ORF Ι of Mycovirus SsNSRV-1 is Associated with Debilitating Symptoms of Sclerotinia sclerotiorum




Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Materials and Growth Conditions
2.2. DNA and RNA Extraction, cDNA Synthesis
2.3. Molecular Cloning, Vector Construction and Genetic Manipulation
2.4. Morphological Observation and Pathogenicity Assay
2.5. Preparation of cDNA Library
2.6. Transcriptome Analyses
2.7. Real-Time Quantitative Reverse Transcription PCR Analysis
3. Results
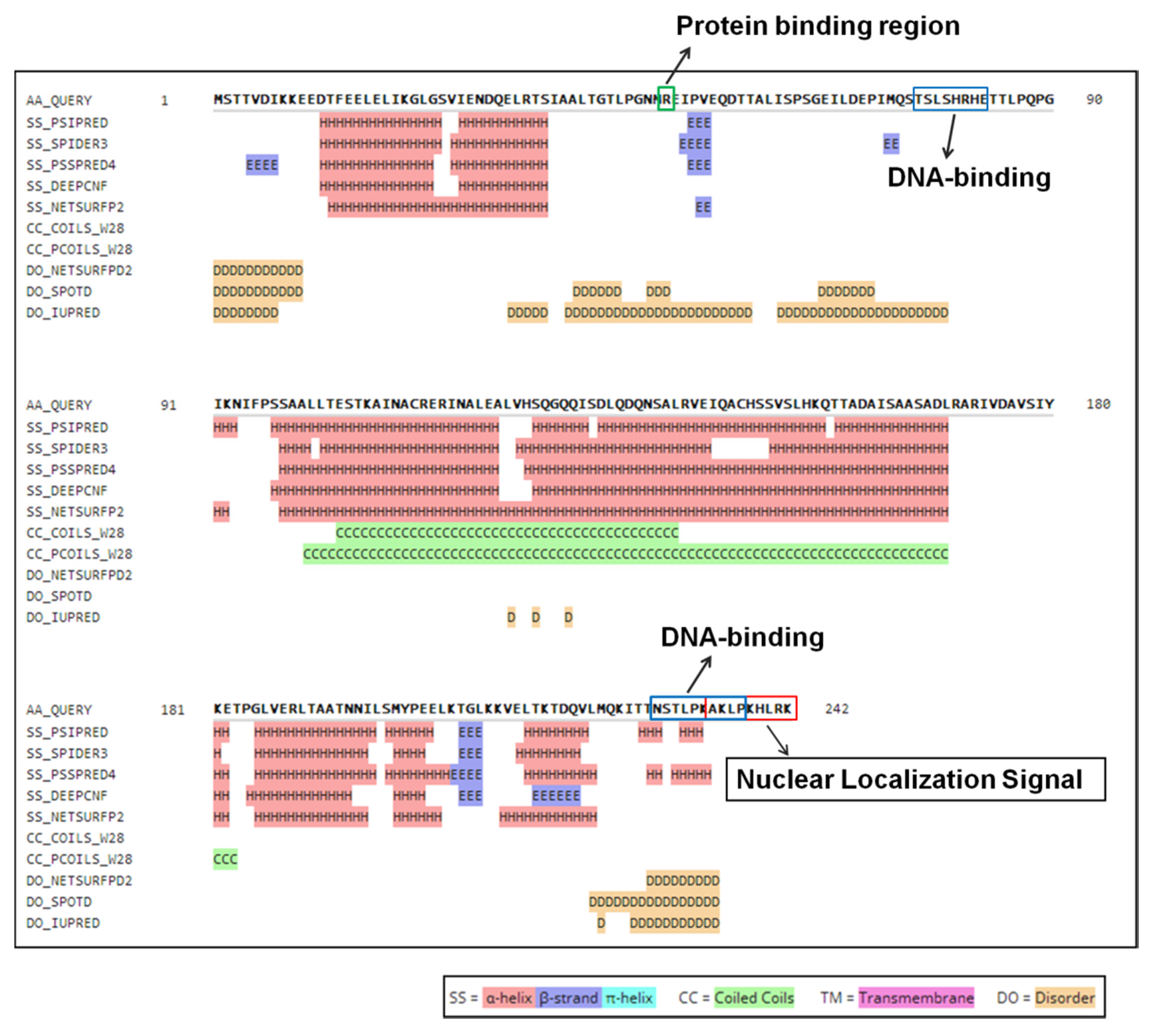
3.1. Predicted Secondary Structure and Function of Protein ORF I
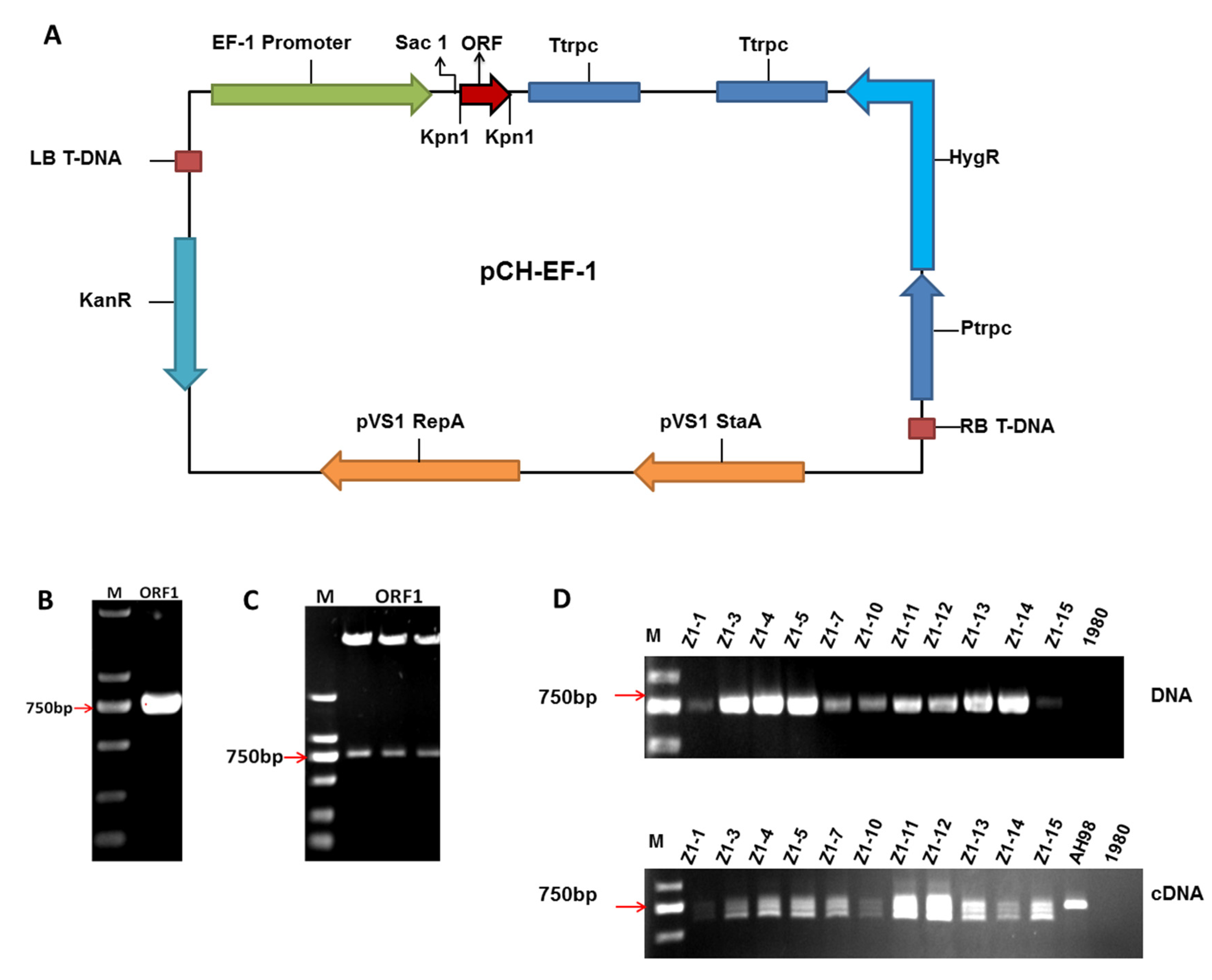
3.2. Construction of Expression Vector and Verification of Mutant Strains
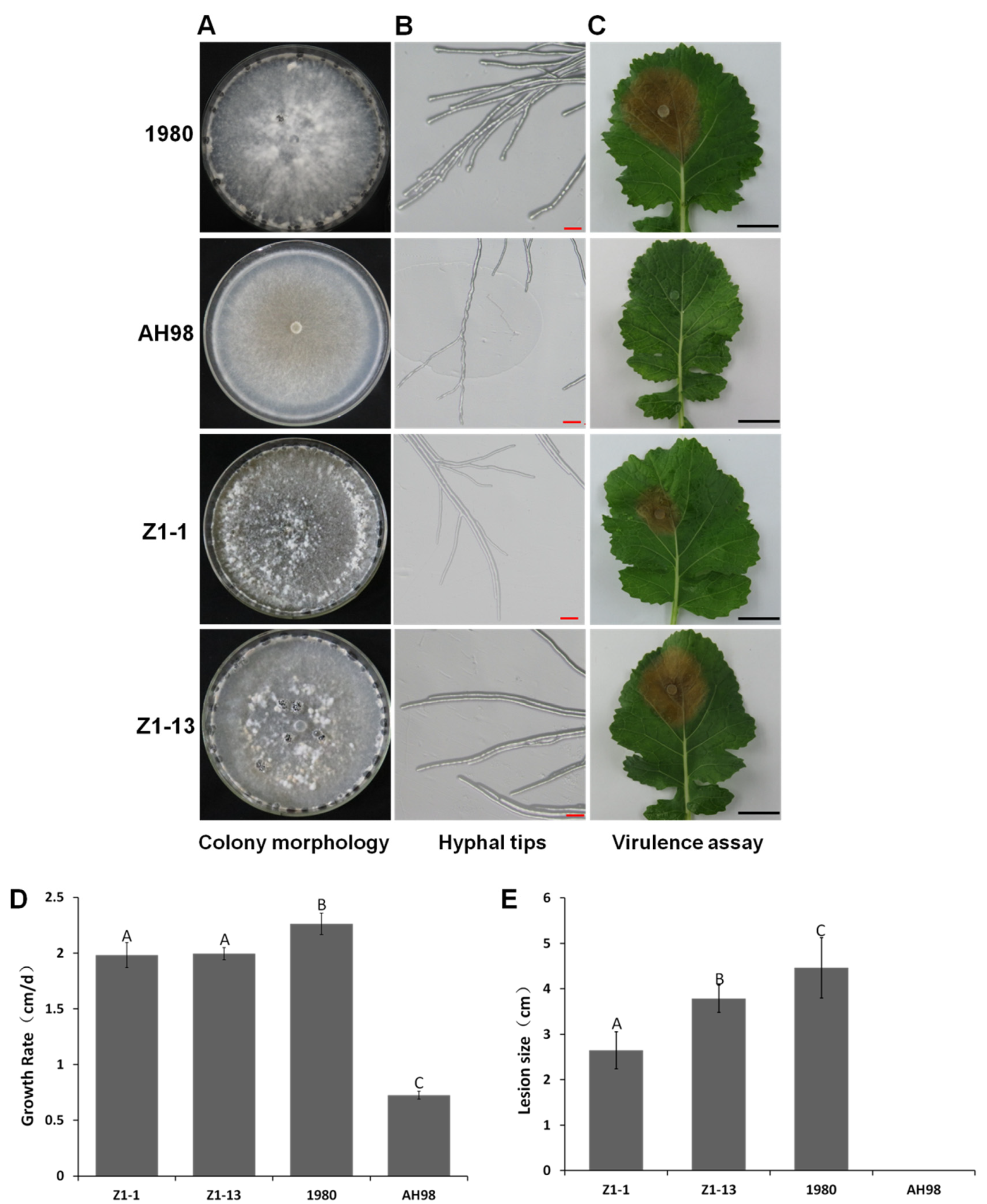
3.3. Biological Characterization of the ORF I -Expressing Strains
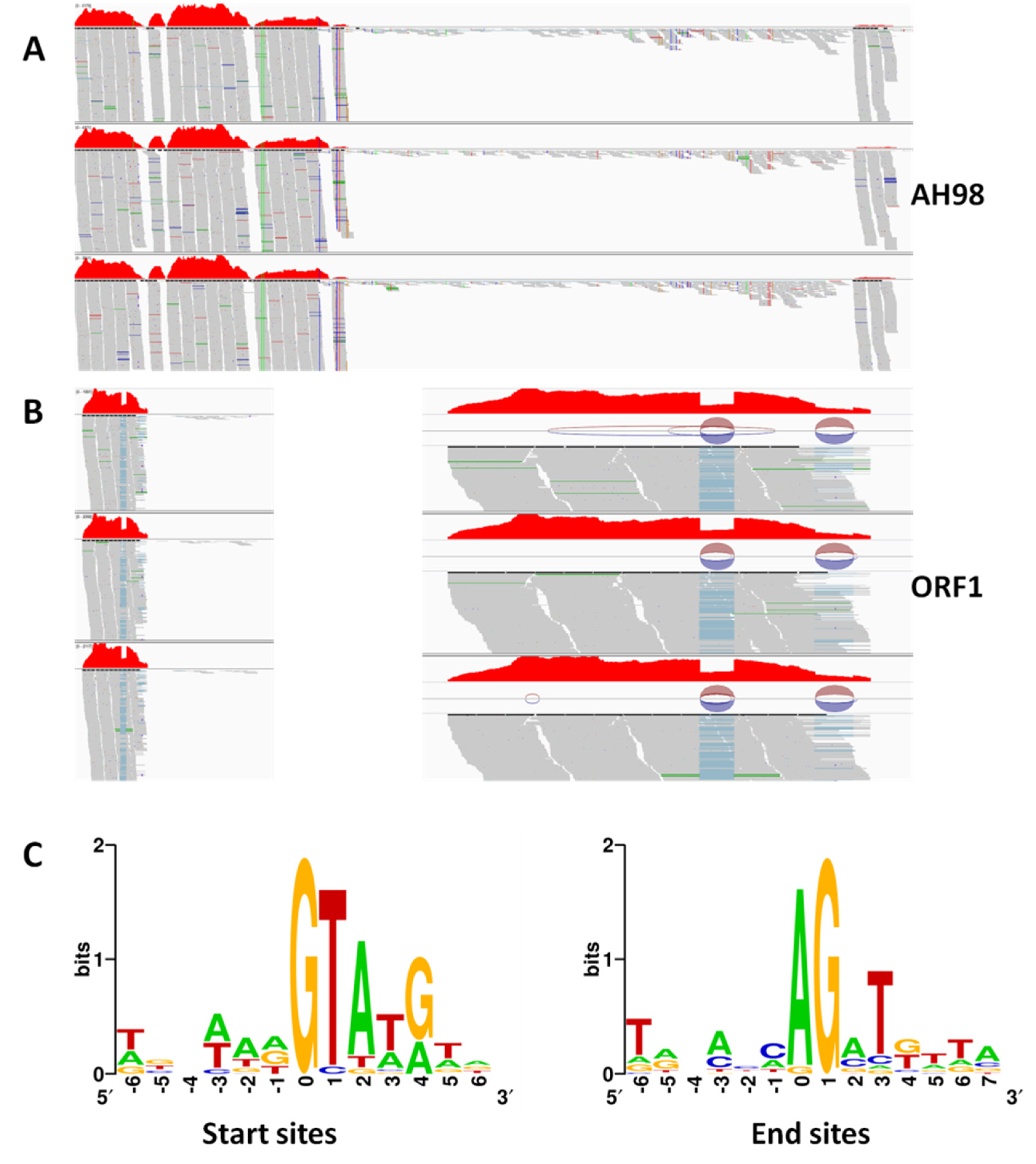
3.4. Transcriptions of Viral ORFs
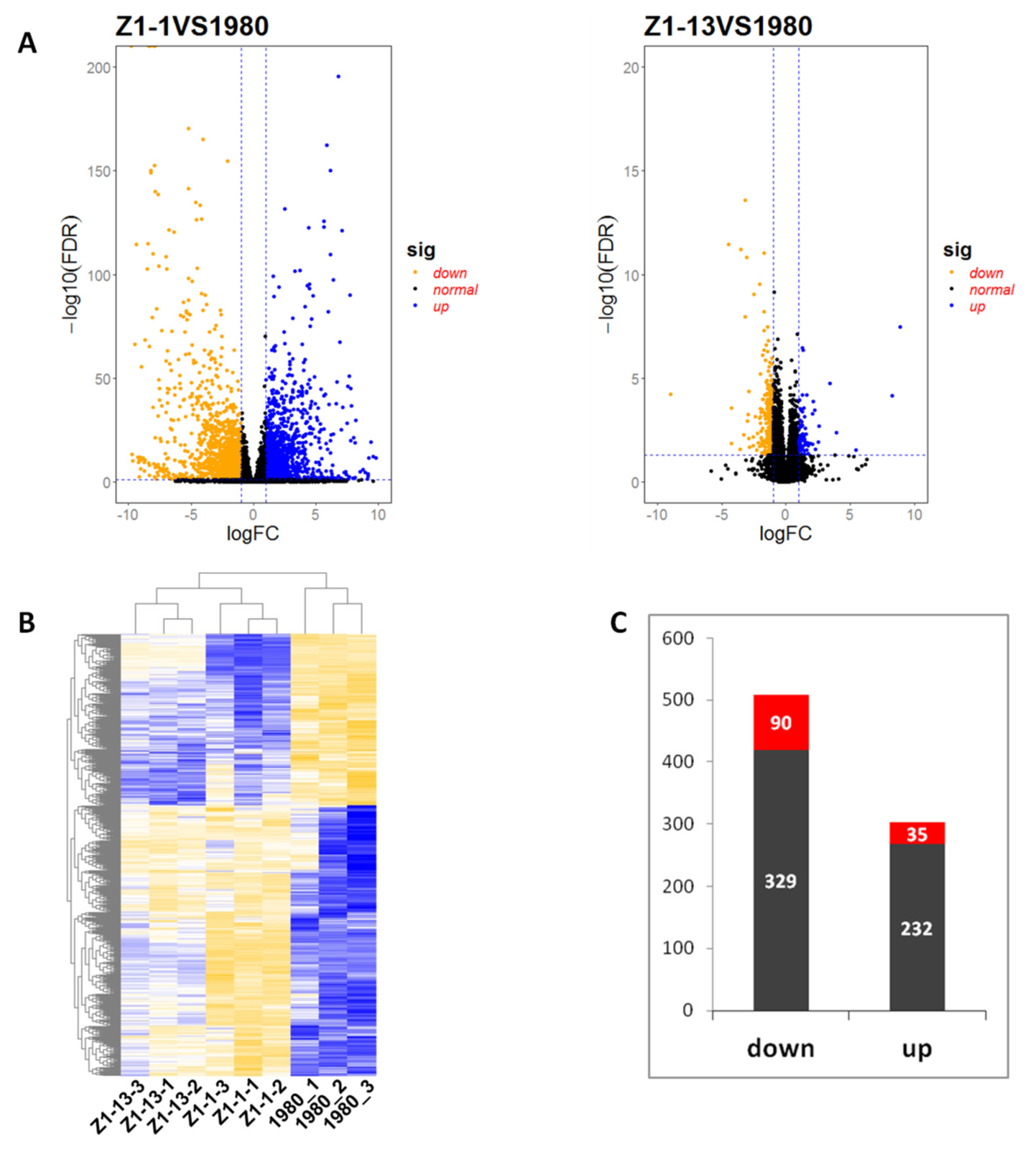
3.5. Overview and Validation of Differentially Expressed Genes (DEGs)
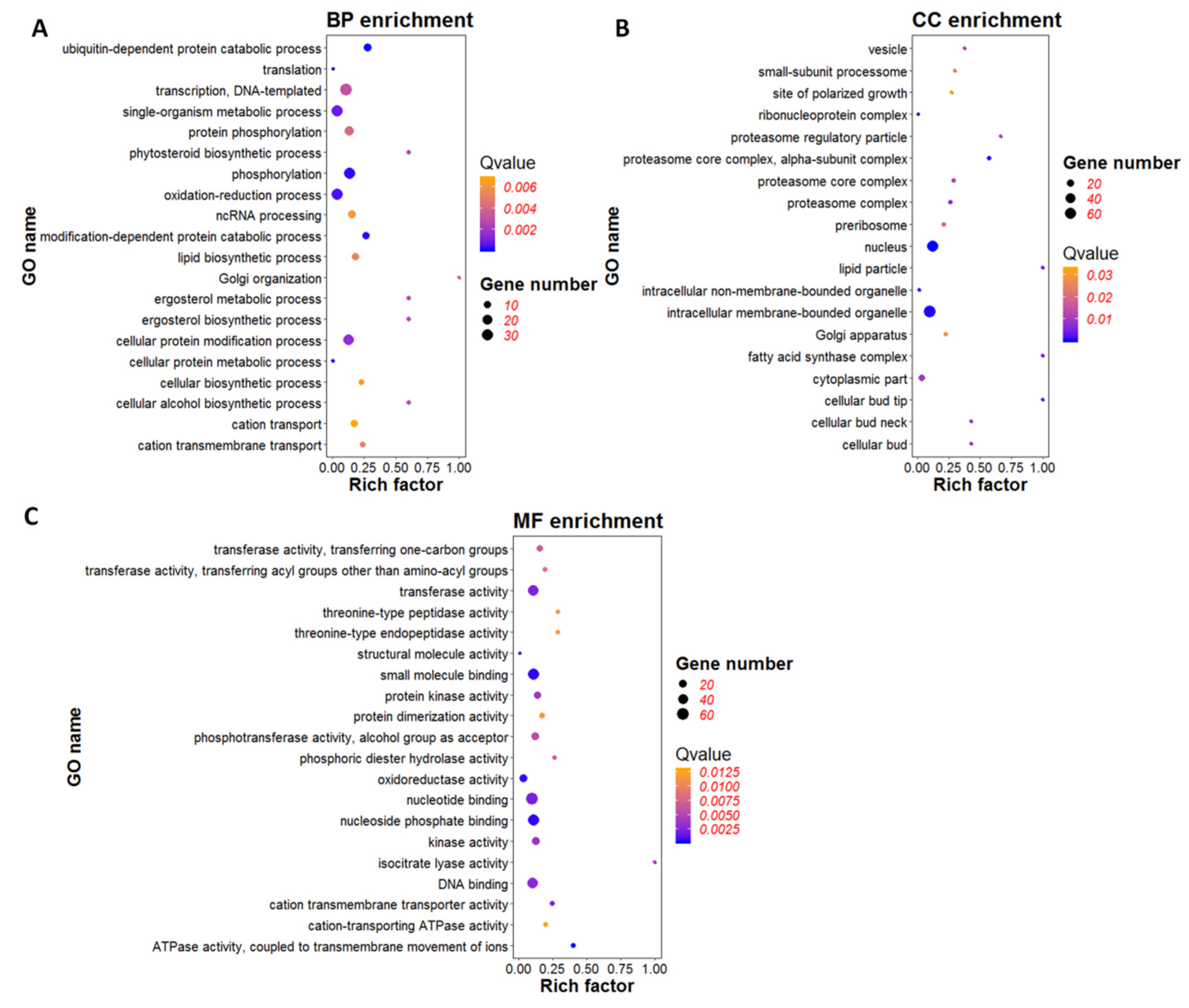
3.6. GO Enrichment Analysis of DEGs
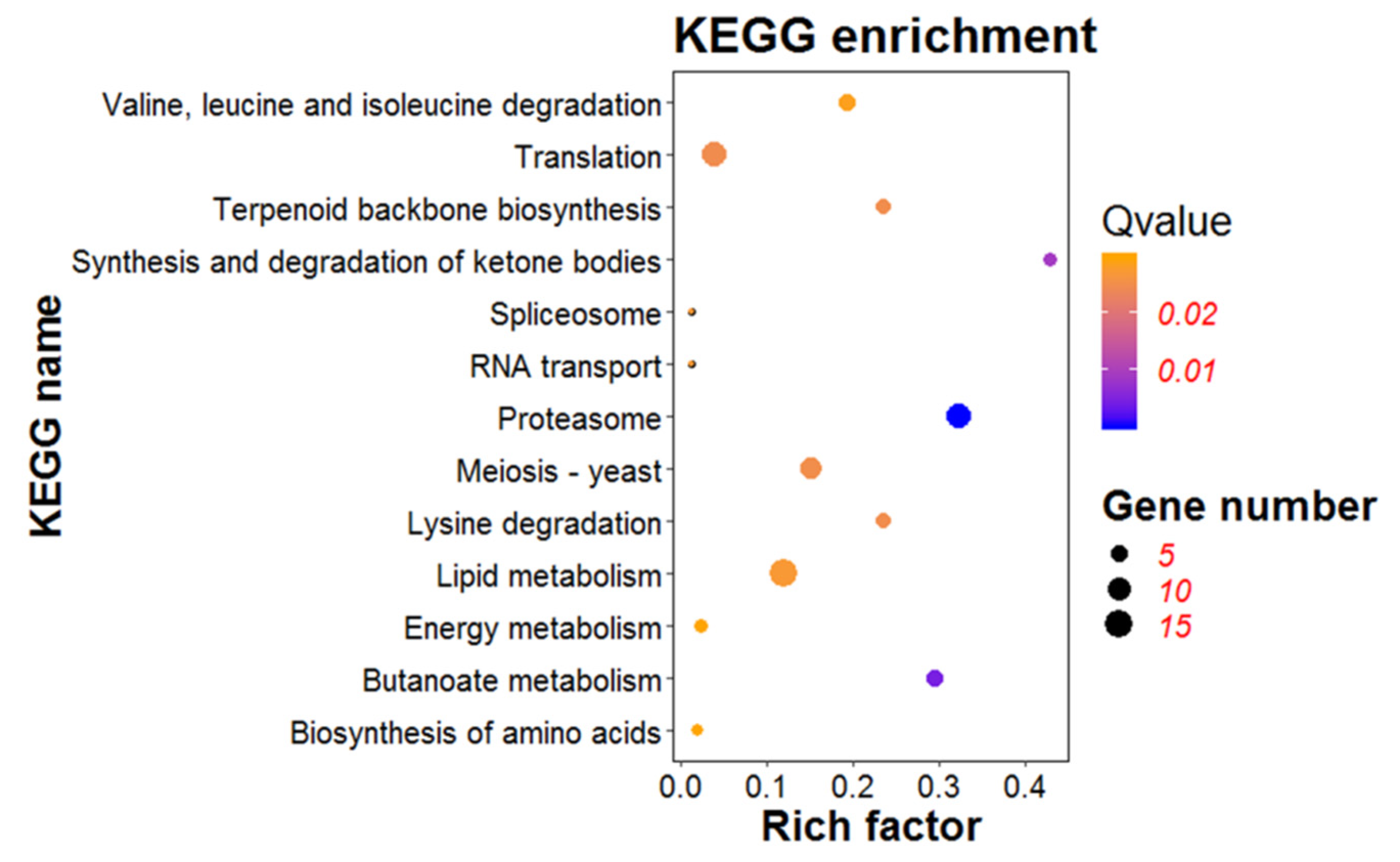
3.7. KEGG Pathway Analysis of DEGs
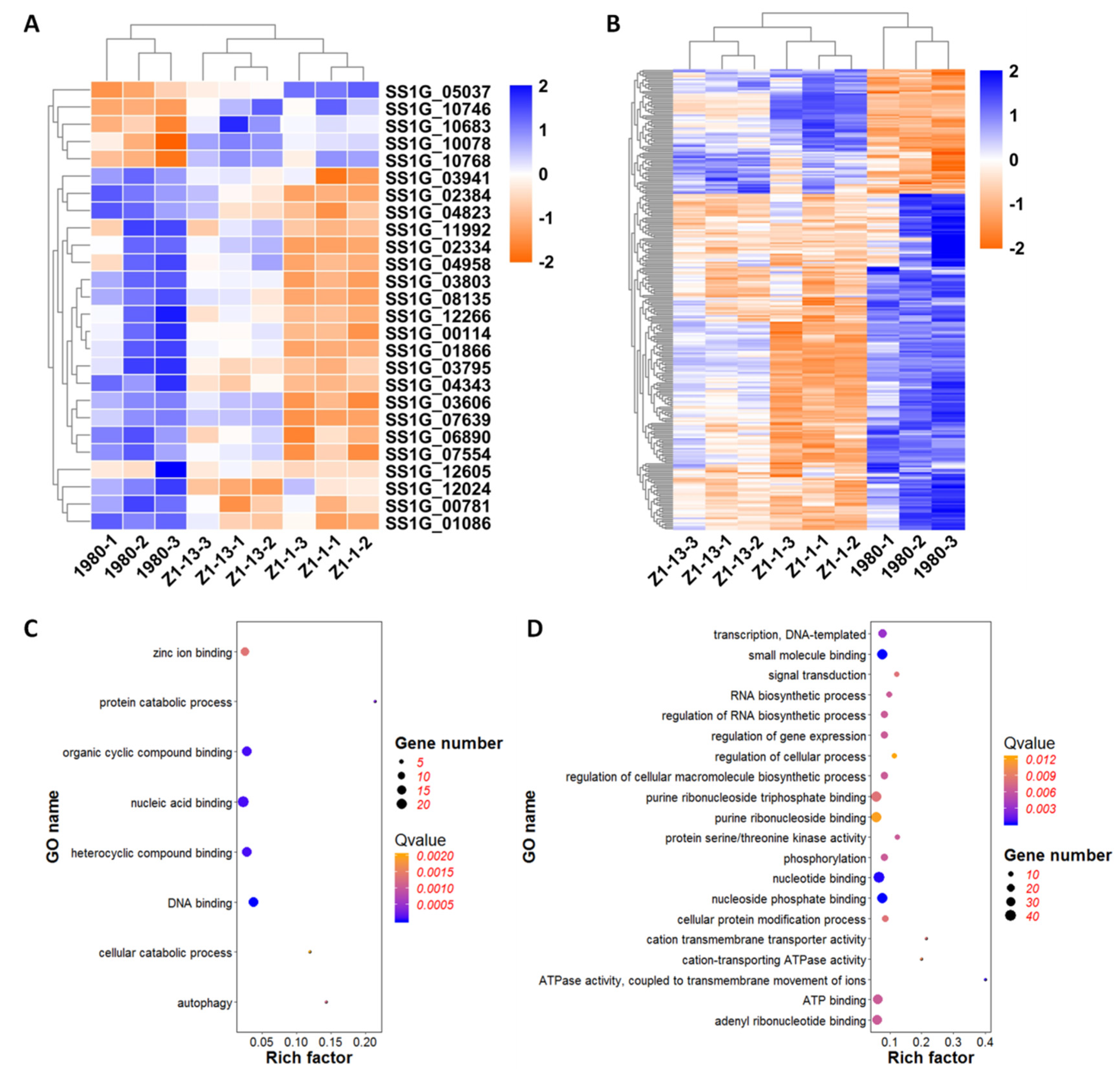
3.8. Heat Map Enrichment Analysis of DEGs Associated with Secretory Proteins and Pathogenesis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ghabrial, S.A.; Caston, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef]
- Anagnostakis, S.L. Biological Control of Chestnut Blight. Science 1982, 215, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.X.; Spiering, M.J.; Nuss, D.L. Characterizing the roles of Cryphonectria parasitica RNA-dependent RNA polymerase-like genes in antiviral defense, viral recombination and transposon transcript accumulation. PLoS ONE 2014, 9, e108653. [Google Scholar] [CrossRef] [Green Version]
