Determination of Protein Interactions among Replication Components of Apple Necrotic Mosaic Virus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Plasmid Construction and Genetic Transformation
2.3. RNA Extraction and Complete Genomic RNA Sequence Acquisition
2.4. Homology Comparison and Phylogenetic Analysis
2.5. Yeast-Two-Hybrid Assay
2.6. Pull-Down Analysis
2.7. Bimolecular Fluorescent Complementation (BiFC) Assay
2.8. Luciferase Complementation Imaging Assay
3. Results
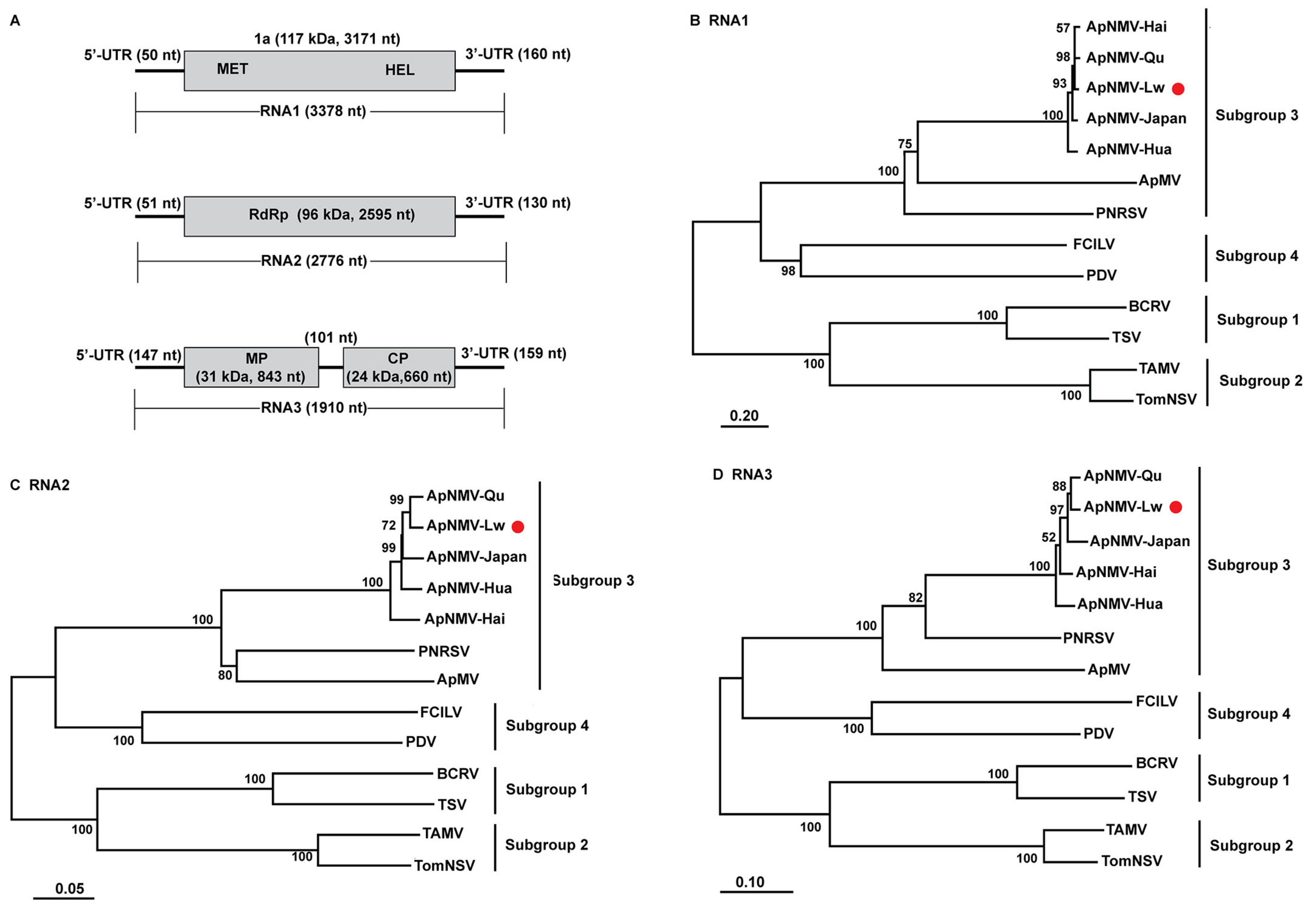
3.1. Complete Nucleotide Sequence and Phylogenetic Analysis of ApNMV–Lw
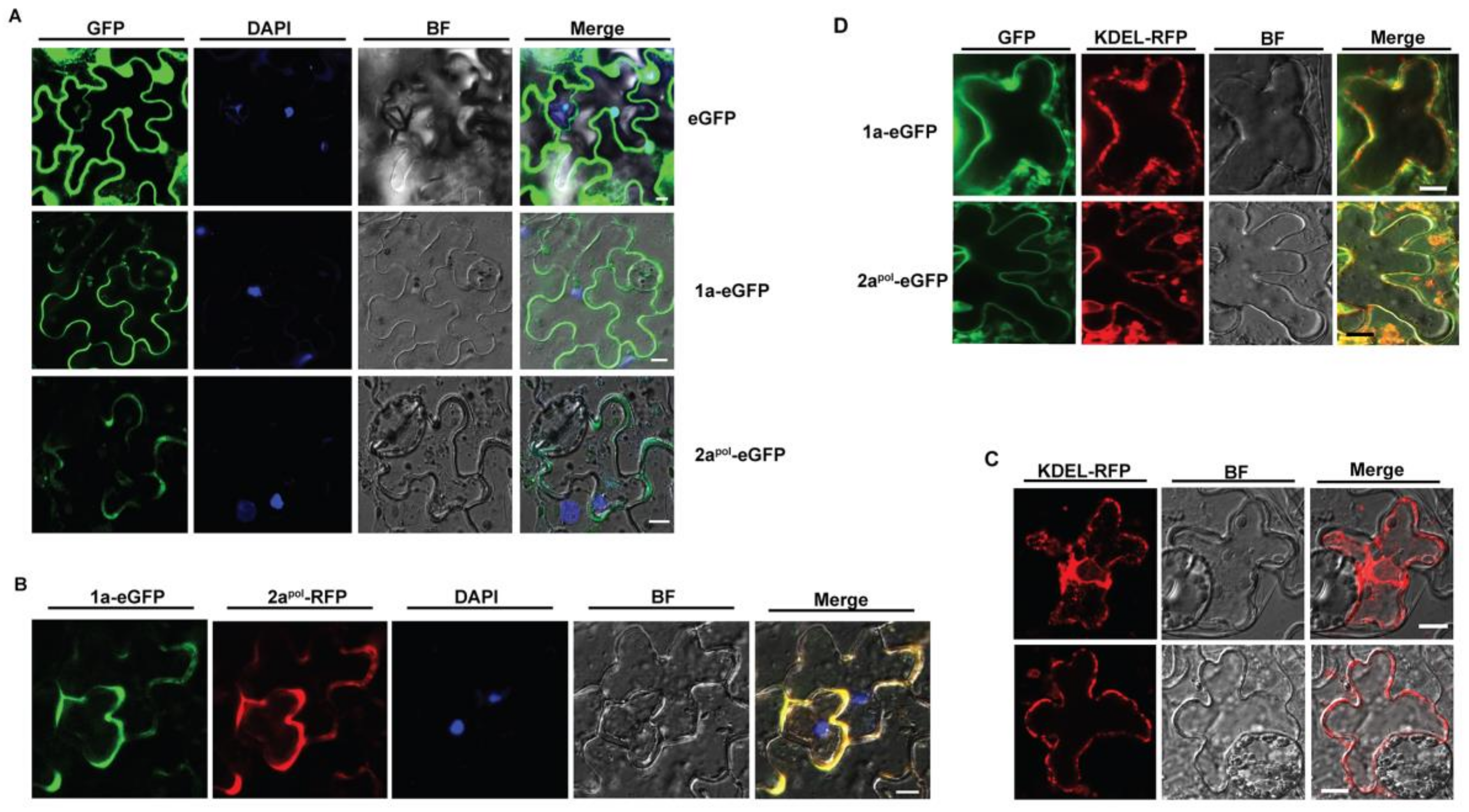
3.2. ApNMV 1a Colocalizes with 2apol in the Cytoplasm
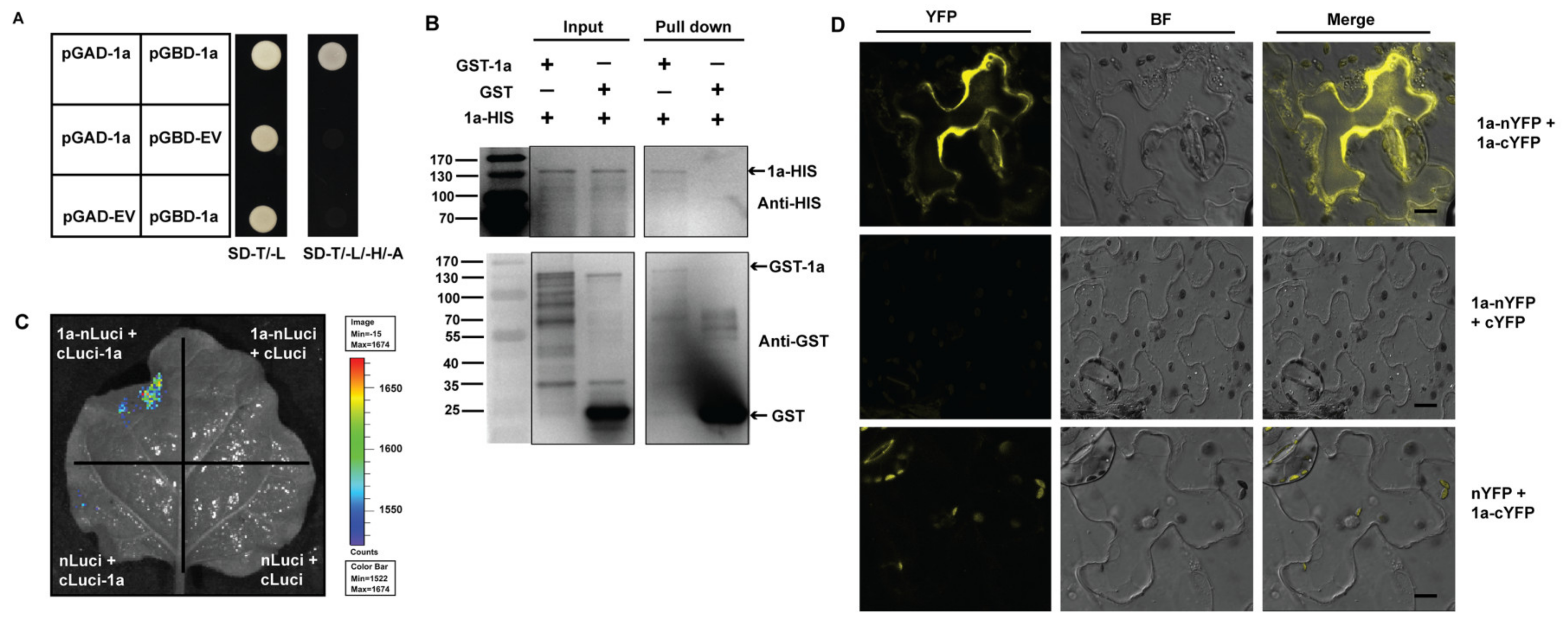
3.3. Inter-Molecular Interactions of ApNMV 1a with Itself
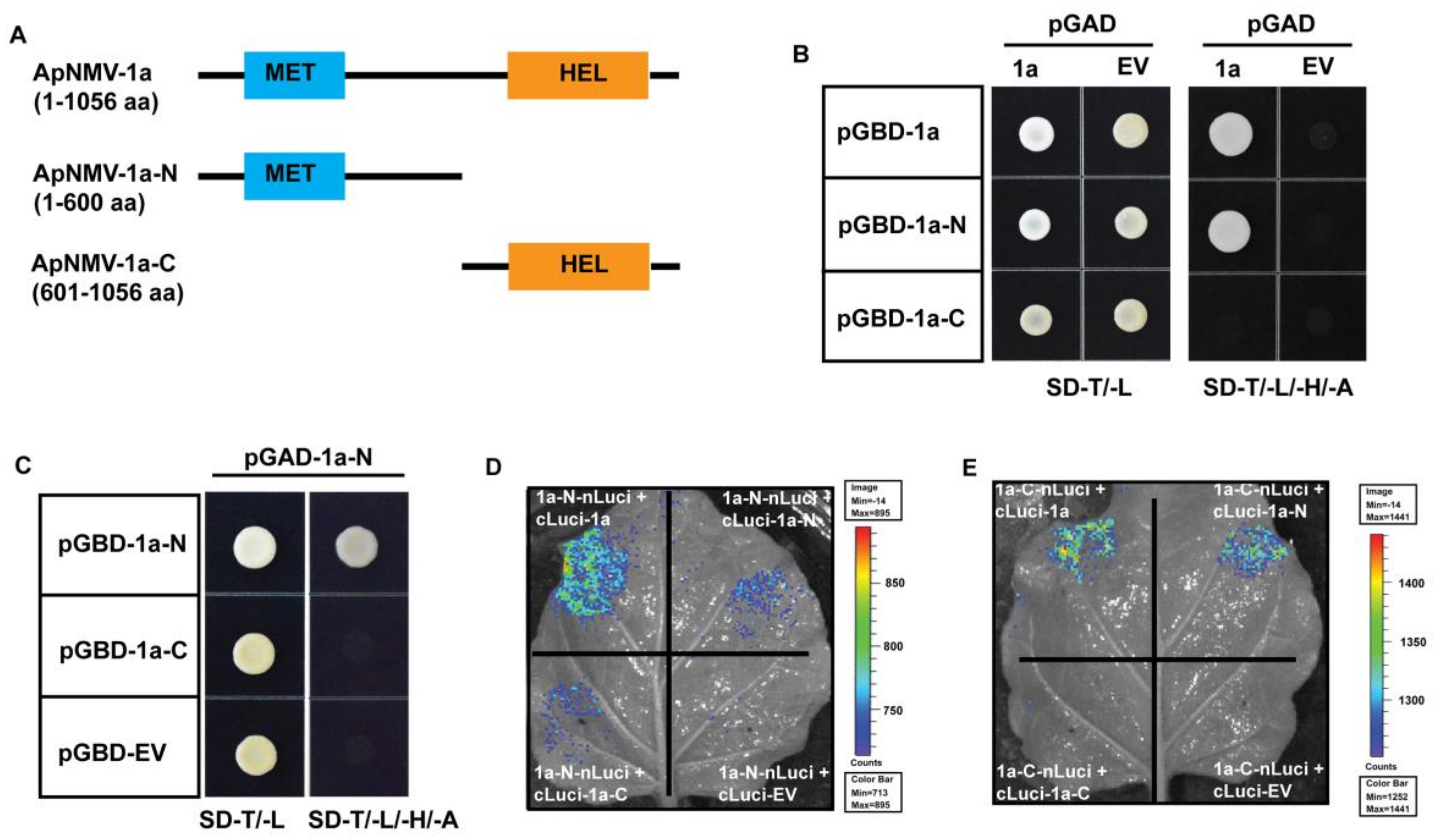
3.4. The N-Terminal of 1a Plays a Key Role in Its Inter- and Intramolecular Interactions
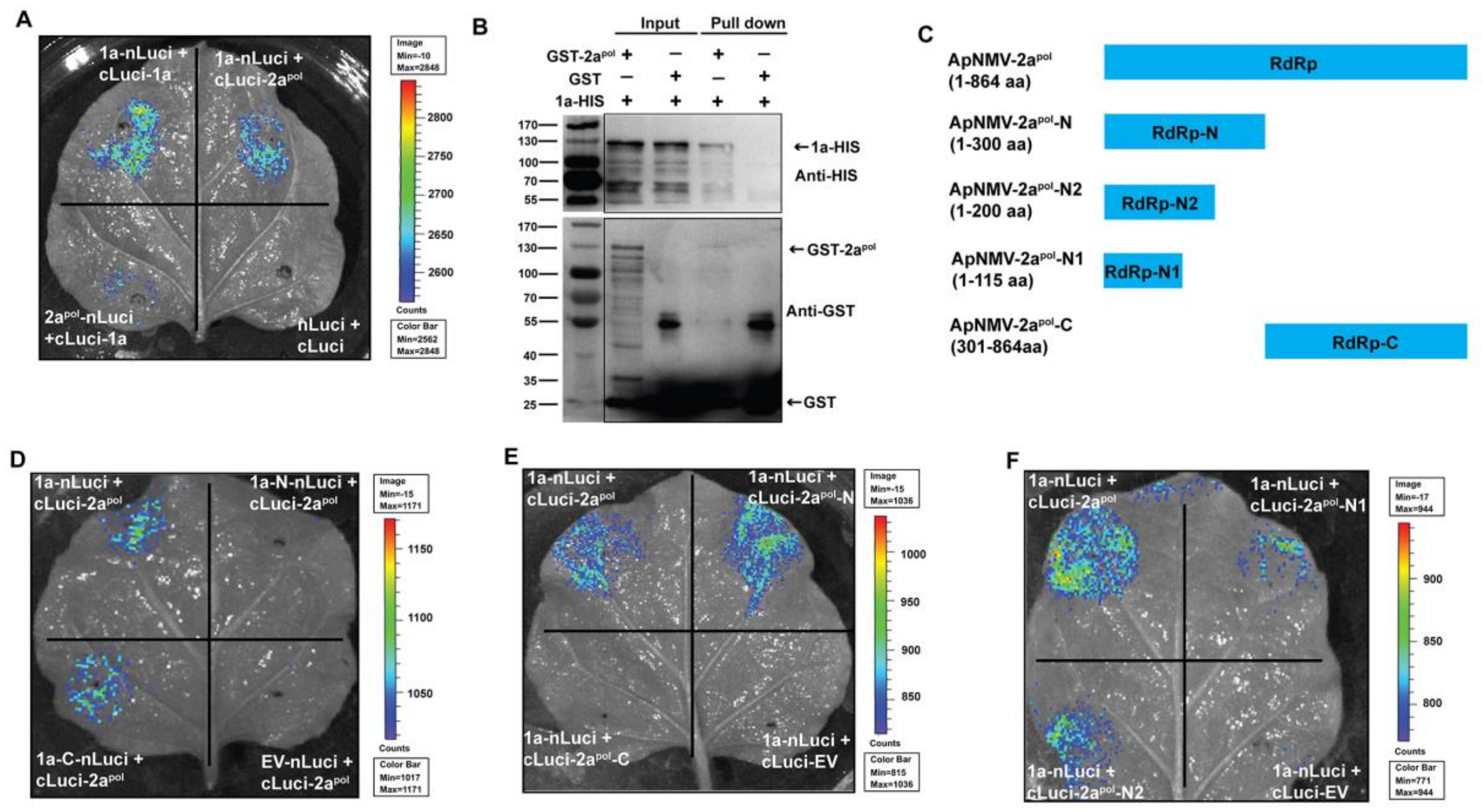
3.5. ApNMV 1a Interacts with 2apol
4. Discussion
4.1. The ApNMV–Lw Isolate was Placed in Subgroup 3 of Ilarvirus
4.2. Interaction between Replication Proteins is a Conserved Feature among Tripartite RNA Viruses in Bromoviridae
4.3. Interactions between Replication Proteins are Critical for Genome Replication and VRC Assembly of Tripartite RNA Viruses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ji, Z.; Zhao, X.; Duan, H.; Hu, T.; Wang, S.; Wang, Y.; Cao, K. Multiplex RT-PCR detection and distribution of four apple viruses in China. Acta Virol. 