Multifaceted Functions of Host Cell Caveolae/Caveolin-1 in Virus Infections


Abstract
:1. Introduction
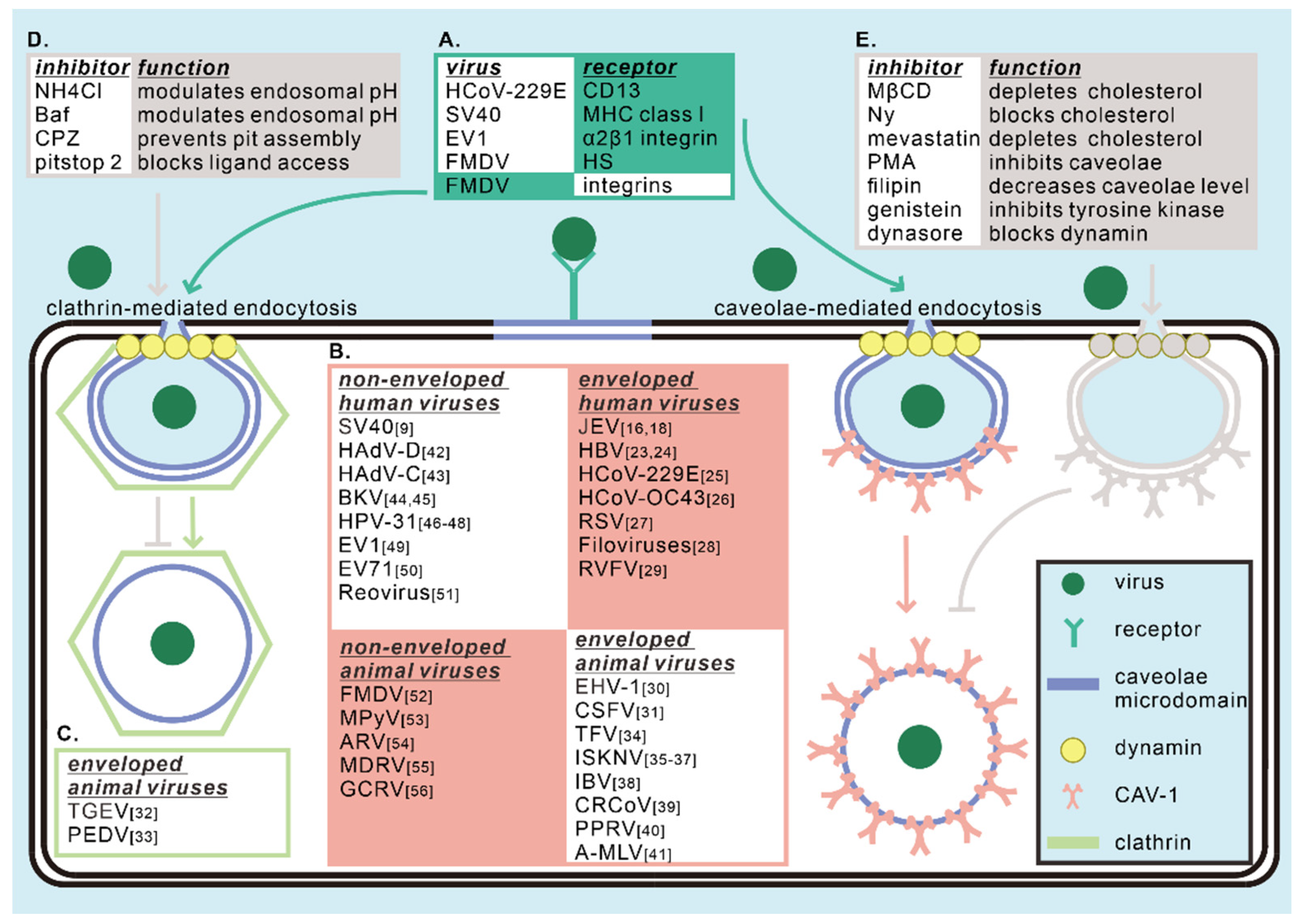
2. The Role of Caveolae/CAV-1 in Virus Entry
2.1. The Role of Caveolae/CAV-1 in the Entry of Enveloped Human Viruses

{kind=link}
{kind=link}
{kind=link}
| Classification | Viruses | Family | Host Cells | Experimental Approaches | References |
|---|---|---|---|---|---|
| Human enveloped viruses | JEV | Flaviviridae | Rat neuroblastoma cells, human neuroblastoma SK-N-SH cells | Chemical inhibitors, RNAi, dominant-negative constructs, Fluorescence imaging | [16,18] |
| HBV | Hepadnaviridae | COS-7 cells, HepaRG cells | Live cell imaging, chemical inhibitors, dominant-negative constructs | [23,24] | |
| HCoV-229E | Coronaviridae | Human fibroblasts L132 | Fluorescence imaging, electron microscopy, siRNA, chemical inhibitors | [25] | |
| HCoV-OC43 | Coronaviridae | HCT-8 cells | Fluorescence imaging, siRNA, chemical inhibitors | [26] | |
| RSV | Paramyxoviridae | Cattle dendritic cells | Chemical inhibitors, fluorescence imaging | [27] | |
| Filoviruses (EBOV, MARV pseudotype viruses) | Filoviridae | 293T cells and Hela cells | Chemical inhibitors, fluorescence imaging, internalization kinetics | [28] | |
| RVFV | Phenuiviridae | HeLa, HepG2, and 293T cells | Chemical inhibitors, RNAi, dominant-negative constructs, fluorescence imaging | [29] | |
| Animal enveloped viruses | EHV-1 | Herpesviridae | Equine brain microvascular endothelial cells | Chemical inhibitors, dominant-negative constructs, fluorescence imaging | [30] |
| CSFV | Flaviviridae | Macrophages | Digital gene expression profiling, fluorescence imaging, siRNA, CAV-1 overexpression | [31] | |
| TGEV | Coronaviridae | Swine testis cells | Live cell imaging, chemical inhibitors | [32] | |
| PEDV | Coronaviridae | Vero cells and IPEC-J2 cells | Internalization kinetics, chemical inhibitors, fluorescence imaging, fractionation | [33] | |
| TFV | Iridoviridae | HepG2 cells | Chemical inhibitors, CAV-1 peptide, fluorescence imaging | [34] | |
| ISKNV | Iridoviridae | MFF-1 cells | Internalization kinetics, chemical inhibitors, fluorescence imaging, far-western blotting, pulldown, coimmunoprecipitation, siRNA, truncation, fractionation | [35,36,37] | |
| IBV | Coronaviridae | Vero cells | Fractionation, fluorescence imaging, chemical inhibitors | [38] | |
| CRCoV | Coronaviridae | HRT-18G cells | Chemical inhibitors, fluorescence imaging, siRNA | [39] | |
| PPRV | Paramyxoviridae | Caprine endometrial epithelial cells | Internalization kinetics, chemical inhibitors, fluorescence imaging, siRNA, electron microscopy | [40] | |
| A-MLV | Retroviridae | NIH 3T3 cells | Internalization kinetics, chemical inhibitors, immunoprecipitation, fluorescence imaging | [41] | |
| Human non-enveloped viruses | SV40 | Polyomaviridae | CV-1 cells | Internalization kinetics, chemical inhibitors, fractionation | [9] |