- Kanematsu, S.; Shimizu, T.; Salaipeth, L.; Yaegashi, H.; Sasaki, A.; Ito, T.; Suzuki, N. Genome rearrangement of a mycovirus Rosellinia necatrix megabirnavirus 1 affecting its ability to attenuate virulence of the host fungus. Virology 2014, 450–451, 308–315. [Google Scholar] [CrossRef]
- Yu, X.; Li, B.; Fu, Y.; Jiang, D.; Ghabrial, S.A.; Li, G.; Peng, Y.; Xie, J.; Cheng, J.; Huang, J.; et al. A geminivirus-related DNA mycovirus that confers hypovirulence to a plant pathogenic fungus. Proc. Natl. Acad. Sci. USA 2010, 107, 8387–8392. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Xie, J.; Cheng, J.; Fu, Y.; Li, G.; Yi, X.; Jiang, D. Fungal negative-stranded RNA virus that is related to bornaviruses and nyaviruses. Proc. Natl. Acad. Sci. USA 2014, 111, 12205–12210. [Google Scholar] [CrossRef] [Green Version]
- Derbyshire, M.; Denton-Giles, M.; Hegedus, D.; Seifbarghy, S.; Rollins, J.; van Kan, J.; Seidl, M.F.; Faino, L.; Mbengue, M.; Navaud, O.; et al. The complete genome sequence of the phytopathogenic fungus Sclerotinia sclerotiorum reveals insights into the genome architecture of broad host range pathogens. Genome Biol. Evol. 2017, 9, 593–618. [Google Scholar] [CrossRef]
- Jiang, D.; Fu, Y.; Guoqing, L.; Ghabrial, S.A. Viruses of the plant pathogenic fungus Sclerotinia sclerotiorum. Adv. Virus Res. 2013, 86, 215–248. [Google Scholar]
- Khalifa, M.E.; Pearson, M.N. Molecular characterisation of an endornavirus infecting the phytopathogen Sclerotinia sclerotiorum. Virus Res. 2014, 189, 303–309. [Google Scholar] [CrossRef]
- Hamid, M.R.; Xie, J.; Wu, S.; Maria, S.K.; Zheng, D.; Assane Hamidou, A.; Wang, Q.; Cheng, J.; Fu, Y.; Jiang, D. A Novel Deltaflexivirus that Infects the Plant Fungal Pathogen, Sclerotinia sclerotiorum, Can Be Transmitted Among Host Vegetative Incompatible Strains. Viruses 2018, 10, 295. [Google Scholar] [CrossRef] [Green Version]