2013, 57, 435–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thokchom, T.; Rana, T.; Hallan, V.; Ram, R.; Zaidi, A.A. Molecular characterization of the Indian strain of Apple mosaic virus isolated from apple (Malus domestica). Phytoparasitica 2009, 37, 375–379. [Google Scholar] [CrossRef]
- Cembali, T.; Folwell, R.J.; Wandschneider, P.; Eastwell, K.C.; Howell, W.E. Economic implications of a virus prevention program in deciduous tree fruits in the US. Crop. Prot. 2003, 22, 1149–1156. [Google Scholar] [CrossRef]
- Chai, G.; Song, L.; Jiang, Z.; Zhang, X.; Zhang, S.; Liu, M.; Tang, Y.; Sun, Y.; Zhao, L. The effect of apple mosaic on photosynthesis of different varieties of apple. Yantai Fruits 2017, 3, 8–9. [Google Scholar]
- Bradford, F.; Joley, L. Infectious variegation in the apple. Agric. Res. 1933, 46, 901–908. [Google Scholar]
- Posnette, A.; Cropley, R. Apple mosaic virus. Host reactions and strain interference. J. Hortic. Sci. 1956, 31, 119–133. [Google Scholar] [CrossRef]
- Petrzik, K.; Lenz, O. Apple mosaic virus in pome fruits. In Virus and Virus-Like Disease of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; APS Press: St. Paul, MN, USA, 2011; pp. 25–28. [Google Scholar]
- Grimova, L.; Winkowska, L.; Konrady, M.; Rysanek, P. Apple mosaic virus. Phytopathology 2016, 55, 1–19. [Google Scholar]
- Bujarski, J.; Figlerowicz, M.; Gallitelli, D.; Roossinck, M.; Scott, S. Family Bromoviridae. In Virus Taxonomy, Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M., Lefkowitz, E., Adams, M.J., Carstens, E.B., Eds.; Acadamic Press: London, UK, 2012; pp. 965–976. [Google Scholar]
