Diversity of Astroviruses Circulating in Humans, Bats, and Wild Birds in Egypt


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.1.1. Human Samples
2.1.2. Bat Samples
2.1.3. Migratory Bird Samples
2.2. Detection of Astrovirus in Bats and Humans
2.3. Detection of Astroviruses in Wild Birds
2.4. Genetic and Phylogenetic Analysis of Astroviruses
2.5. Statistical Analysis
2.6. Ethics Approval
3. Results
3.1. Human Samples
3.2. Bat Samples
3.3. Wild Birds
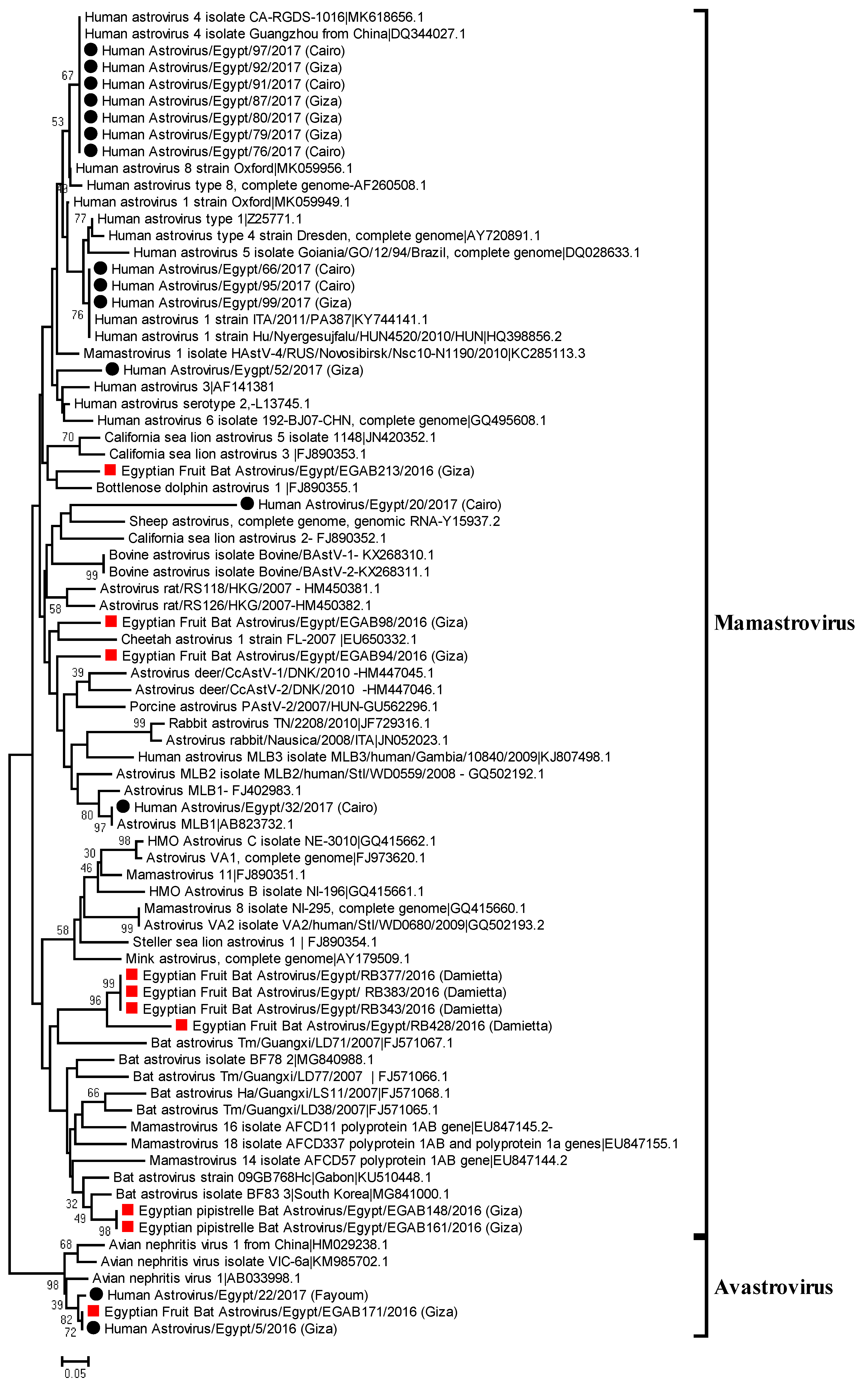
3.4. Phylogenetic Analysis of Bat and Human Astroviruses
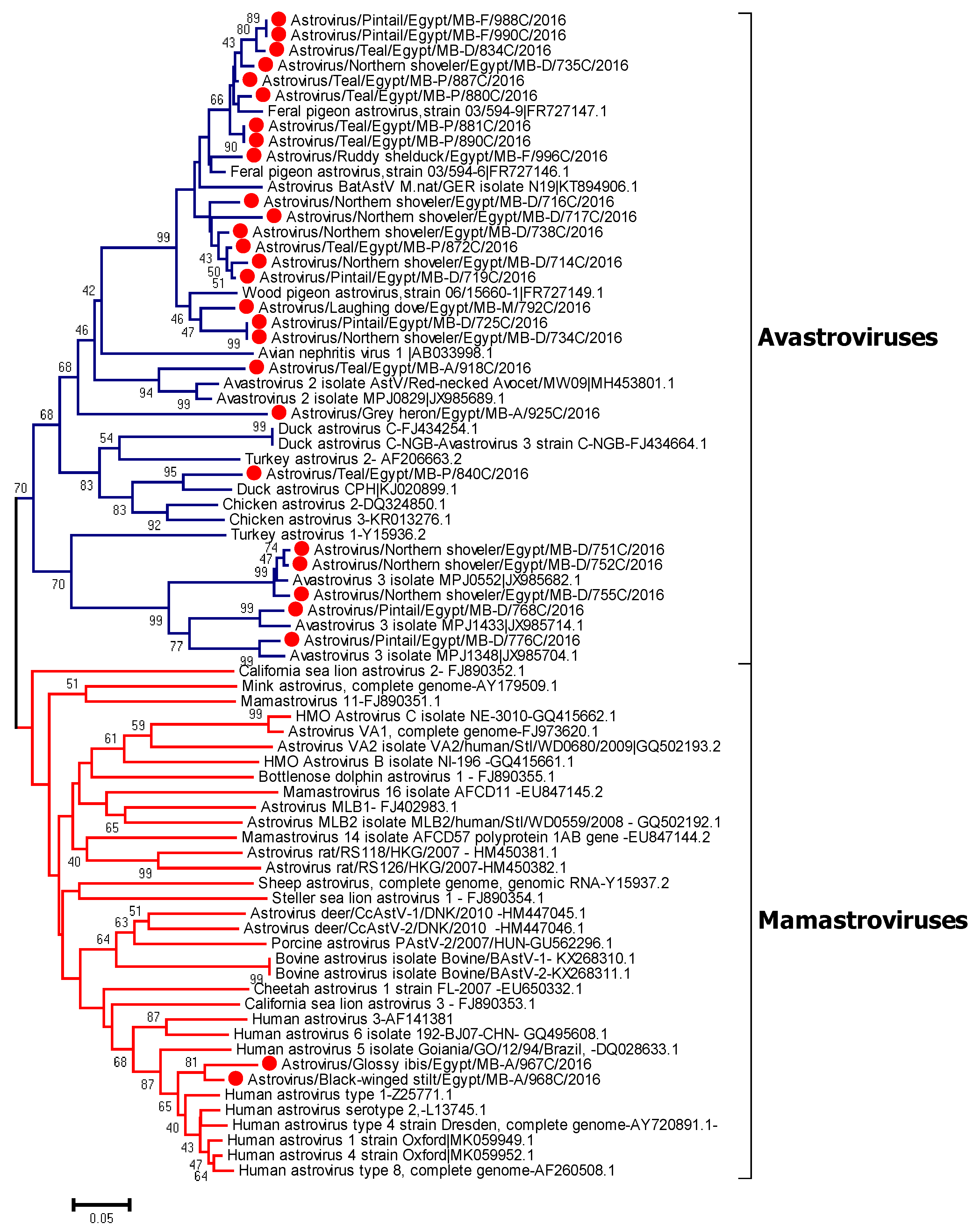
3.5. Phylogenetic Analysis of Avian Astroviruses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Madeley, C.R.; Cosgrove, B.P. Letter: 28 nm particles in faeces in infantile gastroenteritis. Lancet 1975, 2, 451–452. [Google Scholar] [CrossRef]
- Mendenhall, I.H.; Smith, G.J.; Vijaykrishna, D. Ecological drivers of virus evolution: Astrovirus as a case study. J. Virol. 2015, 89, 6978–6981. [Google Scholar] [CrossRef] [Green Version]
- Mendez, E.; Munoz-Yanez, C.; Sanchez-San Martin, C.; Aguirre-Crespo, G.; Banos-Lara Mdel, R.; Gutierrez, M.; Espinosa, R.; Acevedo, Y.; Arias, C.F.; Lopez, S. Characterization of human astrovirus cell entry. J. Virol. 2014, 88, 2452–2460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2011, 11, 1529–1544. [Google Scholar] [CrossRef]
- Imada, T.; Yamaguchi, S.; Mase, M.; Tsukamoto, K.; Kubo, M.; Morooka, A. Avian nephritis virus (anv) as a new member of the family astroviridae and construction of infectious anv cdna. J. Virol. 2000, 74, 8487–8493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moser, L.A.; Schultz-Cherry, S. Pathogenesis of astrovirus infection. Viral. Immunol. 2005, 18, 4–10. [Google Scholar] [CrossRef]
- Woolhouse, M.E.; Gowtage-Sequeria, S. Host range and emerging and reemerging pathogens. Emerg. Infect. Dis. 2005, 11, 1842–1847. [Google Scholar] [CrossRef]