- Mu, F.; Xie, J.; Cheng, S.; You, M.P.; Barbetti, M.J.; Jia, J.; Wang, Q.; Cheng, J.; Fu, Y.; Chen, T.; et al. Virome Characterization of a Collection of S. sclerotiorum from Australia. Front. Microbiol. 2017, 8, 2540. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Zheng, D.; Cheng, J.; Chen, T.; Fu, Y.; Jiang, D.; Xie, J. Characterization of a novel Sclerotinia sclerotiorum RNA virus as the prototype of a new proposed family within the order Tymovirales. Virus Res. 2016, 219, 92–99. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Q.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Molecular characterization of a bipartite double-stranded RNA virus and its satellite-like RNA co-infecting the phytopathogenic fungus Sclerotinia sclerotiorum. Front. Microbiol. 2015, 6, 406. [Google Scholar] [CrossRef] [Green Version]
- Kotta-Loizou, I. Mycoviruses: Past, Present, and Future. Viruses 2019, 11, 361. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, C.K.; Cattaneo, R.; Schnell, M.J. Reverse genetics of Mononegavirales: How they work, new vaccines, and new cancer therapeutics. Virology 2015, 479–480, 331–344. [Google Scholar] [CrossRef] [Green Version]
- Ortin, J.; Martin-Benito, J. The RNA synthesis machinery of negative-stranded RNA viruses. Virology 2015, 479–480, 532–544. [Google Scholar] [CrossRef] [Green Version]
- Irigoyen, S.; Bedre, R.H.; Scholthof, K.B.; Mandadi, K.K. Genomic Approaches to Analyze Alternative Splicing, A Key Regulator of Transcriptome and Proteome Diversity in Brachypodium distachyon. Methods Mol. Biol. 2018, 1667, 73–85. [Google Scholar]
- Blencowe, B.J. Alternative splicing: New insights from global analyses. Cell 2006, 126, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Kelemen, O.; Convertini, P.; Zhang, Z.; Wen, Y.; Shen, M.; Falaleeva, M.; Stamm, S. Function of alternative splicing. Gene 2013, 514, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Graveley, B.R. Alternative Splicing: Increasing Diversity in the Proteomic World. Trends Genet. 2001, 17, 100–107. [Google Scholar] [CrossRef]