- Hu, G.; Dong, Y.; Zhang, Z.; Fan, X.; Ren, F.; Li, Z.; Zhou, J. First report of prunus necrotic ringspot virus infection of apple in China. Plant Dis. 2016, 100, 1995. [Google Scholar] [CrossRef]
- Liang, P.; Zhang, Z.; Liu, F.; Lu, G.; Li, S.; Wang, H. Problems of identification of pathogens associated with apple mosaic symptom and the exploration of its potential pathogens. J. Fruit Sci. 2016, 33, 332–339. [Google Scholar]
- Hu, Y.; Shi, H.; Jing, C.; Li, K.; Sun, X.; Zhou, C.; QIng, J. First report of cucumber mosaic virus infecting apple in China. J. Plant Pathol. 2016, 98, 181. [Google Scholar]
- Noda, H.; Yamagishi, N.; Yaegashi, H.; XIng, F.; Xie, J.; Li, S.; ZHou, T.; Ito, T.; Yoshikawa, N. Apple necrotic mosaic virus, a novel ilarvirus from mosaic-disease apple trees in Japan and China. J. Gen. Plant Pathol. 2017, 83, 83–90. [Google Scholar] [CrossRef]
- Xing, F.; Robe, B.L.; Zhang, Z.; Wang, H.; Li, S. Genomic Analysis, Sequence Diversity, and Occurrence of Apple necrotic mosaic virus, a Novel Ilarvirus Associated with Mosaic Disease of Apple Trees in China. Plant Dis. 2018, 102, 1841–1847. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.; Dong, Y.; Zhang, Z.; Fan, X.; Ren, F. Molecular characterization of Apple necrotic mosaic virus identified in crabapple (Malus spp.) tree of China. J. Integr. Agric. 2019, 18, 698–701. [Google Scholar] [CrossRef]
