Broadly Neutralizing Bovine Antibodies: Highly Effective New Tools against Evasive Pathogens?



Abstract
:1. Introduction
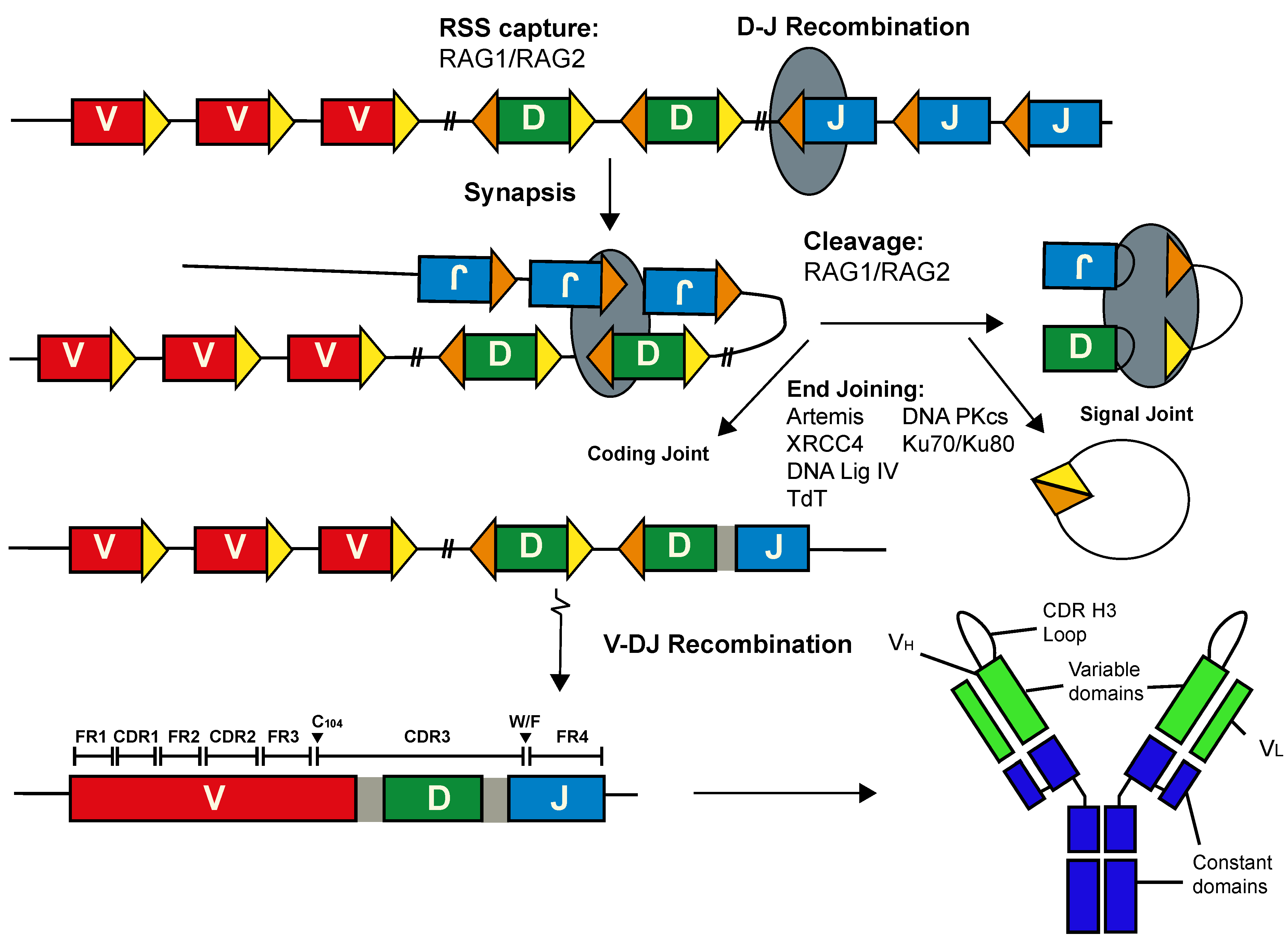
2. The Long CDR H3 Is Generated Inefficiently in Humans
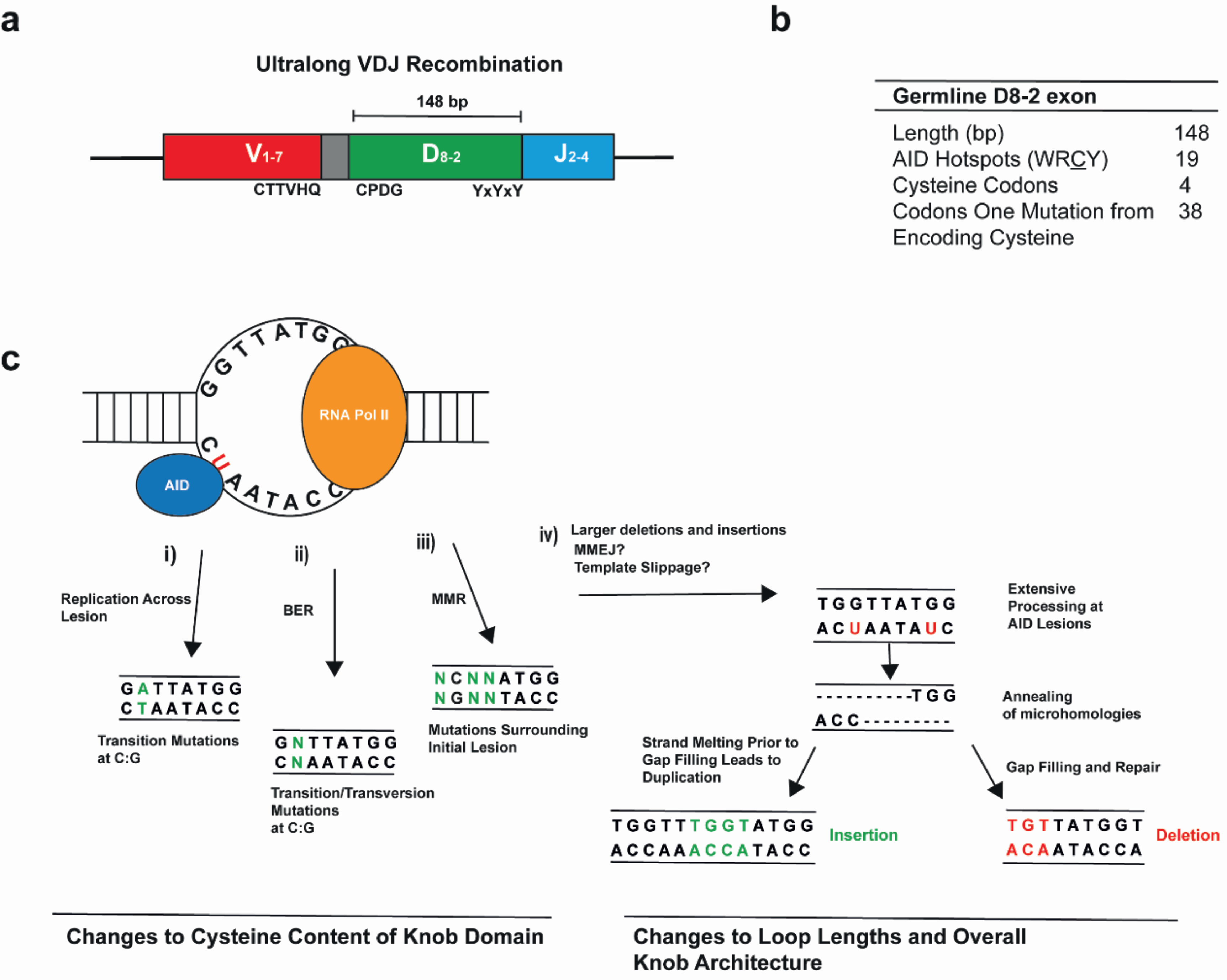
3. Ultralong CDR H3 Are Prevalent in the Cow Ab Repertoire
4. Diversification of the Ultralong CDR H3 in Cows
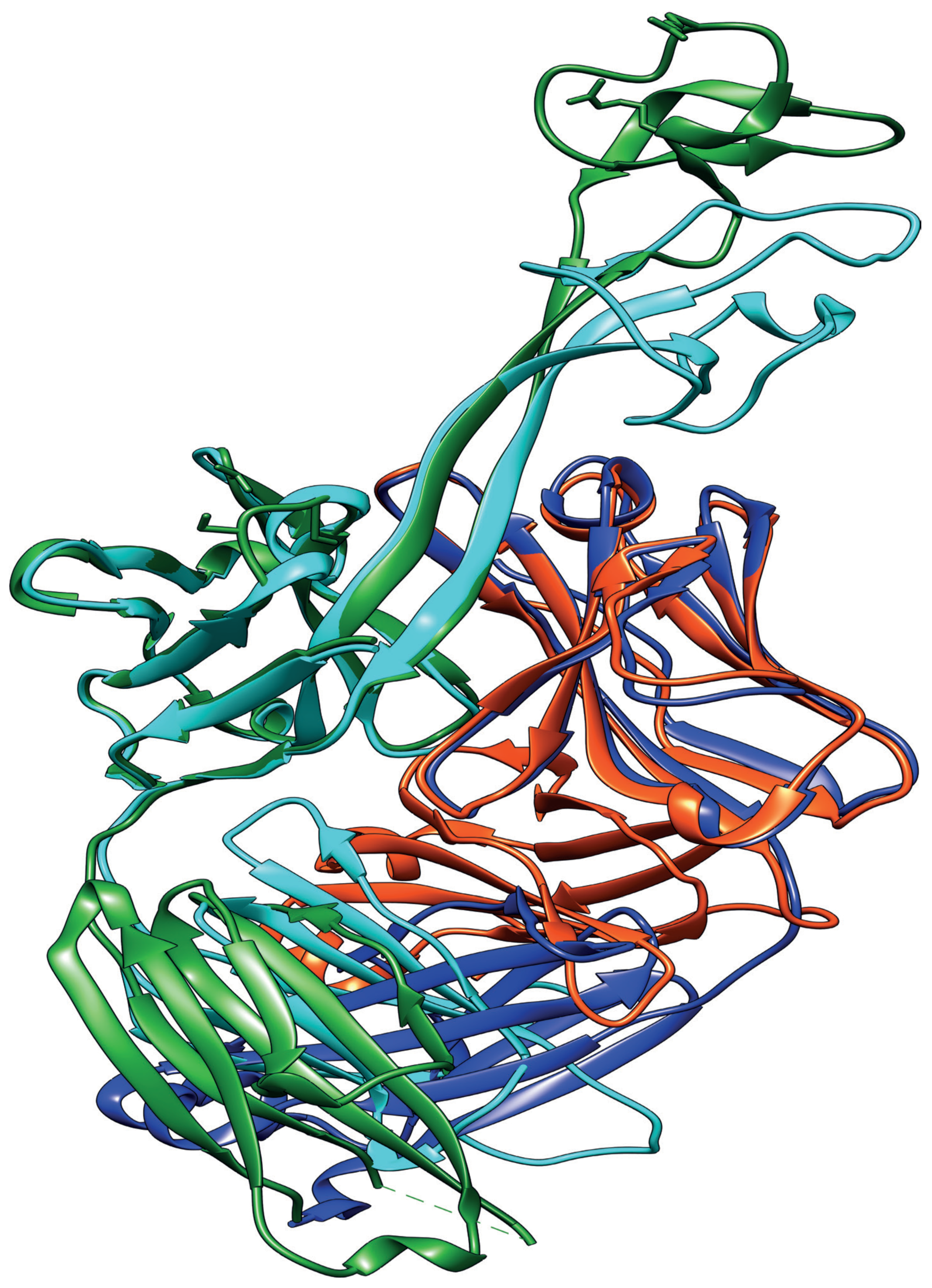
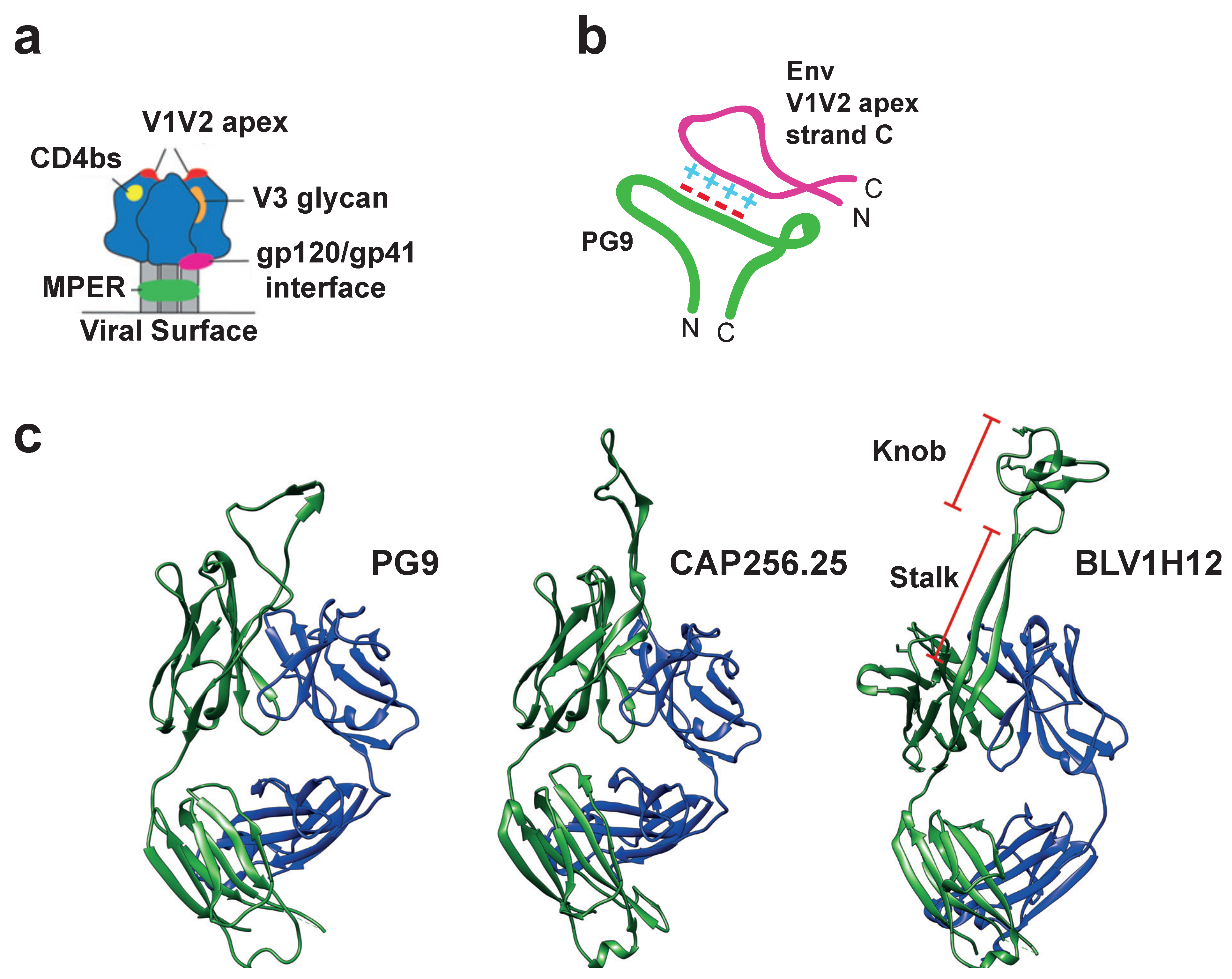
5. Uncovering the Structure of the Cow Ultralong CDR H3
6. Long CDR H3 Structures Confer Broadly Neutralizing Anti-Viral Properties
7. Summary and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Schroeder, H.W., Jr.; Cavacini, L. Structure and Function of Immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef] [Green Version]
- Zemlin, M.; Klinger, M.; Link, J.; Zemlin, C.; Bauer, K.; Engler, J.A.; Schroeder, H.W., Jr.; Kirkham, P.M. Expressed murine and human CDR-H3 intervals of equal length exhibit distinct repertoires that differ in their amino acid composition and predicted range of structures. J. Mol. Biol. 2003, 334, 733–749. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Mizuguchi, K. The diversity of H3 loops determines the antigen-binding tendencies of antibody CDR loops. Protein Sci. 2016, 25, 815–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadofsky, M.J. The RAG proteins in V(D)J recombination: More than just a nuclease. Nucleic Acids Res. 2001, 29, 1399–1409. [Google Scholar] [CrossRef] [PubMed]
- Jung, D.; Alt, F.W. Unraveling V (D) J Recombination: Insights into Gene Regulation V(D)J recombination assembles antigen receptor. Cell 2004, 116, 299–311. [Google Scholar] [CrossRef] [Green Version]
- Schatz, D.G.; Ji, Y. Recombination centres and the orchestration of V(D)J recombination. Nat. Rev. Immunol. 2011, 11, 251–263. [Google Scholar] [CrossRef]
- Bertocci, B.; De Smet, A.; Weill, J.C.; Reynaud, C.A. Nonoverlapping Functions of DNA Polymerases Mu, Lambda, and Terminal Deoxynucleotidyltransferase during Immunoglobulin V(D)J Recombination In Vivo. Immunity 2006, 25, 31–41. [Google Scholar] [CrossRef]