- Parrish, C.R.; Holmes, E.C.; Morens, D.M.; Park, E.C.; Burke, D.S.; Calisher, C.H.; Laughlin, C.A.; Saif, L.J.; Daszak, P. Cross-species virus transmission and the emergence of new epidemic diseases. Microbiol. Mol. Biol. Rev. Mmbr. 2008, 72, 457–470. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, A.; Duong, V.; Hul, V.; San, S.; Davun, H.; Omaliss, K.; Chea, S.; Hassanin, A.; Theppangna, W.; Silithammavong, S.; et al. Diversity of bat astroviruses in lao pdr and cambodia. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2017, 47, 41–50. [Google Scholar] [CrossRef]
- Rougeron, V.; Suquet, E.; Maganga, G.D.; Jiolle, D.; Mombo, I.M.; Bourgarel, M.; Motsch, P.; Arnathau, C.; Durand, P.; Drexler, F.; et al. Characterization and phylogenetic analysis of new bat astroviruses detected in gabon, central africa. Acta Virol. 2016, 60, 386–392. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Son, K.D.; Yong-Sik, K.; Wang, S.J.; Kim, Y.K.; Jheong, W.H.; Oem, J.K. Genetic diversity and phylogenetic analysis of newly discovered bat astroviruses in korea. Arch. Virol. 2018, 163, 3065–3072. [Google Scholar] [CrossRef] [PubMed]
- Amoroso, M.G.; Russo, D.; Lanave, G.; Cistrone, L.; Pratelli, A.; Martella, V.; Galiero, G.; Decaro, N.; Fusco, G. Detection and phylogenetic characterization of astroviruses in insectivorous bats from central-southern italy. Zoonoses Public Health 2018, 65, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Li, J.; Hu, G.; Chen, Z.; Wu, Y.; Chen, Y.; Chen, Z.; Liao, Y.; Zhou, J.; Ke, X.; et al. Isolation and phylogenetic characterization of bat astroviruses in southern china. Arch. Virol. 2011, 156, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Hoarau, F.; Le Minter, G.; Joffrin, L.; Schoeman, M.C.; Lagadec, E.; Ramasindrazana, B.; Dos Santos, A.; Goodman, S.M.; Gudo, E.S.; Mavingui, P.; et al. Bat astrovirus in mozambique. Virol. J. 2018, 15, 104. [Google Scholar] [CrossRef] [Green Version]
- Denny, P. Africa; Finlayson, M., Moser, M., Eds.; International Waterfowl and Wetlands Research Bureau: London, UK, 1991. [Google Scholar]
- Alexander, D.J. An overview of the epidemiology of avian influenza. Vaccine 2007, 25, 5637–5644. [Google Scholar] [CrossRef]