| HAdV-D | Adenoviridae | Corneal cells | Chemical inhibitors, fluorescence imaging, siRNA, fractionation, electron microscopy, animal model | [42] | |
| HAdV-C | Adenoviridae | Plasmocytic cell lines | Chemical inhibitors, dominant-negative constructs, fluorescence imaging | [43] | |
| BKV | Polyomaviridae | Vero cells and human renal proximal tubular epithelial cells | Internalization kinetics, chemical inhibitors, dominant-negative constructs | [44,45] | |
| HPV-31 | Papillomaviridae | COS-7 cells and human keratinocytes | Electron microscopy, chemical inhibitors, dominant-negative constructs, internalization kinetics, fluorescence imaging, flotation | [46,47,48] | |
| EV1 | Picornaviridae | SAOS cells | Fluorescence imaging, electron microscopy, dominant-negative constructs, fractionation | [49] | |
| EV71 | Picornaviridae | Jurkat T and mouse L929 cells | siRNA, chemical inhibitors, fluorescence imaging | [50] | |
| Reovirus | Reoviridae | A549, Hela and HEK-293 cells | Chemical inhibitors, dominant-negative constructs, fluorescence imaging | [51] | |
| Animal non-enveloped viruses | FMDV | Picornaviridae | MCF-10A cells | Fluorescence imaging, chemical inhibitors, siRNA | [52] |
| MPyV | Polyomaviridae | NIH 3T6 fibroblasts and NMuMG epithelial cells | Electron microscopy, chemical inhibitors, fluorescence imaging | [53] | |
| ARV | Reoviridae | Vero and DF-1 cells | Chemical inhibitors, siRNA, coimmunoprecipitation, fluorescence imaging | [54] | |
| MDRV | Reoviridae | Vero and DF-1 cells | Chemical inhibitors, fluorescence imaging | [55] | |
| GCRV | Reoviridae | CIK cells | Chemical inhibitors, fluorescence imaging, dominant-negative constructs | [56] |
2.2. The Role of Caveolae/CAV-1 in the Entry of Enveloped Animal Viruses
2.3. The Role of Caveolae/CAV-1 in the Entry of Non-Enveloped Human Viruses
2.4. The Role of Caveolae/CAV-1 in the Entry of Non-Enveloped Animal Viruses
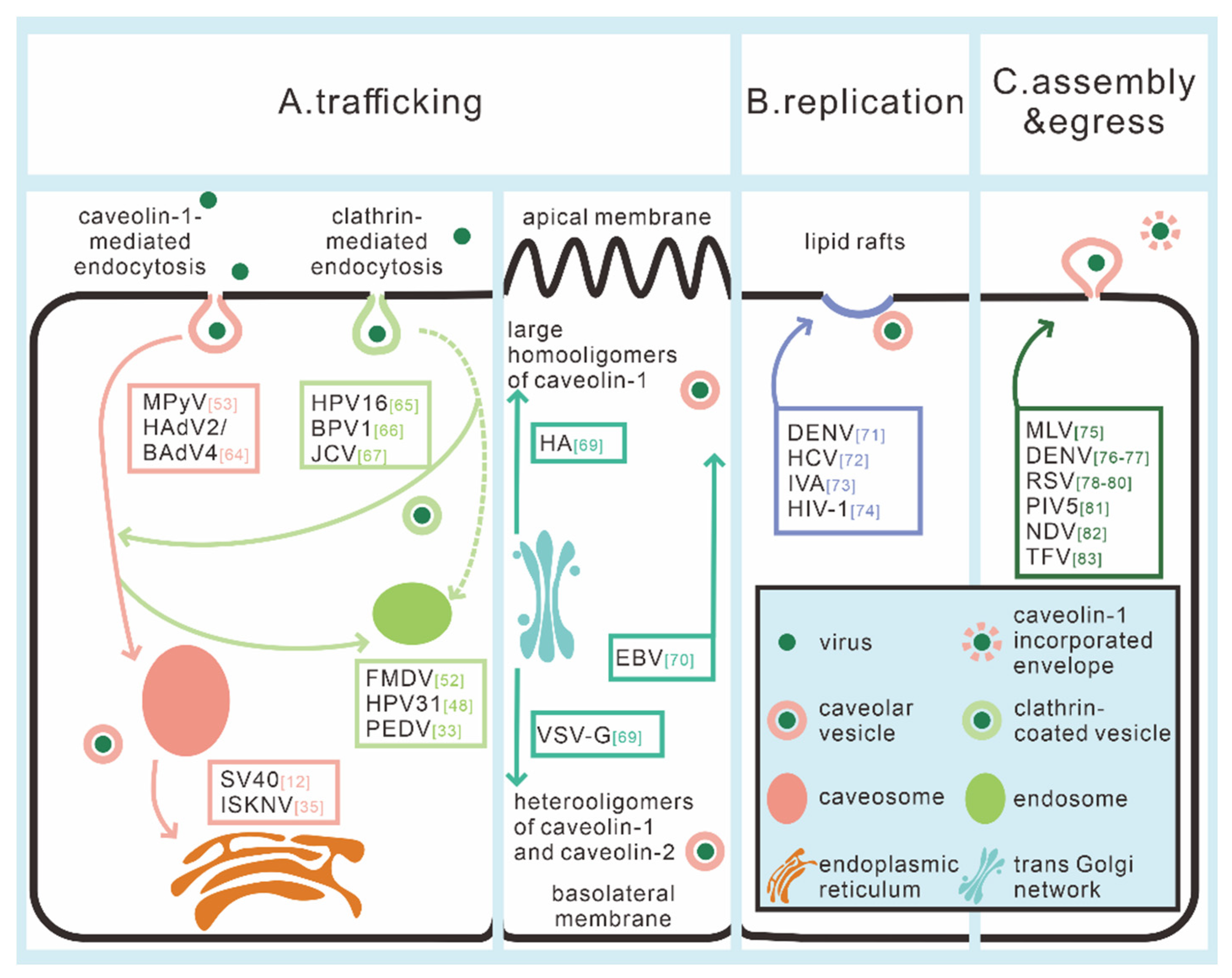
3. The Role of Caveolae/CAV-1 in Virus Intracellular Trafficking
4. The Role of Caveolae/CAV-1 in Virus Replication
5. The Role of Caveolae/CAV-1 in Virus Assembly and Egress
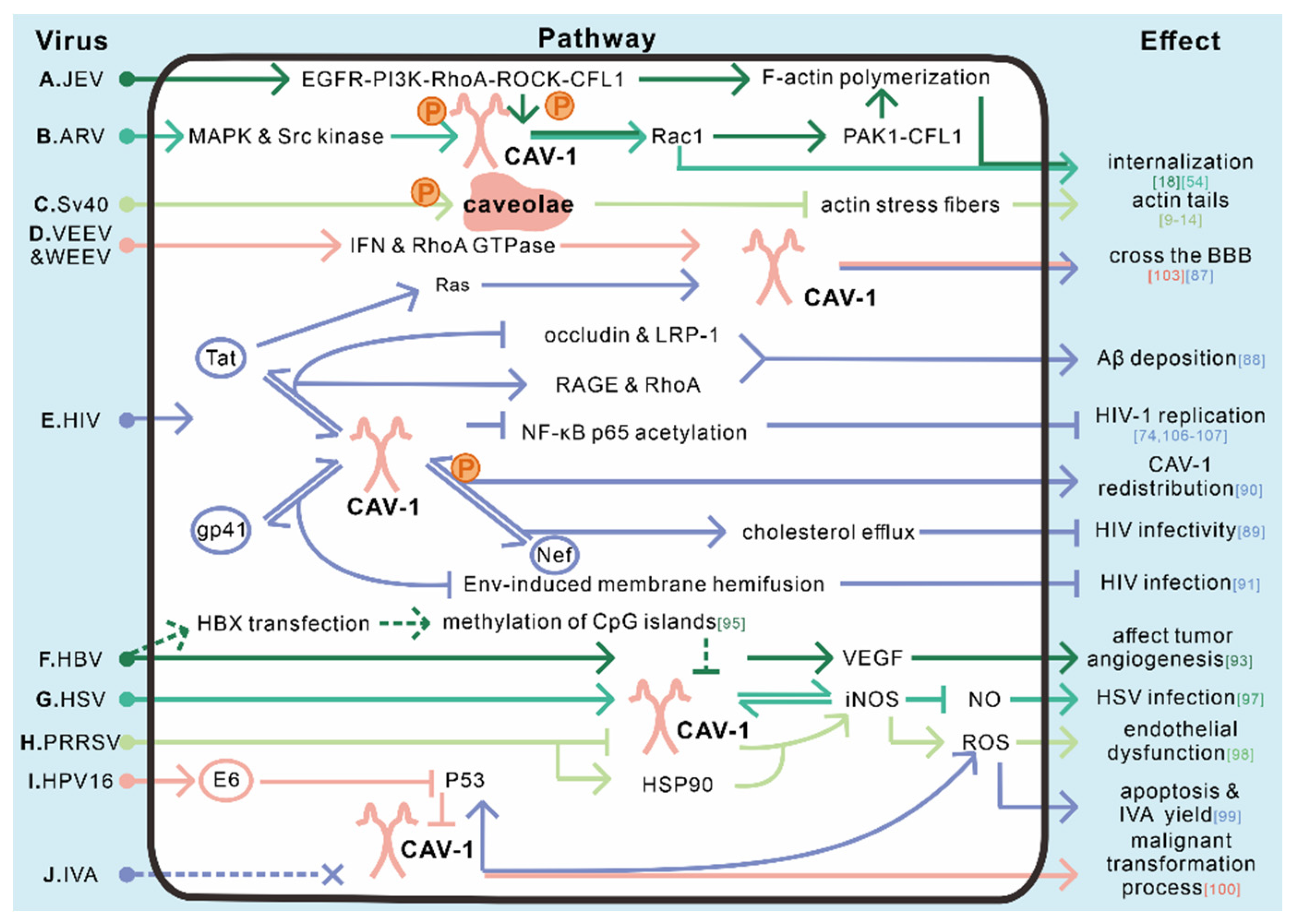
6. The Role of Caveolae/CAV-1 in Virus Infection Related Signal Transduction
6.1. The Role of Caveolae/CAV-1 in HIV Infection-Related Signal Transduction
6.2. The Role of Caveolae/CAV-1 in HBV Infection-Related Signal Transduction
6.3. The Role of Caveolae/CAV-1 in Other Viruses Infection-Related Signal Transduction
7. Conclusions and Perspective
Funding
Conflicts of Interest
Abbreviations
| AIDS | Acquired immunodeficiency syndrome |
| A-MLV | Amphotropic murine leukemia virus |
| ARV | Avian reovirus |
| BBB | Blood-brain barrier |
| BGs | Birbeck granules |
| BKV | Human polyomavirus BK |
| BPV | Bovine papillomavirus |
| CAV-1 | Caveolin-1 |
| CAV-2 | Caveolin-2 |
| CBD | Caveolin-binding domain |