- Jackson, K.J.; Gaeta, B.; Sewell, W.; Collins, A.M. Exonuclease activity and P nucleotide addition in the generation of the expressed immunoglobulin repertoire. BMC Immunol. 2004, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Chandra, V.; Bortnick, A.; Murre, C. AID targeting: Old mysteries and new challenges. Trends Immunol. 2015, 36, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Meyer-Hermann, M.; George, L.A.; Figge, M.T.; Khan, M.; Goodall, M.; Young, S.P.; Reynolds, A.; Falciani, F.; Waisman, A.; et al. Germinal center B cells govern their own fate via antibody feedback. J. Exp. Med. 2013, 210, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Stewart, I.; Radtke, D.; Phillips, B.; McGowan, S.J.; Bannard, O. Germinal Center B Cells Replace Their Antigen Receptors in Dark Zones and Fail Light Zone Entry when Immunoglobulin Gene Mutations are Damaging. Immunity 2018, 49, 477–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liljavirta, J.; Niku, M.; Pessa-Morikawa, T.; Ekman, A.; Iivanainen, A. Expansion of the preimmune antibody repertoire by junctional diversity in Bos taurus. PLoS ONE 2014, 9, e99808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, S.D.; Wrin, T.; Smith, D.M.; Kosakovsky Pond, S.L.; Liu, Y.; Paxinos, E.; Chappey, C.; Galovich, J.; Beauchaine, J.; Petropoulos, C.J.; et al. Neutralizing antibody responses drive the evolution of human immunodeficiency virus type 1 envelope during recent HIV infection. Proc. Natl. Acad. Sci. USA 2005, 102, 18514–18519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilgrim, A.K.; Pantaleo, G.; Cohen, O.J.; Fink, L.M.; Zhou, J.Y.; Zhou, J.T.; Bolognesi, D.P.; Fauci, A.S.; Montefiori, D.C. Neutralizing Antibody Responses to Human Immunodeficiency Virus Type 1 in Primary Infection and Long-Term-Nonprogressive Infection. J. Infect. Dis. 1997, 176, 924–932. [Google Scholar] [CrossRef]
- Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.P.; Wang, S.K.; Ramos, A.; Chan-Hui, P.Y.; Moyle, M.; et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466–470. [Google Scholar] [CrossRef] [Green Version]