- Chu, D.K.; Poon, L.L.; Guan, Y.; Peiris, J.S. Novel astroviruses in insectivorous bats. J. Virol. 2008, 82, 9107–9114. [Google Scholar] [CrossRef] [Green Version]
- Atkins, A.; Wellehan, J.F., Jr.; Childress, A.L.; Archer, L.L.; Fraser, W.A.; Citino, S.B. Characterization of an outbreak of astroviral diarrhea in a group of cheetahs (acinonyx jubatus). Vet. Microbiol. 2009, 136, 160–165. [Google Scholar] [CrossRef]
- Naficy, A.B.; Rao, M.R.; Holmes, J.L.; Abu-Elyazeed, R.; Savarino, S.J.; Wierzba, T.F.; Frenck, R.W.; Monroe, S.S.; Glass, R.I.; Clemens, J.D. Astrovirus diarrhea in egyptian children. J. Infect. Dis. 2000, 182, 685–690. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.F.; Sebeny, P.J.; Klena, J.D.; Pimentel, G.; Mansour, A.; Naguib, A.M.; Bruton, J.; Young, S.Y.; Holtz, L.R.; Wang, D. Novel astroviruses in children, egypt. Emerg. Infect. Dis. 2011, 17, 2391–2393. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, F.A.M.; Al-Attar, B.; Ahmad, M.K.; Follero, P.M. Burden of disease: Prevalence and incidence of endstage renal disease in middle eastern countries. Clin. Nephrol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Dufkova, L.; Strakova, P.; Sirmarova, J.; Salat, J.; Moutelikova, R.; Chrudimsky, T.; Bartonicka, T.; Nowotny, N.; Ruzek, D. Detection of diverse novel bat astrovirus sequences in the czech republic. Vector Borne Zoonotic Dis. 2015, 15, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.; Leung, C.Y.; Perera, H.K.; Ng, E.M.; Gilbert, M.; Joyner, P.H.; Grioni, A.; Ades, G.; Guan, Y.; Peiris, J.S.; et al. A novel group of avian astroviruses in wild aquatic birds. J. Virol. 2012, 86, 13772–13778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Correa, I.; Truchado, D.A.; Gomez-Lucia, E.; Domenech, A.; Perez-Tris, J.; Schmidt-Chanasit, J.; Cadar, D.; Benitez, L. A novel group of avian astroviruses from neotropical passerine birds broaden the diversity and host range of astroviridae. Sci. Rep. 2019, 9, 9513. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, M.N.F.; Abd El-Daim, S.E.; Ahmed, N.I.; Elmahdy, E.M. Molecular detection of three gastroenteritis viruses in an urban sewage treatment plant and river water in egypt. Egypt. J. Aquat. Biol. Fish. 2018, 22, 615–627. [Google Scholar]