| CRCoV | Canine respiratory coronavirus |
| CSD | Caveolin scaffolding domain |
| CSF | Classical swine fever |
| CSFV | Classical swine fever virus |
| DENV | Dengue virus |
| EBMECs | Equine brain microvascular endothelial cells |
| EBOV | Ebola virus |
| EBV | Epstein-Barr virus |
| EHD2 | Eps15-homology domain-containing protein 2 |
| EHV | Equine herpes virus |
| ESCRT | Endosomal sorting complex required for transport |
| EV | Echovirus |
| FMDV | Foot-and-mouth disease virus |
| GCRV | Grass carp reovirus |
| HA | Hemagglutinin |
| HAdV | Human adenoviruses |
| HBsAg | Hepatitis B surface antigen |
| HBV | Hepatitis B virus |
| HBx | HBV-encoded X protein |
| HCC | Hepatocellular carcinoma |
| HCoV-229E | Human coronavirus 229E |
| HCoV-OC43 | Human coronavirus OC43 |
| HCV | Hepatitis C virus |
| HDL | High-density lipoprotein |
| HIV | Human immunodeficiency virus |
| HPV | Human papillomaviruses |
| HRPTEC | Human renal proximal tubular epithelial cells |
| HS | Heparan sulfate |
| HSV | Herpes simplex virus |
| IBV | Infectious bronchitis virus |
| IFN | Interferon |
| iNOS | Inducible nitric oxide synthase |
| ISKN | Infection spleen and kidney necrosis |
| ISKNV | Infection spleen and kidney necrosis virus |
| IVA | Influenza A virus |
| JCV | JC virus |
| JEV | Japanese encephalitis virus |
| LCs | Langerhans cells |
| LD | Lipid droplets |
| LTNP | Long-term non-progressor |
| M | Matrix protein |
| MAPK | Mitogen-activated protein kinase |
| MARV | Marburg virus |
| MCP | Major capsid protein |
| MDRV | Muscovy duck reovirus |
| mEFs | Mouse embryo fibroblasts |
| MHC | Major histocompatibility complex |
| MPyV | Mouse polyomavirus |
| MβCD | Methyl-β-cyclodextrin |
| NDV | Newcastle disease virus |
| NP | Nucleocapsid protein |
| NSP | Nonstructural protein |
| Ny | Nystatin |
| PACSIN2 | Protein kinase C and casein kinase substrate in neurons 2 |
| PEDV | Porcine epidemic diarrhea virus |
| PIV | Parainfluenza virus |
| PMA | Phorbol-12-myristate-13-acetate |
| PPRV | Peste des petits ruminants virus |
| PRRSV | Porcine reproductive and respiratory syndrome virus |
| RNAi | RNA interference |
| ROR1 | Receptor tyrosine kinase-like orphan receptor 1 |
| RSV | Respiratory syncytial virus |
| RVFV | Rift valley fever virus |
| siRNA | Small interfering RNA |
| ST cells | Swine testis cells |
| SV40 | Simian virus 40 |
| TFV | Tiger frog virus |
| TGEV | Transmissible gastroenteritis virus |
| TGN | Trans Golgi network |
| VCP | Valosin-containing protein |
| VEEV | Venezuelan virus |
| VEGF | Vascular endothelial growth factor |
| VSV | Vesicular stomatitis virus |
| WEEV | Western equine encephalitis virus |
| WHO | World Health Organization |
References
- China.org.cn. Public Health Emergency of International Concern (The Fight Against COVID-19). Available online: http://www.china.org.cn/english/china_key_words/2020-03/06/content_75783332.htm (accessed on 6 March 2020).