- Doria-Rose, N.A.; Schramm, C.A.; Gorman, J.; Moore, P.L.; Bhiman, J.N.; DeKosky, B.J.; Ernandes, M.J.; Georgiev, I.S.; Kim, H.J.; Pancera, M.; et al. Developmental pathway for potent V1V2-directed HIV-neutralizing antibodies. Nature 2014, 508, 55–62. [Google Scholar] [CrossRef]
- Yu, L.; Guan, Y. Immunologic basis for long HCDR3s in broadly neutralizing antibodies against HIV-1. Front. Immunol. 2014, 5, 250. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Ekiert, D.C.; Ahmad, I.; Yu, W.; Zhang, Y.; Bazirgan, O.; Torkamani, A.; Raudsepp, T.; Mwangi, W.; Criscitiello, M.F.; et al. Reshaping antibody diversity. Cell 2013, 153, 1379–1393. [Google Scholar] [CrossRef] [Green Version]
- Stanfield, R.L.; Haakenson, J.; Deiss, T.C.; Criscitiello, M.F.; Wilson, I.A.; Smider, V.V. The Unusual Genetics and Biochemistry of Bovine Immunoglobulins. Adv. Immunol. 2018, 137, 135–164. [Google Scholar] [CrossRef]
- Sok, D.; Le, K.M.; Vadnais, M.; Saye-Francisco, K.L.; Jardine, J.G.; Torres, J.L.; Berndsen, Z.T.; Kong, L.; Stanfield, R.; Ruiz, J.; et al. Rapid elicitation of broadly neutralizing antibodies to HIV by immunization in cows. Nature 2017, 548, 108–111. [Google Scholar] [CrossRef] [Green Version]
- Koch, K.; Kalusche, S.; Torres, J.L.; Stanfield, R.L.; Danquah, W.; Khazanehdari, K.; von Briesen, H.; Geertsma, E.R.; Wilson, I.A.; Wernery, U.; et al. Selection of Nanobodies with Broad Neutralizing Potential Against Primary HIV1 Strains Using Soluble Subtype C gp140 Envelope Trimers. Sci. Rep. 2017, 7, 8390–8405, s41598–s41617. [Google Scholar] [CrossRef]
- Muyldermans, S.; Smider, V.V. Distinct Antibody Species: Structural Differences Creating Therapeutic Opportunities. Curr. Opin. Immunol. 2016, 40, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Qin, T.; Chu, D.; Cheng, X.; Wang, J.; Wang, X.; Wang, P.; Han, H.; Ren, L.; Aitken, R.; et al. Internal Duplications of DH, JH, and C Region Genes Create an Unusual IgH Gene Locus in Cattle. J. Immunol. 2016, 196, 4358–4366. [Google Scholar] [CrossRef] [Green Version]