- Morsy El-Senousy, W.; Guix, S.; Abid, I.; Pinto, R.M.; Bosch, A. Removal of astrovirus from water and sewage treatment plants, evaluated by a competitive reverse transcription-pcr. Appl. Environ. Microbiol. 2007, 73, 164–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxendale, W.; Mebatsion, T. The isolation and characterisation of astroviruses from chickens. Avian. Pathol. 2004, 33, 364–370. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Variable | No. Collected Samples (% *) | No. of Astroviruses-Positive Samples (% †) | p-Value |
|---|---|---|---|
| Human | 100 | 28 (28) | |
| Governorate | |||
| Fayoum | 2 (2) | 1 (50) | NS |
| Giza | 64 (64) | 20 (31.25) | |
| Cairo | 26 (26) | 7 (26.92) | |
| Alexandria | 1 (1) | 0 (0) | |
| Aswan | 1 (1) | 0(0) | |
| Qaulibia | 4 (4) | 0 (0) | |
| Suiz | 2 (2) | 0(0) | |
| Sex | |||
| Male | 66 (66) | 15 (22.72) | p < 0.05 |
| Female | 34 (34) | 13 (38.23) | |
| Age | |||
| ≤1 | 71 (71) | 25 (35.21) | p < 0.05 |
| >1 | 29 (29) | 3 (10.34) | |
| Bats | 417 | 119 (28.53) | |
| Governorate | |||
| Damietta | 116 (27.81) | 13 (11.21) | p < 0.05 |
| Giza | 301 (72.19) | 106 (35.22) | |
| Species | |||
| Egyptian Fruit Bat (Rousettus aegyptiacus) | 288 (69.06) | 96 (33.33) | p < 0.05 |
| Egyptian Pipistrelle (Pipistrellus aegyptius) | 93 (22.3) | 14 (15.05) | |
| Egyptian slit-faced bat (Nycteris thebaica) | 25 (5.99) | 6 (24) | |
| Egyptian tomb bat (Taphozous perforates) | 11 (2.64) | 3 (27.27) | |
| Age | |||
| Adult | 407 (97.6) | 116 (28.5) | NS |
| Juvenile | 10 (2.4) | 3 (30) | |
| Sex | |||
| Male | 170 (40.7) | 58 (34.11) | NS |
| Female | 131 (31.41) | 48 (36.64) | |
| Unknown | 116 (27.82) | 13 (11.20) | |
| Birds | 301 | 80 (26.5) | |
| Governorate | |||
| Aswan | 79 (26.25) | 13 (16.45) | p < 0.05 |
| Damietta | 113 (37.54) | 42 (13.95) | |
| Fayoum | 19 (6.31) | 10 (52.63) | |
| Mersa Matruh | 24 (7.97) | 4 (16.66) | |
| Port Said | 66 (21.92) | 11 (16.66) | |
| Species | |||
| African swamphen (Porphyrio madagascariensis) | 5 (1.66) | 0 (0) | p < 0.05 |