- Li, S. Regulation of Ribosomal Proteins on Viral Infection. Cells 2019, 8, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Gao, W.; Li, J.; Wu, W.; Jiu, Y. The Role of Host Cytoskeleton in Flavivirus Infection. Virol. Sin. 2019, 34, 30–41. [Google Scholar] [CrossRef]
- Mercer, J.; Schelhaas, M.; Helenius, A. Virus entry by endocytosis. Annu. Rev. Biochem. 2010, 79, 803–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parton, R.G. Caveolae: Structure, Function, and Relationship to Disease. Annu. Rev. Cell. Dev. Biol. 2018, 34, 111–136. [Google Scholar] [CrossRef]
- Stan, R.V. Structure of caveolae. Biochim. Biophys. Acta 2005, 1746, 334–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, F.S.; Rodriguez, N.E.; Adesse, D.; Garzoni, L.R.; Esper, L.; Lisanti, M.P.; Burk, R.D.; Albanese, C.; Van Doorslaer, K.; Weiss, L.M.; et al. Recent developments in the interactions between caveolin and pathogens. Adv. Exp. Med. Biol. 2012, 729, 65–82. [Google Scholar] [CrossRef] [Green Version]
- Mergia, A. The Role of Caveolin 1 in HIV Infection and Pathogenesis. Viruses 2017, 9, 129. [Google Scholar] [CrossRef] [Green Version]
- Anderson, H.A.; Chen, Y.; Norkin, L.C. Bound simian virus 40 translocates to caveolin-enriched membrane domains, and its entry is inhibited by drugs that selectively disrupt caveolae. Mol. Biol. Cell. 1996, 7, 1825–1834. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Norkin, L.C. Extracellular simian virus 40 transmits a signal that promotes virus enclosure within caveolae. Exp. Cell. Res. 1999, 246, 83–90. [Google Scholar] [CrossRef]
- Norkin, L.C. Simian virus 40 infection via MHC class I molecules and caveolae. Immunol. Rev. 1999, 168, 13–22. [Google Scholar] [CrossRef]
- Pelkmans, L.; Kartenbeck, J.; Helenius, A. Caveolar endocytosis of simian virus 40 reveals a new two-step vesicular-transport pathway to the ER. Nat. Cell. Biol. 2001, 3, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Pelkmans, L.; Püntener, D.; Helenius, A. Local actin polymerization and dynamin recruitment in SV40-induced internalization of caveolae. Science 2002, 296, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Luetterforst, R.; Harding, A.; Apolloni, A.; Etheridge, M.; Stang, E.; Rolls, B.; Hancock, J.F.; Parton, R.G. Dominant-negative caveolin inhibits H-Ras function by disrupting cholesterol-rich plasma membrane domains. Nat. Cell. Biol. 1999, 1, 98–105. [Google Scholar] [CrossRef]
- Turtle, L.; Solomon, T. Japanese encephalitis—The prospects for new treatments. Nat. Rev. Neurol. 2018, 14, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.Z.; Xu, Q.Q.; Wu, D.G.; Ren, H.; Zhao, P.; Lao, W.G.; Wang, Y.; Tao, Q.Y.; Qian, X.J.; Wei, Y.H.; et al. Japanese encephalitis virus enters rat neuroblastoma cells via a pH-dependent, dynamin and caveola-mediated endocytosis pathway. J. Virol. 2012, 86, 13407–13422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Jadhav, H.R.; Bhatt, T. Dynamin Functions and Ligands: Classical Mechanisms Behind. Mol. Pharmacol. 2017, 91, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Cao, M.; Song, H.; Chen, S.; Qian, X.; Zhao, P.; Ren, H.; Tang, H.; Wang, Y.; Wei, Y.; et al. Caveolin-1-mediated Japanese encephalitis virus entry requires a two-step regulation of actin reorganization. Future Microbiol. 2016, 11, 1227–1248. [Google Scholar] [CrossRef]
- Khasa, R.; Vaidya, A.; Vrati, S.; Kalia, M. Membrane trafficking RNA interference screen identifies a crucial role of the clathrin endocytic pathway and ARP2/3 complex for Japanese encephalitis virus infection in HeLa cells. J. Gen. Virol. 2019, 100, 176–186. [Google Scholar] [CrossRef]
- Liu, C.-C.; Zhang, Y.-N.; Li, Z.-Y.; Hou, J.-X.; Zhou, J.; Kan, L.; Zhou, B.; Chen, P.-Y. Rab5 and Rab11 Are Required for Clathrin-Dependent Endocytosis of Japanese Encephalitis Virus in BHK-21 Cells. J. Virol. 2017, 91, e01113-17. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; He, M.; Liu, X.; Li, X.; Fan, B.; Zhao, S. Japanese encephalitis virus infects porcine kidney epithelial PK15 cells via clathrin- and cholesterol-dependent endocytosis. Virol. J. 2013, 10, 258. [Google Scholar] [CrossRef] [Green Version]
- Peters, M.G. Hepatitis B Virus Infection: What Is Current and New. Top. Antivir. Med. 2019, 26, 112–116. [Google Scholar] [PubMed]
- Hao, X.; Shang, X.; Wu, J.; Shan, Y.; Cai, M.; Jiang, J.; Huang, Z.; Tang, Z.; Wang, H. Single-particle tracking of hepatitis B virus-like vesicle entry into cells. Small 2011, 7, 1212–1218. [Google Scholar] [CrossRef] [PubMed]
- Macovei, A.; Radulescu, C.; Lazar, C.; Petrescu, S.; Durantel, D.; Dwek, R.A.; Zitzmann, N.; Nichita, N.B. Hepatitis B virus requires intact caveolin-1 function for productive infection in HepaRG cells. J. Virol. 2010, 84, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, R.; Kiyota, A.; Suzaki, E.; Kataoka, K.; Ohe, Y.; Miyamoto, K.; Senda, T.; Fujimoto, T. Human coronavirus 229E binds to CD13 in rafts and enters the cell through caveolae. J. Virol. 2004, 78, 8701–8708. [Google Scholar] [CrossRef] [Green Version]
- Owczarek, K.; Szczepanski, A.; Milewska, A.; Baster, Z.; Rajfur, Z.; Sarna, M.; Pyrc, K. Early events during human coronavirus OC43 entry to the cell. Sci. Rep. 2018, 8, 7124. [Google Scholar] [CrossRef]
- Werling, D.; Hope, J.C.; Chaplin, P.; Collins, R.A.; Taylor, G.; Howard, C.J. Involvement of caveolae in the uptake of respiratory syncytial virus antigen by dendritic cells. J. Leukoc. Biol. 1999, 66, 50–58. [Google Scholar] [CrossRef]
- Empig, C.J.; Goldsmith, M.A. Association of the caveola vesicular system with cellular entry by filoviruses. J. Virol. 2002, 76, 5266–5270. [Google Scholar] [CrossRef] [Green Version]