- Lefranc, M.P.; Pommié, C.; Ruiz, M.; Giudicelli, V.; Foulquier, E.; Truong, L.; Thouvenin-Contet, V.; Lefranc, G. IMGT unique numbering for immunoglobulin and T cell receptor variable domains and Ig superfamily V-like domains. Dev. Comp. Immunol. 2003, 27, 55–77. [Google Scholar] [CrossRef]
- Roskin, K.M.; Jackson, K.; Lee, J.Y.; Hoh, R.A.; Joshi, S.A.; Hwang, K.K.; Bonsignori, M.; Pedroza-Pacheco, I.; Liao, H.X.; Moody, M.A.; et al. Aberrant B cell repertoire selection associated with HIV neutralizing antibody breadth. Nat. Immunol. 2020, 21, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Mroczek, E.S.; Ippolito, G.C.; Rogosch, T.; Hoi, K.H.; Hwangpo, T.A.; Brand, M.G.; Zhuang, Y.; Liu, C.R.; Schneider, D.A.; Zemlin, M.; et al. Differences in the composition of the human antibody repertoire by b cell subsets in the blood. Front. Immunol. 2014, 5, 96. [Google Scholar] [CrossRef] [PubMed]
- Sliepen, K.; Medina-Ramírez, M.; Yasmeen, A.; Moore, J.P.; Klasse, P.J.; Sanders, R.W. Binding of inferred germline precursors of broadly neutralizing HIV-1 antibodies to native-like envelope trimers. Virology 2015, 486, 116–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, L.M.; Phogat, S.K.; Chan-Hui, P.Y.; Wagner, D.; Phung, P.; Goss, J.L.; Wrin, T.; Simek, M.D.; Fling, S.; Mitcham, J.L.; et al. Broad and potent neutralizing antibodies from an african donor reveal a new HIV-1 vaccine target. Science 2009, 326, 285–289. [Google Scholar] [CrossRef] [Green Version]
- McLellan, J.S.; Pancera, M.; Carrico, C.; Gorman, J.; Julien, J.P.; Khayat, R.; Louder, R.; Pejchal, R.; Sastry, M.; Dai, K.; et al. Structure of HIV-1 gp120 V1/V2 domain with broadly neutralizing antibody PG9. Nature 2011, 480, 336–343. [Google Scholar] [CrossRef]
- Ekiert, D.C.; Kashyap, A.K.; Steel, J.; Rubrum, A.; Bhabha, G.; Khayat, R.; Lee, J.H.; Dillon, M.A.; O’Neil, R.E.; Faynboym, A.M.; et al. Cross-neutralization of influenza A viruses mediated by a single antibody loop. Nature 2012, 489, 526–532. [Google Scholar] [CrossRef]
