RNA polymerases lacking a GDD motif. Virology 2002, 296, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Petit, S.; Lejal, N.; Huet, J.C.; Delmas, B. Active residues and viral substrate cleavage sites of the protease of the birnavirus infectious pancreatic necrosis virus. J. Virol. 2000, 74, 2057–2066. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, B.; Chevalier, C.; Henry, C.; Huet, J.C.; Petit, S.; Lepault, J.; Boot, H.; Delmas, B. The capsid of infectious bursal disease virus contains several small peptides arising from the maturation process of pVP2. J. Virol. 2002, 76, 2393–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Costa, B.; Soignier, S.; Chevalier, C.; Henry, C.; Thory, C.; Huet, J.C.; Delmas, B. Blotched snakehead virus is a new aquatic birnavirus that is slightly more related to avibirnavirus than to aquabirnavirus. J. Virol. 2003, 77, 719–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galloux, M.; Chevalier, C.; Henry, C.; Huet, J.C.; Costa, B.D.; Delmas, B. Peptides resulting from the pVP2 C-terminal processing are present in infectious pancreatic necrosis virus particles. J. Gen. Virol. 2004, 85, 2231–2236. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Virus | PTBV | CaZV | CuYV | MXV | ESV | DXV | ERV | IPNV | TABV | VTABV | IBDV | BSHV | LCBV | RBV | DBV | TV-1 | JmBLV |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Entomobirnavirus | PTBV | |||||||||||||||||
| CaZV | 87.8 | |||||||||||||||||
| CuYV | 85.8 | 92.2 | ||||||||||||||||
| MXV * | 85.9 | 92.5 | 98.5 | |||||||||||||||
| ESV | 86.1 | 92.1 | 98.8 | 98.1 | ||||||||||||||
| DXV * | 72.3 | 72.5 | 72.5 | 72.8 | 72.5 | |||||||||||||
| ERV | 69.4 | 71.3 | 71.6 | 71.7 | 71.6 | 84.7 | ||||||||||||
| Aquabirnavirus | IPNV * | 28.4 | 28.4 | 28.3 | 28.1 | 28.1 | 29.2 | 28.7 | ||||||||||
| TABV | 28.6 | 28.0 | 28.4 | 28.3 | 28.3 | 28.1 | 28.0 | 90.8 | ||||||||||
| VTABV | 29.0 | 29.0 | 28.7 | 28.6 | 28.6 | 29.4 | 29.1 | 94.5 | 90.4 | |||||||||
| Avibirnavirus | IBDV * | 29.1 | 28.8 | 28.1 | 28.1 | 27.9 | 28.1 | 27.7 | 48.0 | 47.4 | 47.4 | |||||||
| Blosnavirus | BSHV * | 29.6 | 29.8 | 29.9 | 30.1 | 29.8 | 29.5 | 29.8 | 49.2 | 48.2 | 49.0 | 52.3 | ||||||
| LCBV * | 28.6 | 29.0 | 29.5 | 29.6 | 29.1 | 28.6 | 28.4 | 50.4 | 48.9 | 49.7 | 51.0 | 62.2 | ||||||
| Ronavirus | RBV * | 26.1 | 25.1 | 24.7 | 25.0 | 24.9 | 25.0 | 24.9 | 30.9 | 31.3 | 31.0 | 30.2 | 31.7 | 30.7 | ||||
| Dronavirus | DBV * | 24.5 | 23.4 | 23.9 | 24.2 | 23.9 | 23.4 | 22.4 | 29.5 | 29.2 | 29.8 | 27.7 | 29.6 | 29.6 | 24.6 | |||
| Telnavirus | TV-1 * | 24.5 | 24.2 | 23.9 | 23.5 | 23.6 | 25.0 | 25.1 | 30.3 | 30.5 | 30.1 | 31.1 | 31.8 | 33.3 | 28.7 | 25.0 | ||
| unassigned | JmBLV | 28.1 | 27.3 | 27.3 | 27.3 | 27.3 | 27.3 | 28.0 | 31.7 | 31.0 | 31.1 | 30.2 | 30.6 | 30.7 | 26.9 | 23.2 | 26.4 |
| Genus | Virus | PTBV | CaZV | CuYV | MXV | ESV | DXV | ERV | IPNV | TABV | VTABV | IBDV | BSHV | LCBV | RBV | DBV | TV-1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Entomobirnavirus | PTBV | ||||||||||||||||
| CaZV | 91.7 | ||||||||||||||||
| CuYV | 85.9 | 89.2 | |||||||||||||||
| MXV * | 86.2 | 89.7 | 98.4 | ||||||||||||||
| ESV | 85.1 | 88.1 | 98.4 | 97.3 | |||||||||||||