- Harmon, B.; Schudel, B.R.; Maar, D.; Kozina, C.; Ikegami, T.; Tseng, C.T.; Negrete, O.A. Rift Valley fever virus strain MP-12 enters mammalian host cells via caveola-mediated endocytosis. J. Virol. 2012, 86, 12954–12970. [Google Scholar] [CrossRef] [Green Version]
- Hasebe, R.; Sasaki, M.; Sawa, H.; Wada, R.; Umemura, T.; Kimura, T. Infectious entry of equine herpesvirus-1 into host cells through different endocytic pathways. Virology 2009, 393, 198–209. [Google Scholar] [CrossRef] [Green Version]
- Ning, P.; Gao, L.; Zhou, Y.; Hu, C.; Lin, Z.; Gong, C.; Guo, K.; Zhang, X. Caveolin-1-mediated endocytic pathway is involved in classical swine fever virus Shimen infection of porcine alveolar macrophages. Vet. Microbiol. 2016, 195, 81–86. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Wang, S.; Liu, F. Dynamics of transmissible gastroenteritis virus internalization unraveled by single-virus tracking in live cells. FASEB J. 2020, 34, 4653–4669. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; She, G.; Wu, T.; Xue, C.; Cao, Y. PEDV enters cells through clathrin-, caveolae-, and lipid raft-mediated endocytosis and traffics via the endo-/lysosome pathway. Vet. Res. 2020, 51, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.J.; Liu, D.; Wu, Y.Y.; Yang, X.B.; Yang, L.S.; Mi, S.; Huang, Y.X.; Luo, Y.W.; Jia, K.T.; Liu, Z.Y.; et al. Entry of tiger frog virus (an Iridovirus) into HepG2 cells via a pH-dependent, atypical, caveola-mediated endocytosis pathway. J. Virol. 2011, 85, 6416–6426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.J.; Wu, Y.Y.; Yang, L.S.; Yang, X.B.; He, J.; Mi, S.; Jia, K.T.; Weng, S.P.; Yu, X.Q.; He, J.G. Infectious spleen and kidney necrosis virus (a fish iridovirus) enters Mandarin fish fry cells via caveola-dependent endocytosis. J. Virol. 2012, 86, 2621–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.J.; Yang, X.B.; Wu, Y.Y.; Yang, L.S.; Mi, S.; Liu, Z.Y.; Jia, K.T.; Huang, Y.X.; Weng, S.P.; Yu, X.Q.; et al. Involvement of caveolin-1 in the Jak-Stat signaling pathway and infectious spleen and kidney necrosis virus infection in mandarin fish (Siniperca chuatsi). Mol. Immunol. 2011, 48, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.T.; Wu, Y.Y.; Liu, Z.Y.; Mi, S.; Zheng, Y.W.; He, J.; Weng, S.P.; Li, S.C.; He, J.G.; Guo, C.J. Mandarin fish caveolin 1 interaction with major capsid protein of infectious spleen and kidney necrosis virus and its role in early stages of infection. J. Virol. 2013, 87, 3027–3038. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Huang, M.; Yuan, Q.; Wei, Y.; Gao, Y.; Mao, L.; Gu, L.; Tan, Y.W.; Zhong, Y.; Liu, D.; et al. The Important Role of Lipid Raft-Mediated Attachment in the Infection of Cultured Cells by Coronavirus Infectious Bronchitis Virus Beaudette Strain. PLoS ONE 2017, 12, e0170123. [Google Scholar] [CrossRef] [Green Version]
- Szczepanski, A.; Owczarek, K.; Milewska, A.; Baster, Z.; Rajfur, Z.; Mitchell, J.A.; Pyrc, K. Canine respiratory coronavirus employs caveolin-1-mediated pathway for internalization to HRT-18G cells. Vet. Res. 2018, 49, 55. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Qi, X.; Guo, H.; Jia, P.; Chen, S.; Chen, Z.; Wang, T.; Wang, J.; Xue, Q. Peste des Petits Ruminants Virus Enters Caprine Endometrial Epithelial Cells via the Caveolae-Mediated Endocytosis Pathway. Front. Microbiol. 2018, 9, 210. [Google Scholar] [CrossRef]
- Beer, C.; Andersen, D.S.; Rojek, A.; Pedersen, L. Caveola-dependent endocytic entry of amphotropic murine leukemia virus. J. Virol. 2005, 79, 10776–10787. [Google Scholar] [CrossRef] [Green Version]
- Yousuf, M.A.; Zhou, X.; Mukherjee, S.; Chintakuntlawar, A.V.; Lee, J.Y.; Ramke, M.; Chodosh, J.; Rajaiya, J. Caveolin-1 associated adenovirus entry into human corneal cells. PLoS ONE 2013, 8, e77462. [Google Scholar] [CrossRef] [PubMed]
- Colin, M.; Mailly, L.; Rogée, S.; D’Halluin, J.-C. Efficient species C HAdV infectivity in plasmocytic cell lines using a clathrin-independent lipid raft/caveola endocytic route. Mol. Ther. 2005, 11, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Eash, S.; Querbes, W.; Atwood, W.J. Infection of vero cells by BK virus is dependent on caveolae. J. Virol. 2004, 78, 11583–11590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriyama, T.; Marquez, J.P.; Wakatsuki, T.; Sorokin, A. Caveolar endocytosis is critical for BK virus infection of human renal proximal tubular epithelial cells. J. Virol. 2007, 81, 8552–8562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bousarghin, L.; Touzé, A.; Sizaret, P.-Y.; Coursaget, P. Human papillomavirus types 16, 31, and 58 use different endocytosis pathways to enter cells. J. Virol. 2003, 77, 3846–3850. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.L.; Campos, S.K.; Ozbun, M.A. Human papillomavirus type 31 uses a caveolin 1- and dynamin 2-mediated entry pathway for infection of human keratinocytes. J. Virol. 2007, 81, 9922–9931. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.L.; Campos, S.K.; Wandinger-Ness, A.; Ozbun, M.A. Caveolin-1-dependent infectious entry of human papillomavirus type 31 in human keratinocytes proceeds to the endosomal pathway for pH-dependent uncoating. J. Virol. 2008, 82, 9505–9512. [Google Scholar] [CrossRef] [Green Version]
- Marjomaki, V.; Pietiainen, V.; Matilainen, H.; Upla, P.; Ivaska, J.; Nissinen, L.; Reunanen, H.; Huttunen, P.; Hyypia, T.; Heino, J. Internalization of echovirus 1 in caveolae. J. Virol. 2002, 76, 1856–1865. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.Y.; Yang, Y.T.; Yu, S.L.; Hsiao, K.N.; Liu, C.C.; Sia, C.; Chow, Y.H. Caveolar endocytosis is required for human PSGL-1-mediated enterovirus 71 infection. J. Virol. 2013, 87, 9064–9076. [Google Scholar] [CrossRef] [Green Version]