- Bowers, P.M.; Verdino, P.; Wang, Z.; da Silva Correia, J.; Chhoa, M.; Macondray, G.; Do, M.; Neben, T.Y.; Horlick, R.A.; Stanfield, R.L.; et al. Nucleotide insertions and deletions complement point mutations to massively expand the diversity created by somatic hypermutation of antibodies. J. Biol. Chem. 2014, 289, 33557–33567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flyak, A.I.; Ruiz, S.E.; Salas, J.; Rho, S.; Bailey, J.R.; Bjorkman, P.J. An ultralong CDRH2 in HCV neutralizing antibody demonstrates structural plasticity of antibodies against E2 glycoprotein. eLife 2020, 9, e53169. [Google Scholar] [CrossRef]
- Walker, L.M.; Burton, D.R. Passive immunotherapy of viral infections: ‘super-antibodies’ enter the fray. Nat. Rev. Immunol. 2018, 18, 297–308. [Google Scholar] [CrossRef]
- Caskey, M.; Klein, F.; Nussenzweig, M.C. Broadly neutralizing anti-HIV-1 monoclonal antibodies in the clinic. Nat. Med. 2019, 25, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Escolano, A.; Steichen, J.M.; Dosenovic, P.; Kulp, D.W.; Golijanin, J.; Sok, D.; Freund, N.T.; Gitlin, A.D.; Oliveira, T.; Araki, T.; et al. Sequential Immunization Elicits Broadly Neutralizing Anti-HIV-1 Antibodies in Ig Knockin Mice. Cell 2016, 166, 1445–1458.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, W.B.; Zhang, J.; Jiang, C.; Nicely, N.I.; Fera, D.; Luo, K.; Moody, M.A.; Liao, H.X.; Alam, S.M.; Kepler, T.B.; et al. Initiation of HIV neutralizing B cell lineages with sequential envelope immunizations. Nat. Commun. 2017, 8, 1732–1751. [Google Scholar] [CrossRef]
- Deiss, T.C.; Vadnais, M.; Wang, F.; Chen, P.L.; Torkamani, A.; Mwangi, W.; Lefranc, M.P.; Criscitiello, M.F.; Smider, V.V. Immunogenetic factors driving formation of ultralong VH CDR3 in Bos taurus antibodies. Cell. Mol. Immunol. 2017, 14, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Yeap, L.S.; Hwang, J.K.; Du, Z.; Meyers, R.M.; Meng, F.L.; Jakubauskaitė, A.; Liu, M.; Mani, V.; Neuberg, D.; Kepler, T.B.; et al. Sequence-Intrinsic Mechanisms that Target AID Mutational Outcomes on Antibody Genes. Cell 2015, 163, 1124–1137. [Google Scholar] [CrossRef] [Green Version]
- Saini, S.S.; Allore, B.; Jacobs, R.M.; Kaushik, A. Exceptionally long CDR3H region with multiple cysteine residues in functional bovine IgM antibodies. Eur. J. Immunol. 1999, 29, 2420–2426. [Google Scholar] [CrossRef]