- Stamm, S.; Ben-Ari, S.; Rafalska, I.; Tang, Y.; Zhang, Z.; Toiber, D.; Thanaraj, T.A.; Soreq, H. Function of alternative splicing. Gene 2005, 344, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Boudreault, S.; Roy, P.; Lemay, G.; Bisaillon, M. Viral modulation of cellular RNA alternative splicing: A new key player in virus-host interactions? Wiley Interdiscip. Rev. RNA 2019, 10, e1543. [Google Scholar] [CrossRef] [Green Version]
- Mueller, N.; van Bel, N.; Berkhout, B.; Das, A.T. HIV-1 splicing at the major splice donor site is restricted by RNA structure. Virology 2014, 468–470, 609–620. [Google Scholar] [CrossRef]
- Chua, M.A.; Schmid, S.; Perez, J.T.; Langlois, R.A.; tenOever, B.R. Influenza A Virus Utilizes Suboptimal Splicing to Coordinate the Timing of Infection. Cell Rep. 2013, 3, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Kuwata, R.; Isawa, H.; Hoshino, K.; Tsuda, Y.; Yanase, T.; Sasaki, T.; Kobayashi, M.; Sawabe, K. RNA splicing in a new rhabdovirus from Culex mosquitoes. J. Virol. 2011, 85, 6185–6196. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, U.; Benoit-Pilven, C.; Lacroix, V.; Navratil, V.; Naffakh, N. Advances in Analyzing Virus-Induced Alterations of Host Cell Splicing. Trends Microbiol. 2019, 27, 268–281. [Google Scholar] [CrossRef]
- Bouton, C.; Geldreich, A.; Ramel, L.; Ryabova, L.A.; Dimitrova, M.; Keller, M. Cauliflower mosaic virus Transcriptome Reveals a Complex Alternative Splicing Pattern. PLoS ONE 2015, 10, e0132665. [Google Scholar] [CrossRef]
- Mandadi, K.K.; Scholthof, K.B. Genome-wide analysis of alternative splicing landscapes modulated during plant-virus interactions in Brachypodium distachyon. Plant Cell 2015, 27, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Wei, D.; Jiang, D.; Fu, Y.; Li, G.; Ghabrial, S.; Peng, Y. Characterization of debilitation-associated mycovirus infecting the plant-pathogenic fungus Sclerotinia sclerotiorum. J. Gen. Virol. 2006, 87, 241–249. [Google Scholar] [CrossRef] [PubMed]