| DXV * | 69.6 | 70.5 | 69.3 | 68.8 | 68.9 | ||||||||||||
| ERV | 71.3 | 72.7 | 70.7 | 70.4 | 70.4 | 86.1 | |||||||||||
| Aquabirnavirus | IPNV * | 25.2 | 25.2 | 25.0 | 24.9 | 24.8 | 24.8 | 25.6 | |||||||||
| TABV | 25.1 | 24.5 | 24.4 | 24.4 | 24.2 | 24.3 | 25.1 | 84.7 | |||||||||
| VTABV | 25.3 | 25.0 | 24.5 | 24.4 | 24.4 | 24.3 | 25.3 | 88.8 | 86.8 | ||||||||
| Avibirnavirus | IBDV * | 25.1 | 24.8 | 25.2 | 25.2 | 25.0 | 25.6 | 27.0 | 35.5 | 35.5 | 36.1 | ### | |||||
| Blosnavirus | BSHV * | 26.6 | 26.6 | 26.4 | 26.3 | 26.4 | 25.6 | 27.7 | 35.3 | 36.2 | 35.8 | 39.8 | |||||
| LCBV * | 24.5 | 25.0 | 25.0 | 24.9 | 24.9 | 23.8 | 25.0 | 34.5 | 34.8 | 33.8 | 39.6 | 51.5 | |||||
| Ronavirus | RBV * | 24.9 | 24.7 | 25.0 | 24.5 | 24.7 | 25.7 | 26.5 | 27.5 | 26.6 | 27.0 | 27.7 | 28.2 | 26.3 | |||
| Dronavirus | DBV * | 24.9 | 24.5 | 24.0 | 24.0 | 24.0 | 24.3 | 24.0 | 27.5 | 27.7 | 28.7 | 29.2 | 27.4 | 27.7 | 25.7 | ||
| Telnavirus | TV-1 * | 21.5 | 21.6 | 20.9 | 21.1 | 20.5 | 22.6 | 22.6 | 23.8 | 24.3 | 22.9 | 26.0 | 26.7 | 26.2 | 25.7 | 23.1 |
| Virus Name (Abbreviation) | Source | Locality | Accession Numbers | Reference |
|---|---|---|---|---|
| Drosophila X virus (DXV) | Drosophila melanogaster cell culture | France | U60650; AF196645 | [2] |
| Eridge virus (ERV) | D. melanogaster cell culture | UK | KU754527; KU754528 | [5] |
| Culicine-associated Z virus (CaZV) | Ochlerotatus caspius and Oc. detritus | France | KF298271; KF298272 | [6] |
| Espirito Santo virus (ESV) | Aedes albopictus cell culture | Brazil | JN589003; NJ589002 | [7] |
| Mosquito X virus (MXV) | Anopheles sinensis | China | JX403941; JX403942 | [3] |
| Culex Y virus (CuYV) | Culex pipiens complex | Germany | JQ659254; JQ659255 | [4] |
| Thirlmere virus | Water | UK | Not available | [8] |
| Port Bolivar virus (PTBV) | Aedes sollcitans | USA | MT263973 MT263974 | Present paper |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tesh, R.B.; Bolling, B.G.; Guzman, H.; Popov, V.L.; Wilson, A.; Widen, S.G.; Wood, T.G.; Walker, P.J.; Vasilakis, N. Characterization of Port Bolivar Virus, a Novel Entomobirnavirus (Birnaviridae) Isolated from Mosquitoes Collected in East Texas, USA. Viruses 2020, 12, 390. https://doi.org/10.3390/v12040390
Tesh RB, Bolling BG, Guzman H, Popov VL, Wilson A, Widen SG, Wood TG, Walker PJ, Vasilakis N. Characterization of Port Bolivar Virus, a Novel Entomobirnavirus (Birnaviridae) Isolated from Mosquitoes Collected in East Texas, USA. Viruses. 2020; 12(4):390. https://doi.org/10.3390/v12040390
Chicago/Turabian StyleTesh, Robert B., Bethany G. Bolling, Hilda Guzman, Vsevolod L. Popov, Ashley Wilson, Steven G. Widen, Thomas G. Wood, Peter J. Walker, and Nikos Vasilakis. 2020. "Characterization of Port Bolivar Virus, a Novel Entomobirnavirus (Birnaviridae) Isolated from Mosquitoes Collected in East Texas, USA" Viruses 12, no. 4: 390. https://doi.org/10.3390/v12040390
APA StyleTesh, R. B., Bolling, B. G., Guzman, H., Popov, V. L., Wilson, A., Widen, S. G., Wood, T. G., Walker, P. J., & Vasilakis, N. (2020). Characterization of Port Bolivar Virus, a Novel Entomobirnavirus (Birnaviridae) Isolated from Mosquitoes Collected in East Texas, USA. Viruses, 12(4), 390. https://doi.org/10.3390/v12040390