- Saini, S.S.; Kaushik, A. Extensive CDR3H length heterogeneity exists in bovine foetal VDJ rearrangements. Scand. J. Immunol. 2002, 55, 140–148. [Google Scholar] [CrossRef]
- Walther, S.; Czerny, C.P.; Diesterbeck, U.S. Exceptionally Long CDR3H Are Not Isotype Restricted in Bovine Immunoglobulins. PLoS ONE 2013, 8, e64234. [Google Scholar] [CrossRef] [PubMed]
- Jack, H.; Wu, G.; Martin, D.; Bradl, H.; Collins, T.; Roth, E. Selection of Ig μ Heavy Chains by Composition Complementarity-Determining Regions 3 Length and Amino Acid Composition. J. Immunol. 2003, 171, 4663–4671. [Google Scholar] [CrossRef]
- Wu, T.; Johnson, G.; Kabat, E. Length Distribution of CDR H3 in Antibodies. Proteins Struct. Funct. Bioinform. 1993, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Haakenson, J.K.; Huang, R.; Smider, V.V. Diversity in the cow ultralong CDR H3 antibody repertoire. Front. Immunol. 2018, 9, 1262. [Google Scholar] [CrossRef] [PubMed]
- Ramiro, A.R.; Jankovic, M.; Eisenreich, T.; Difilippantonio, S.; Chen-Kiang, S.; Muramatsu, M.; Honjo, T.; Nussenzweig, A.; Nussenzweig, M.C. AID is required for the cMyc/IgH chromosome translocation in vivo. Cell 2004, 118, 431–438. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Finn, J.A.; Larsen, P.A.; Smith, T.; Crowe, J.E., Jr. Structural Diversity of Ultralong CDRH3s in Seven Bovine Antibody Heavy Chains. Front. Immunol. 2019, 10, 558. [Google Scholar] [CrossRef]
- Stanfield, R.L.; Wilson, I.A.; Smider, V.V. Conservation and diversity in the ultralong third heavy-chain complementarity-determining region of bovine antibodies. Sci. Immunol. 2016, 1, aaf7962. [Google Scholar] [CrossRef] [Green Version]
- Vadnais, M.L.; Smider, V.V. Bos taurus ultralong CDR H3 antibodies. Curr. Opin. Struct. Biol. 2016, 38, 62–67. [Google Scholar] [CrossRef]
- Sippl, M.J.; Wiederstein, M. Detection of Spatial Correlations in Protein Structures and Molecular Complexes. Structure 2012, 20, 718–728. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Andrabi, R.; Su, C.Y.; Yasmeen, A.; Julien, J.P.; Kong, L.; Wu, N.C.; McBride, R.; Sok, D.; Pauthner, M.; et al. A Broadly Neutralizing Antibody Targets the Dynamic HIV Envelope Trimer Apex via a Long, Rigidified, and Anionic β-Hairpin Structure. Immunity 2017, 46, 690–702. [Google Scholar] [CrossRef] [Green Version]