- Rivers, T. VIruses and Koch’s postulates. J. Bacteriol. 1937, 32, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Y.; Mao, K.; Zhao, C.; Zhao, X.Y.; Zhang, H.L.; Shu, H.R.; Hao, Y.J. MdCOP1 ubiquitin E3 ligases interact with MdMYB1 to regulate light-induced anthocyanin biosynthesis and red fruit coloration in apple. Plant Physiol. 2012, 160, 1011–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Zou, Y.; Shang, Y.; Lin, H.; Wang, Y.; Cai, R.; Tang, X.; Zhou, J.M. Firefly luciferase complementation imaging assay for protein-protein interactions in plants. Plant Physiol. 2008, 146, 368–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, M.; Chaban, C.; Schutze, K.; Batistic, O.; Weckermann, K.; Nake, C.; Blazevic, D.; Grefen, C.; Schumacher, K.; Oecking, C.; et al. Visualization of protein interactions in living plant cells using bimolecular fluorescence complementation. Plant J. 2004, 40, 428–438. [Google Scholar] [CrossRef]
- Kohrer, K.; Domdey, H. Preparation of high molecular weight RNA. Methods Enzymol. 1991, 194, 398–405. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbour-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Wang, X.F.; An, J.P.; Liu, X.; Su, L.; You, C.X.; Hao, Y.J. The Nitrate-Responsive Protein MdBT2 Regulates Anthocyanin Biosynthesis by Interacting with the MdMYB1 Transcription Factor. Plant Physiol. 2018, 178, 890–906. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ahlquist, P.; Brome, M.V. Encyclopedia of Virology; Elsevier: Oxford, UK, 2008; pp. 381–386. [Google Scholar]