- Schulz, W.L.; Haj, A.K.; Schiff, L.A. Reovirus uses multiple endocytic pathways for cell entry. J. Virol. 2012, 86, 12665–12675. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, V.; Larocco, M.; Baxt, B. Heparan sulfate-binding foot-and-mouth disease virus enters cells via caveola-mediated endocytosis. J. Virol. 2008, 82, 9075–9085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richterova, Z.; Liebl, D.; Horak, M.; Palkova, Z.; Stokrova, J.; Hozak, P.; Korb, J.; Forstova, J. Caveolae are involved in the trafficking of mouse polyomavirus virions and artificial VP1 pseudocapsids toward cell nuclei. J. Virol. 2001, 75, 10880–10891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.R.; Wang, Y.C.; Chi, P.I.; Wang, L.; Wang, C.Y.; Lin, C.H.; Liu, H.J. Cell entry of avian reovirus follows a caveolin-1-mediated and dynamin-2-dependent endocytic pathway that requires activation of p38 mitogen-activated protein kinase (MAPK) and Src signaling pathways as well as microtubules and small GTPase Rab5 protein. J. Biol. Chem. 2011, 286, 30780–30794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Yan, P.; Liu, Z.; Cai, D.; Luo, Y.; Wu, X.; Wang, Q.; Huang, Y.; Wu, Y. Muscovy duck reovirus enters susceptible cells via a caveolae-mediated endocytosis-like pathway. Virus Res. 2020, 276, 197806. [Google Scholar] [CrossRef]
- Zhang, F.; Guo, H.; Zhang, J.; Chen, Q.; Fang, Q. Identification of the caveolae/raft-mediated endocytosis as the primary entry pathway for aquareovirus. Virology 2018, 513, 195–207. [Google Scholar] [CrossRef]
- Cai, Q.C.; Jiang, Q.W.; Zhao, G.M.; Guo, Q.; Cao, G.W.; Chen, T. Putative caveolin-binding sites in SARS-CoV proteins. Acta Pharmacol. Sin. 2003, 24, 1051–1059. [Google Scholar]
- Couet, J.; Li, S.; Okamoto, T.; Ikezu, T.; Lisanti, M.P. Identification of peptide and protein ligands for the caveolin-scaffolding domain. Implications for the interaction of caveolin with caveolae-associated proteins. J. Biol. Chem. 1997, 272, 6525–6533. [Google Scholar] [CrossRef] [Green Version]
- Inoue, Y.; Tanaka, N.; Tanaka, Y.; Inoue, S.; Morita, K.; Zhuang, M.; Hattori, T.; Sugamura, K. Clathrin-dependent entry of severe acute respiratory syndrome coronavirus into target cells expressing ACE2 with the cytoplasmic tail deleted. J. Virol. 2007, 81, 8722–8729. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yang, P.; Liu, K.; Guo, F.; Zhang, Y.; Zhang, G.; Jiang, C. SARS coronavirus entry into host cells through a novel clathrin-and caveolae-independent endocytic pathway. Cell. Res. 2008, 18, 290–301. [Google Scholar] [CrossRef] [Green Version]
- Byrne, D.P.; Dart, C.; Rigden, D.J. Evaluating caveolin interactions: Do proteins interact with the caveolin scaffolding domain through a widespread aromatic residue-rich motif? PLoS ONE 2012, 7, e44879. [Google Scholar] [CrossRef]
- Salata, C.; Calistri, A.; Alvisi, G.; Celestino, M.; Parolin, C.; Palù, G. Ebola Virus Entry: From Molecular Characterization to Drug Discovery. Viruses 2019, 11, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oladunni, F.S.; Horohov, D.W.; Chambers, T.M. EHV-1: A Constant Threat to the Horse Industry. Front. Microbiol. 2019, 10, 2668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, O.; Boucke, K.; Hammer, S.V.; Keller, S.; Stidwill, R.P.; Hemmi, S.; Greber, U.F. Adenovirus triggers macropinocytosis and endosomal leakage together with its clathrin-mediated uptake. J. Cell. Biol. 2002, 158, 1119–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, O.; Gastaldelli, M.; Boucke, K.; Hemmi, S.; Greber, U.F. Early steps of clathrin-mediated endocytosis involved in phagosomal escape of Fcgamma receptor-targeted adenovirus. J. Virol. 2005, 79, 2604–2613. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Wang, Y.; Shang, Y.; Zhang, Z.; Liu, X. How foot-and-mouth disease virus receptor mediates foot-and-mouth disease virus infection. Virol. J. 2015, 12, 9. [Google Scholar] [CrossRef] [Green Version]
- Berryman, S.; Clark, S.; Monaghan, P.; Jackson, T. Early events in integrin alphavbeta6-mediated cell entry of foot-and-mouth disease virus. J. Virol. 2005, 79, 8519–8534. [Google Scholar] [CrossRef] [Green Version]
- Pietiäinen, V.M.; Marjomäki, V.; Heino, J.; Hyypiä, T. Viral entry, lipid rafts and caveosomes. Ann. Med. 2005, 37, 394–403. [Google Scholar] [CrossRef]
- Damm, E.M.; Pelkmans, L.; Kartenbeck, J.; Mezzacasa, A.; Kurzchalia, T.; Helenius, A. Clathrin-and caveolin-1-independent endocytosis: Entry of simian virus 40 into cells devoid of caveolae. J. Cell. Biol. 2005, 168, 477–488. [Google Scholar] [CrossRef]
- Rogée, S.; Grellier, E.; Bernard, C.; Loyens, A.; Beauvillain, J.C.; D’Halluin J, C.; Colin, M. Intracellular trafficking of a fiber-modified adenovirus using lipid raft/caveolae endocytosis. Mol. Ther. 2007, 15, 1963–1972. [Google Scholar] [CrossRef]
- Laniosz, V.; Dabydeen, S.A.; Havens, M.A.; Meneses, P.I. Human papillomavirus type 16 infection of human keratinocytes requires clathrin and caveolin-1 and is brefeldin a sensitive. J. Virol. 2009, 83, 8221–8232. [Google Scholar] [CrossRef] [Green Version]
- Laniosz, V.; Holthusen, K.A.; Meneses, P.I. Bovine papillomavirus type 1: From clathrin to caveolin. J. Virol. 2008, 82, 6288–6298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Querbes, W.; O’Hara, B.A.; Williams, G.; Atwood, W.J. Invasion of host cells by JC virus identifies a novel role for caveolae in endosomal sorting of noncaveolar ligands. J. Virol. 2006, 80, 9402–9413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez Boulan, E.; Pendergast, M. Polarized distribution of viral envelope proteins in the plasma membrane of infected epithelial cells. Cell 1980, 20, 45–54. [Google Scholar] [CrossRef]
- Scheiffele, P.; Verkade, P.; Fra, A.M.; Virta, H.; Simons, K.; Ikonen, E. Caveolin-1 and -2 in the exocytic pathway of MDCK cells. J. Cell. Biol. 1998, 140, 795–806. [Google Scholar] [CrossRef]