- Sok, D.; Burton, D.R. Recent progress in broadly neutralizing antibodies to HIV. Nat. Immunol. 2018, 19, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Lazniewski, M.; Dawson, W.K.; Szczepinska, T.; Plewczynski, D. The structural variability of the influenza A hemagglutinin receptor-binding site. Brief. Funct. Genom. 2018, 17, 415–427. [Google Scholar] [CrossRef]
- Hellert, J.; Buchrieser, J.; Larrous, F.; Minola, A.; de Melo, G.D.; Soriaga, L.; England, P.; Haouz, A.; Telenti, A.; Schwartz, O.; et al. Structure of the prefusion-locking broadly neutralizing antibody RVC20 bound to the rabies virus glycoprotein. Nat. Commun. 2020, 11, 596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, R.W.; Derking, R.; Cupo, A.; Julien, J.P.; Yasmeen, A.; de Val, N.; Kim, H.J.; Blattner, C.; de la Peña, A.T.; Korzun, J.; et al. A Next-Generation Cleaved, Soluble HIV-1 Env Trimer, BG505 SOSIP.664 gp140, Expresses Multiple Epitopes for Broadly Neutralizing but Not Non-Neutralizing Antibodies. PLoS Pathog. 2013, 9, e1003618. [Google Scholar] [CrossRef] [Green Version]
- Pauthner, M.; Havenar-Daughton, C.; Sok, D.; Nkolola, J.P.; Bastidas, R.; Boopathy, A.V.; Carnathan, D.G.; Chandrashekar, A.; Cirelli, K.M.; Cottrell, C.A.; et al. Elicitation of Robust Tier 2 Neutralizing Antibody Responses in Nonhuman Primates by HIV Envelope Trimer Immunization Using Optimized Approaches. Immunity 2017, 46, 1073–1088.e6. [Google Scholar] [CrossRef] [Green Version]
- Ng, W.; Wong, V.; Muller, B.; Rawlin, G.; Brown, L. Prevention and Treatment of Influenza with Hyperimmune Bovine Colostrum Antibody. PLoS ONE 2010, 5, e13622. [Google Scholar] [CrossRef] [Green Version]