- Hammond, R.W.; Crosslin, J.M. The complete nucleotide sequence of RNA 3 of a peach isolate of Prunus necrotic ringspot virus. Virology 1995, 208, 349–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, A.; Wang, X. Bromovirus-induced remodeling of host membranes during viral RNA replication. Curr. Opin. Virol. 2014, 9, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Wood, G.; Chamberlain, E.; Atkinson, H.; Hunter, J. Field studies with apple mosaic virus. N. Z. J. Agric. Res. 1975, 18, 399–404. [Google Scholar] [CrossRef]
- Li, D.; Zhao, H.; Hu, Z.; HU, X.; ZHang, Y. Studies on the damage, loss of production and control to apple mosaic disease. J. NW Sci. Tech. Univ. Agric. For. 2002, 30, 77–80. [Google Scholar]
- Sanchez-Navarro, J.; Plallas, V. Nucleotide sequence of apple mosaic ilarvirus RNA4. J. Gen. Virol. 1994, 75, 1441–1445. [Google Scholar] [CrossRef]
- Shiel, P.; Alrefai, R.; Domier, L.; Korban, S.; Berger, P. The complete nucleotide sequence of apple mosaic virus RNA-3. Arch. Virol. 1995, 140, 1247–1256. [Google Scholar] [CrossRef]
- Shiel, P.; Berger, P. The complete nucleotide sequence of apple mosaic virus (ApMV) RNA1 and RNA2: ApMV is more closely related to alfalfa mosaic virus than to other ilarviruses. J. Gen. Virol. 2000, 81, 273–278. [Google Scholar] [CrossRef]
- O’Reilly, E.K.; Wang, Z.; French, R.; Kao, C.C. Interactions between the structural domains of the RNA replication proteins of plant-infecting RNA viruses. J. Virol. 1998, 72, 7160–7169. [Google Scholar] [CrossRef] [Green Version]