- Tugizov, S.M.; Herrera, R.; Palefsky, J.M. Epstein-Barr virus transcytosis through polarized oral epithelial cells. J. Virol. 2013, 87, 8179–8194. [Google Scholar] [CrossRef] [Green Version]
- García Cordero, J.; León Juárez, M.; González, Y.M.J.A.; Cedillo Barrón, L.; Gutiérrez Castañeda, B. Caveolin-1 in lipid rafts interacts with dengue virus NS3 during polyprotein processing and replication in HMEC-1 cells. PLoS ONE 2014, 9, e90704. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Wang, L.; Lee, J.; Ou, J.J. Hepatitis C Virus Induces the Localization of Lipid Rafts to Autophagosomes for Its RNA Replication. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Hemgård, G.V.; Susanto, S.A.; Wirth, M. Caveolin-1 influences human influenza A virus (H1N1) multiplication in cell culture. Virol. J. 2010, 7, 108. [Google Scholar] [CrossRef] [Green Version]
- Simmons, G.E., Jr.; Taylor, H.E.; Hildreth, J.E. Caveolin-1 suppresses human immunodeficiency virus-1 replication by inhibiting acetylation of NF-κB. Virology 2012, 432, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Beer, C.; Koester, M.; Wirth, M. Caveolin-1 interacts with the Gag precursor of murine leukaemia virus and modulates virus production. Virol. J. 2006, 3, 73. [Google Scholar] [CrossRef] [Green Version]
- Alcalá, A.C.; Hernández-Bravo, R.; Medina, F.; Coll, D.S.; Zambrano, J.L.; Del Angel, R.M.; Ludert, J.E. The dengue virus non-structural protein 1 (NS1) is secreted from infected mosquito cells via a non-classical caveolin-1-dependent pathway. J. Gen. Virol. 2017, 98, 2088–2099. [Google Scholar] [CrossRef]
- Rosales Ramirez, R.; Ludert, J.E. The Dengue Virus Nonstructural Protein 1 (NS1) Is Secreted from Mosquito Cells in Association with the Intracellular Cholesterol Transporter Chaperone Caveolin Complex. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, G.; Aitken, J.; Rixon, H.W.M.; Sugrue, R.J. Caveolin-1 is incorporated into mature respiratory syncytial virus particles during virus assembly on the surface of virus-infected cells. J. Gen. Virol. 2002, 83, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Kipper, S.; Hamad, S.; Caly, L.; Avrahami, D.; Bacharach, E.; Jans, D.A.; Gerber, D.; Bajorek, M. New host factors important for respiratory syncytial virus (RSV) replication revealed by a novel microfluidics screen for interactors of matrix (M) protein. Mol. Cell. Proteom. MCP 2015, 14, 532–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, A.; Nguyen, T.H.; Leong, D.; Ravi, L.I.; Tan, B.H.; Sandin, S.; Sugrue, R.J. Caveolae provide a specialized membrane environment for respiratory syncytial virus assembly. J. Cell. Sci. 2017, 130, 1037–1050. [Google Scholar] [CrossRef] [Green Version]
- Ravid, D.; Leser, G.P.; Lamb, R.A. A role for caveolin 1 in assembly and budding of the paramyxovirus parainfluenza virus 5. J. Virol. 2010, 84, 9749–9759. [Google Scholar] [CrossRef] [Green Version]
- Laliberte, J.P.; McGinnes, L.W.; Peeples, M.E.; Morrison, T.G. Integrity of membrane lipid rafts is necessary for the ordered assembly and release of infectious Newcastle disease virus particles. J. Virol. 2006, 80, 10652–10662. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Zheng, Y.W.; Lin, Y.F.; Mi, S.; Qin, X.W.; Weng, S.P.; He, J.G.; Guo, C.J. Caveolae Restrict Tiger Frog Virus Release in HepG2 cells and Caveolae-Associated Proteins Incorporated into Virus Particles. Sci. Rep. 2016, 6, 21663. [Google Scholar] [CrossRef] [Green Version]
- Llano, M.; Kelly, T.; Vanegas, M.; Peretz, M.; Peterson, T.E.; Simari, R.D.; Poeschla, E.M. Blockade of human immunodeficiency virus type 1 expression by caveolin-1. J. Virol. 2002, 76, 9152–9164. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, L.M.; Ribeiro, C.M.; Zijlstra-Willems, E.M.; de Witte, L.; Fluitsma, D.; Tigchelaar, W.; Everts, V.; Geijtenbeek, T.B. Caveolin-1 mediated uptake via langerin restricts HIV-1 infection in human Langerhans cells. Retrovirology 2014, 11, 123. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Wang, X.M.; Nadeau, P.E.; Mergia, A. HIV infection upregulates caveolin 1 expression to restrict virus production. J. Virol. 2010, 84, 9487–9496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Smart, E.J.; Weksler, B.; Couraud, P.O.; Hennig, B.; Toborek, M. Caveolin-1 regulates human immunodeficiency virus-1 Tat-induced alterations of tight junction protein expression via modulation of the Ras signaling. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 7788–7796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, M.; Huang, W.; Jiang, W.; Wu, Y.; Chen, Q. Role of Cav-1 in HIV-1 Tat-Induced Dysfunction of Tight Junctions and Aβ-Transferring Proteins. Oxidative Med. Cell. Longev. 2019, 2019, 3403206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.; Nadeau, P.E.; Wang, X.; Mergia, A. Caveolin-1 reduces HIV-1 infectivity by restoration of HIV Nef mediated impairment of cholesterol efflux by apoA-I. Retrovirology 2012, 9, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.; Nadeau, P.E.; Mergia, A. HIV inhibits endothelial reverse cholesterol transport through impacting subcellular Caveolin-1 trafficking. Retrovirology 2015, 12, 62. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.M.; Nadeau, P.E.; Lo, Y.T.; Mergia, A. Caveolin-1 modulates HIV-1 envelope-induced bystander apoptosis through gp41. J. Virol. 2010, 84, 6515–6526. [Google Scholar] [CrossRef] [Green Version]