- Devilder, M.C.; Moyon, M.; Gautreau-Rolland, L.; Navet, B.; Perroteau, J.; Delbos, F.; Gesnel, M.C.; Breathnach, R.; Saulquin, X. Ex vivo evolution of human antibodies by CRISPR-X: From a naive B cell repertoire to affinity matured antibodies. BMC Biotechnol. 2019, 19, 14. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Liu, Y.; Wang, Y.; Hull, M.; Schultz, P.G.; Wang, F. Rational design of CXCR4 specific antibodies with elongated CDRs. J. Am. Chem. Soc. 2014, 136, 10557–10560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koti, M.; Saini, S.S.; Sachan, A.; Kaushik, A.K. Engineered Bovine Antibodies in the Development of Novel Therapeutics, Immunomodulators and Vaccines. Antibodies 2014, 3, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Pieper, K.; Tan, J.; Piccoli, L.; Foglierini, M.; Barbieri, S.; Chen, Y.; Silacci-Fregni, C.; Wolf, T.; Jarrossay, D.; Anderle, M.; et al. Public Antibodies to Malaria Antigens Generated by Two LAIR1 Insertion Modalities. Nature 2017, 548, 597–601. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Qureshi, H.; Deshpande, S.; Bhattacharya, J. Broadly neutralizing antibodies in HIV-1 treatment and prevention. Ther. Adv. Vaccines Immunother. 2018, 6, 61–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Advantage | Possible Drawbacks |
|---|---|---|
| Ultralong CDR H3 stalk and knob structure |
|
|
| Knob domain is highly stable |
|
|
| IGHD8-2 gene encoding the knob domain is primed for diversification |
|
|
| All other CDRs are invariable |
|
|
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burke, M.J.; Stockley, P.G.; Boyes, J. Broadly Neutralizing Bovine Antibodies: Highly Effective New Tools against Evasive Pathogens? Viruses 2020, 12, 473. https://doi.org/10.3390/v12040473
Burke MJ, Stockley PG, Boyes J. Broadly Neutralizing Bovine Antibodies: Highly Effective New Tools against Evasive Pathogens? Viruses. 2020; 12(4):473. https://doi.org/10.3390/v12040473
Chicago/Turabian StyleBurke, Matthew J., Peter G. Stockley, and Joan Boyes. 2020. "Broadly Neutralizing Bovine Antibodies: Highly Effective New Tools against Evasive Pathogens?" Viruses 12, no. 4: 473. https://doi.org/10.3390/v12040473
APA StyleBurke, M. J., Stockley, P. G., & Boyes, J. (2020). Broadly Neutralizing Bovine Antibodies: Highly Effective New Tools against Evasive Pathogens? Viruses, 12(4), 473. https://doi.org/10.3390/v12040473