- The Gene Ontology Consortium. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, L.; Stephens, A.; Nam, S.-Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.; Pant, R.; Raghunath, A.; Irvine, A.G.; Pedro, H.; Hammond-Kosack, K.E. The Pathogen-Host Interactions database (PHI-base): Additions and future developments. Nucleic Acids Res. 2015, 43, D645–D655. [Google Scholar] [CrossRef] [Green Version]
- Guyon, K.; Balagué, C.; Roby, D.; Raffaele, S. Secretome analysis reveals effector candidates associated with broad host range necrotrophy in the fungal plant pathogen Sclerotinia sclerotiorum. BMC Genom. 2014, 15, 336. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lubbers, R.J.M.; Simon, A.; Stassen, J.H.M.; Kan, J.A.L.V. A novel Zn2Cys6 transcription factor BcGaaR regulates D-galacturonic acid utilization in Botrytis cinerea. Mol. Microbiol. 2015, 100, 247–262. [Google Scholar] [CrossRef]
- Arakawa, G.Y.; Kudo, H.; Yanase, A.; Eguchi, Y.; Kodama, H.; Ogawa, M.; Koyama, Y.; Shindo, H.; Hosaka, M.; Tokuoka, M. A unique Zn(II)2-Cys6-type protein, KpeA, is involved in secondary metabolism and conidiation in Aspergillus oryzae. Fungal Genet. Biol. 2019, 127, 35–44. [Google Scholar] [CrossRef]
- Rybak, K.; See, P.T.; Phan, H.T.; Syme, R.A.; Moffat, C.S.; Oliver, R.P.; Tan, K.C. A functionally conserved Zn2Cys6 binuclear cluster transcription factor class regulates necrotrophic effector gene expression and host-specific virulence of two major Pleosporales fungal pathogens of wheat. Mol. Plant Pathol. 2017, 18, 420–434. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.W.S.; Simpson, C.G. Splice site selection in plant pre-mRNA splicing. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 77–95. [Google Scholar] [CrossRef]
- Twyffels, L.; Gueydan, C.; Kruys, V. Shuttling SR proteins: More than splicing factors. FEBS J. 2011, 278, 3246–3255. [Google Scholar] [CrossRef] [PubMed]