- Diaz, A.; Gallei, A.; Ahlquist, P. Bromovirus RNA replication compartment formation requires concerted action of 1a’s self-interacting RNA capping and helicase domains. J. Virol. 2012, 86, 821–834. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, E.K.; Paul, J.D.; Kao, C.C. Analysis of the interaction of viral RNA replication proteins by using the yeast two-hybrid assay. J. Virol. 1997, 71, 7526–7532. [Google Scholar] [CrossRef] [Green Version]
- Kao, C.C.; Ahlquist, P. Identification of the domains required for direct interaction of the helicase-like and polymerase-like RNA replication proteins of brome mosaic virus. J. Virol. 1992, 66, 7293–7302. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, E.K.; Tang, N.; Ahlquist, P.; Kao, C.C. Biochemical and genetic analyses of the interaction between the helicase-like and polymerase-like proteins of the brome mosaic virus. Virology 1995, 214, 59–71. [Google Scholar] [CrossRef] [Green Version]
- RestrepoHartwig, M.A.; Ahlquist, P. Brome mosaic virus helicase- and polymerase-like proteins colocalize on the endoplasmic reticulum at sites of viral RNA synthesis. J. Virol. 1996, 70, 8908–8916. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Ahlquist, P. Brome mosaic virus polymerase-like protein 2a is directed to the endoplasmic reticulum by helicase-like viral protein 1a. J. Virol. 2000, 74, 4310–4318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restrepo-Hartwig, M.; Ahlquist, P. Brome mosaic virus RNA replication proteins 1a and 2a colocalize and 1a independently localizes on the yeast endoplasmic reticulum. J. Virol. 