| Cormorant (Phalacrocorax carbo) | 2 (0.66) | 0 (0) | |
| Squacco heron (Ardeola ralloides) | 2 (0.66) | 1(50) | |
| Glossy ibis (Plegadis falcinellus) | 4 (1.33) | 1 (25) | |
| Grey heron (Ardea cinereal) | 10 (3.32) | 3 (30) | |
| Laughing dove (Spilopelia senegalensis) | 6 (1.99) | 1 (16.66) | |
| Moorhen (Gallinula chloropus) | 2 (0.66) | 0(0) | |
| Teal (Anas crecca) | 65 (21.59) | 15 (23.07) | |
| Wigeon (Anas Penelope) | 1 (0.33) | 1 (100) | |
| Black-winged stilt (Himantopus himantopus) | 9 (2.99) | 1 (11.11) | |
| Coot (Fulica atra) | 12 (3.98) | 2 (16.66) | |
| Egyptian goose (Alopochen aegyptiacus) | 5 (1.66) | 1 (20) | |
| Mallard (Anas platyrhynchos) | 6 (1.99) | 3 (50) | |
| Namaqua dove (Oena capensis) | 4 (1.33) | 0 (0) | |
| Northern shoveler (Anas clypeat) | 101 (33.55) | 26 (25.74) | |
| Pintail (Anas acuta) | 44 (14.61) | 20 (45.45) | |
| Purple heron (Ardea purpurea) | 4 (1.32) | 1 (25) | |
| Spotted redshank (Tringa erythropus) | 10 (3.32) | 2 (20) | |
| Habitat | |||
| Migratory birds | 270 (89.70) | 74 (27.40) | NS |
| Resident birds | 31 (10.30) | 6 (19.35) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Taweel, A.; Kandeil, A.; Barakat, A.; Alfaroq Rabiee, O.; Kayali, G.; Ali, M.A. Diversity of Astroviruses Circulating in Humans, Bats, and Wild Birds in Egypt. Viruses 2020, 12, 485. https://doi.org/10.3390/v12050485
El Taweel A, Kandeil A, Barakat A, Alfaroq Rabiee O, Kayali G, Ali MA. Diversity of Astroviruses Circulating in Humans, Bats, and Wild Birds in Egypt. Viruses. 2020; 12(5):485. https://doi.org/10.3390/v12050485
Chicago/Turabian StyleEl Taweel, Ahmed, Ahmed Kandeil, Ahmed Barakat, Omar Alfaroq Rabiee, Ghazi Kayali, and Mohamed Ahmed Ali. 2020. "Diversity of Astroviruses Circulating in Humans, Bats, and Wild Birds in Egypt" Viruses 12, no. 5: 485. https://doi.org/10.3390/v12050485
APA StyleEl Taweel, A., Kandeil, A., Barakat, A., Alfaroq Rabiee, O., Kayali, G., & Ali, M. A. (2020). Diversity of Astroviruses Circulating in Humans, Bats, and Wild Birds in Egypt. Viruses, 12(5), 485. https://doi.org/10.3390/v12050485