- Fica, S.M.; Nagai, K. Cryo-electron microscopy snapshots of the spliceosome: Structural insights into a dynamic ribonucleoprotein machine. Nat. Struct. Mol. Biol. 2017, 24, 791–799. [Google Scholar] [CrossRef]
- De, M.F.A.; Guillermo, R.; Iglesias, N.G.; Priya, S.; Berta, P.; Gebhard, L.G.; Pablo, M.; Estefania, M.; Yanovsky, M.J.; Raul, A.; et al. The Dengue Virus NS5 Protein Intrudes in the Cellular Spliceosome and Modulates Splicing. PLoS Pathog. 2016, 12, e1005841. [Google Scholar]
- Papasaikas, P.; Valcarcel, J. The Spliceosome: The Ultimate RNA Chaperone and Sculptor. Trends Biochem. Sci. 2016, 41, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Tomonaga, K.; Kobayashi, T.; Lee, B.J.; Watanabe, M.; Kamitani, W.; Ikuta, K. Identification of alternative splicing and negative splicing activity of a nonsegmented negative-strand RNA virus, Borna disease virus. Proc. Natl. Acad. Sci. USA 2000, 97, 12788–12793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.; Etingov, I.; Pante, N. Effect of viral infection on the nuclear envelope and nuclear pore complex. Int. Rev. Cell Mol. Biol. 2012, 299, 117–159. [Google Scholar]
- Avota, E.; Harms, H.; Schneider-Schaulies, S. Measles virus induces expression of SIP110, a constitutively membrane clustered lipid phosphatase, which inhibits T cell proliferation. Cell. Microbiol 2006, 8, 1826–1839. [Google Scholar] [CrossRef]
- Conzelmann, K.K. Reverse Genetics of Mononegavirales. In Biology of Negative Strand RNA Viruses: The Power of Reverse Genetics; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2004; Volume 283, pp. 1–41. [Google Scholar]
- Green, T.J.; Zhang, X.; Wertz, G.W.; Luo, M. Structure of the vesicular stomatitis virus nucleoprotein-RNA complex. Science 2006, 313, 357–360. [Google Scholar] [CrossRef]
- Lou, Z. A Structural View of Negative-Sense RNA Virus Nucleocapsid Protein and Its Functions Beyond. In Virus Protein and Nucleoprotein Complexes; Harris, J.R., Bhella, D., Eds.; Springer: Singapore, 2018; pp. 19–37. [Google Scholar]