1999, 73, 10303–10309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Westler, W.M.; den Boon, J.A.; Wang, X.; Diaz, A.; Steinberg, H.A.; Ahlquist, P. An amphipathic alpha-helix controls multiple roles of brome mosaic virus protein 1a in RNA replication complex assembly and function. PLoS Pathog. 2009, 5, e1000351. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lee, W.M.; Watanabe, T.; Schwartz, M.; Janda, M.; Ahlquist, P. Brome mosaic virus 1a nucleoside triphosphatase/helicase domain plays crucial roles in recruiting RNA replication templates. J. Virol. 2005, 79, 13747–13758. [Google Scholar] [CrossRef] [Green Version]
- Ahola, T.; den Boon, J.A.; Ahlquist, P. Helicase and capping enzyme active site mutations in brome mosaic virus protein 1a cause defects in template recruitment, negative-strand RNA synthesis, and viral RNA capping. J. Virol. 2000, 74, 8803–8811. [Google Scholar] [CrossRef] [Green Version]
- Kroner, P.A.; Young, B.M.; Ahlquist, P. Analysis of the role of brome mosaic virus la protein domains in RNA replication, using linker insertion mutagenesis. J. Virol. 1990, 64, 6110–6120. [Google Scholar] [CrossRef] [Green Version]
- Kao, C.C.; Quadt, R.; Hershberger, R.P.; Ahlquist, P. Brome mosaic virus RNA replication proteins 1a and 2a from a complex in vitro. J. Virol. 1992, 66, 6322–6329. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ApNMV-Qu | ApNMV-Hua | ApNMV-Hai | ApNMV-JP | ApMV | PNRSV | ||
|---|---|---|---|---|---|---|---|
| nt % | RNA1 | 97.37 | 94.00 | 97.17 | 96.33 | 59.44 | 64.00 |
| RNA2 | 96.45 | 94.91 | 93.55 | 94.23 | 58.02 | 58.20 | |
| RNA3 | 96.10 | 88.47 | 93.04 | 91.57 | 49.73 | 59.39 | |
| aa % | 1a | 99.53 | 99.05 | 99.24 | 98.67 | 63.72 | 69.60 |
| 2apol | 96.76 | 94.56 | 96.06 | 93.29 | 57.08 | 60.07 | |
| MP | 96.79 | 92.86 | 94.29 | 93.21 | 50.70 | 55.83 | |
| CP | 96.35 | 94.52 | 95.89 | 95.89 | 40.54 | 56.83 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.-L.; Zhang, F.-J.; Zheng, P.-F.; Xie, Y.-H.; You, C.-X.; Hao, Y.-J. Determination of Protein Interactions among Replication Components of Apple Necrotic Mosaic Virus. Viruses 2020, 12, 474. https://doi.org/10.3390/v12040474
Zhang Z-L, Zhang F-J, Zheng P-F, Xie Y-H, You C-X, Hao Y-J. Determination of Protein Interactions among Replication Components of Apple Necrotic Mosaic Virus. Viruses. 2020; 12(4):474. https://doi.org/10.3390/v12040474
Chicago/Turabian StyleZhang, Zhen-Lu, Fu-Jun Zhang, Peng-Fei Zheng, Yin-Huan Xie, Chun-Xiang You, and Yu-Jin Hao. 2020. "Determination of Protein Interactions among Replication Components of Apple Necrotic Mosaic Virus" Viruses 12, no. 4: 474. https://doi.org/10.3390/v12040474
APA StyleZhang, Z. -L., Zhang, F. -J., Zheng, P. -F., Xie, Y. -H., You, C. -X., & Hao, Y. -J. (2020). Determination of Protein Interactions among Replication Components of Apple Necrotic Mosaic Virus. Viruses, 12(4), 474. https://doi.org/10.3390/v12040474