- Takai, Y.; Sasaki, T.; Matozaki, T. Small GTP-binding proteins. Physiol. Rev. 2001, 81, 153–208. [Google Scholar] [CrossRef]
- Sodhi, A.; Montaner, S.; Patel, V.; Zohar, M.; Bais, C.; Mesri, E.A.; Gutkind, J.S. The Kaposi’s Sarcoma-associated Herpes Virus G Protein-coupled Receptor Up-Regulates Vascular Endothelial Growth Factor Expression and Secretion through Mitogen-activated Protein Kinase and p38 Pathways Acting on Hypoxia-inducible Factor 1α. Cancer Res. 2000, 60, 4873–4880. [Google Scholar]
- Zhen, Z.; Bradel-Tretheway, B.; Sumagin, S.; Bidlack, J.M.; Dewhurst, S. The human herpesvirus 6 G protein-coupled receptor homolog U51 positively regulates virus replication and enhances cell-cell fusion in vitro. J. Virol. 2005, 79, 11914–11924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenya, E.; Trusov, Y.; Dietzgen, R.G.; Botella, J.R. Heterotrimeric G-proteins facilitate resistance to plant pathogenic viruses in Arabidopsis thaliana (L.) Heynh. Plant. Signal. Behav. 2016, 11, e1212798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J. The effect of Sclerotinia sclerotiorum negative-stranded RNA virus 1 (SsNSRV-1) genes on its host. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2017; pp. 27–35. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hit Name | Function | Probability | E-Value |
|---|---|---|---|
| 5CWS_D | Nucleoporin; nucleocytoplasmic transport, PROTEIN TRANSPORT | 82.52 | 8.3 |
| 3TNU_B | Keratin, type I cytoskeletal 14; Coiled-coil, Structural Support | 71.8 | 22 |
| 4EGW_A | Magnesium transport protein CorA; magnesium transporter, magnesium binding | 68.46 | 48 |
| 5IJO_S | Nuclear pore complex protein Nup155; Nuclear pore complex, Nucleocytoplasmic transport | 66.44 | 23 |
| 1NLW_E | MAD PROTEIN/MAX PROTEIN/DNA; transcription factor | 64.78 | 8.7 |
| 1NLW_A | MAD PROTEIN/MAX PROTEIN/DNA; transcription factor | 64.72 | 11 |
| 4ZRY_A | Keratin, type I cytoskeletal 10; keratin, intermediate filament, coiled-coil | 63.03 | 50 |
| 3NVO_A | Zinc transport protein zntB | 61.26 | 90 |
| 5N77_C | Magnesium transport protein CorA; Homopentamer Complex Transport Membrane, transport | 60.49 | 87 |
| 6GAO_B | Outer capsid protein sigma-1; cell attachment protein, reovirus sigma1 | 57.97 | 74 |
| 2O3E_A | Neurolysin, thermolysin-like domain, substrate-binding channel | 52.51 | 110 |
| 4RSI_B | Structural maintenance of chromosomes protein; Smc hinge domain with coiled | 52.12 | 140 |
| 6G1L_A | Microphthalmia-associated transcription factor/DNA Complex; melanocyte, autophagy, transcription factor | 51.9 | 40 |
| 5IJO_H | Nuclear pore complex protein Nup155, Nucleocytoplasmic transport | 50.41 | 120 |
| 6GAP_A | Outer capsid protein sigma-1; cell attachment protein, reovirus sigma1 | 50.13 | 93 |
| 4ATH_A | DNA binding protein, transcription factor | 50.13 | 96 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Z.; Wu, J.; Jiang, D.; Xie, J.; Cheng, J.; Lin, Y. ORF Ι of Mycovirus SsNSRV-1 is Associated with Debilitating Symptoms of Sclerotinia sclerotiorum. Viruses 2020, 12, 456. https://doi.org/10.3390/v12040456
Gao Z, Wu J, Jiang D, Xie J, Cheng J, Lin Y. ORF Ι of Mycovirus SsNSRV-1 is Associated with Debilitating Symptoms of Sclerotinia sclerotiorum. Viruses. 2020; 12(4):456. https://doi.org/10.3390/v12040456
Chicago/Turabian StyleGao, Zhixiao, Junyan Wu, Daohong Jiang, Jiatao Xie, Jiasen Cheng, and Yang Lin. 2020. "ORF Ι of Mycovirus SsNSRV-1 is Associated with Debilitating Symptoms of Sclerotinia sclerotiorum" Viruses 12, no. 4: 456. https://doi.org/10.3390/v12040456
APA StyleGao, Z., Wu, J., Jiang, D., Xie, J., Cheng, J., & Lin, Y. (2020). ORF Ι of Mycovirus SsNSRV-1 is Associated with Debilitating Symptoms of Sclerotinia sclerotiorum. Viruses, 12(4), 456. https://doi.org/10.3390/v12040456