- Díez-Fuertes, F.; De La Torre-Tarazona, H.E.; Calonge, E.; Pernas, M.; Bermejo, M.; García-Pérez, J.; Álvarez, A.; Capa, L.; García-García, F.; Saumoy, M.; et al. Association of a single nucleotide polymorphism in the ubxn6 gene with long-term non-progression phenotype in HIV-positive individuals. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2020, 26, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Pan, Y.; Yao, Y.; Zhu, Z.; Chen, J.; Sun, X.; Qiu, Y.; Ding, Y. Expression and significance of caveolin-1 in hepatitis B virus-associated hepatocellular carcinoma. Exp. Ther. Med. 2017, 14, 4356–4362. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xue, F.; Chen, S.; Zhang, D.; Lu, C.; Tang, G. The influence of caveolin-1 gene polymorphisms on hepatitis B virus-related hepatocellular carcinoma susceptibility in Chinese Han population: A case-control study. Medicine 2017, 96, e7359. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Lu, Q.; Dong, J.; Li, X.; Ma, K.; Cai, L. Hepatitis B virus X protein suppresses caveolin-1 expression in hepatocellular carcinoma by regulating DNA methylation. BMC Cancer 2012, 12, 353. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Tey, S.K.; Ko, F.C.F.; Kwong, E.M.L.; Gao, Y.; Ng, I.O.; Cheung, S.T.; Guan, X.Y.; Yam, J.W.P. C-terminal truncated HBx protein activates caveolin-1/LRP6/β-catenin/FRMD5 axis in promoting hepatocarcinogenesis. Cancer Lett. 2019, 444, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Geng, S.; Bi, Y.; Liu, H.; Hu, Y.; Li, X.; Zhang, Y.; Zhou, X.; Zheng, G.; He, B.; et al. Herpes Simplex Virus 1 Suppresses the Function of Lung Dendritic Cells via Caveolin-1. Clin. Vaccine Immunol. CVI 2015, 22, 883–895. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.; Hou, M.; Liu, J.; Zhang, S.; Liu, B.; Wu, X.; Liu, G. Regulation of iNOS-Derived ROS Generation by HSP90 and Cav-1 in Porcine Reproductive and Respiratory Syndrome Virus-Infected Swine Lung Injury. Inflammation 2017, 40, 1236–1244. [Google Scholar] [CrossRef]
- Bohm, K.; Sun, L.; Thakor, D.; Wirth, M. Caveolin-1 limits human influenza A virus (H1N1) propagation in mouse embryo-derived fibroblasts. Virology 2014, 462–463, 241–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razani, B.; Altschuler, Y.; Zhu, L.; Pestell, R.G.; Mostov, K.E.; Lisanti, M.P. Caveolin-1 expression is down-regulated in cells transformed by the human papilloma virus in a p53-dependent manner. Replacement of caveolin-1 expression suppresses HPV-mediated cell transformation. Biochemistry 2000, 39, 13916–13924. [Google Scholar] [CrossRef] [PubMed]
- Mir, K.D.; Parr, R.D.; Schroeder, F.; Ball, J.M. Rotavirus NSP4 interacts with both the amino- and carboxyl-termini of caveolin-1. Virus Res. 2007, 126, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Parr, R.D.; Storey, S.M.; Mitchell, D.M.; McIntosh, A.L.; Zhou, M.; Mir, K.D.; Ball, J.M. The rotavirus enterotoxin NSP4 directly interacts with the caveolar structural protein caveolin-1. J. Virol. 2006, 80, 2842–2854. [Google Scholar] [CrossRef] [Green Version]
- Salimi, H.; Cain, M.D.; Jiang, X.; Roth, R.A.; Beatty, W.L.; Sun, C.; Klimstra, W.B.; Hou, J.; Klein, R.S. Encephalitic Alphaviruses Exploit Caveola-Mediated Transcytosis at the Blood-Brain Barrier for Central Nervous System Entry. mBio 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Chazal, N.; Gerlier, D. Virus entry, assembly, budding, and membrane rafts. Microbiol. Mol. Biol. Rev. MMBR 2003, 67, 226–237. [Google Scholar] [CrossRef] [Green Version]
- Garoff, H.; Hewson, R.; Opstelten, D.J. Virus maturation by budding. Microbiol. Mol. Biol. Rev. MMBR 1998, 62, 1171–1190. [Google Scholar] [CrossRef] [Green Version]
- Humphries, A.C.; Way, M. The non-canonical roles of clathrin and actin in pathogen internalization, egress and spread. Nat. Rev. Microbiol. 2013, 11, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Votteler, J.; Sundquist, W.I. Virus budding and the ESCRT pathway. Cell Host Microbe 2013, 14, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boscher, C.; Nabi, I.R. Caveolin-1: Role in cell signaling. Adv. Exp. Med. Biol. 2012, 729, 29–50. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. Caveolin-1 and Vascular Dysfunction. Available online: https://www.clinicaltrials.gov/ct2/show/NCT01426529?term=caveolin&draw=2&rank=1 (accessed on 31 August 2011).
- ClinicalTrials.gov. Safety, Tolerability and Pharmacokinetic Study of LTI-03 in Healthy Adult Subjects. Available online: https://www.clinicaltrials.gov/ct2/show/NCT04233814?term=caveolin&draw=2&rank=2 (accessed on 18 January 2020).
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.R. HLA class I antigen serves as a receptor for human coronavirus OC43. Immunol. Investig. 1993, 22, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Krempl, C.; Schultze, B.; Herrler, G. Analysis of cellular receptors for human coronavirus OC43. Adv. Exp. Med. Biol. 1995, 380, 371–374. [Google Scholar] [CrossRef] [Green Version]
- Yeager, C.L.; Ashmun, R.A.; Williams, R.K.; Cardellichio, C.B.; Shapiro, L.H.; Look, A.T.; Holmes, K.V. Human aminopeptidase N is a receptor for human coronavirus 229E. Nature 1992, 357, 420–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]



| Viral Life Cycle Steps | Viruses | Description | References |
|---|---|---|---|
| Intracellular trafficking | SV40 | Go through a two-step transport route from plasma membrane caveolae to the ER through caveosomes | [12] |
| ISKNV | [35] | ||
| MPyV | Trafficking through the CAV-1-containing monopinocytic vesicles and the involvement of the cytoskeletal system | [53] | |
| HAdV2/BAdV4 | Induces the formation of mega-caveosomes, and subsequent trafficking does not require endosomes | [70] | |
| HPV16 | Entry via clathrin-mediated endocytosis, and shuttle from endosome pathway to caveolae route and then on to ER | [71] | |
| BPV1 | [72] | ||
| JCV | [73] | ||
| FMDV | Enters cells via caveolar pathway and then shuttles towards endosomes | [52] | |
| HPV31 | [48] | ||
| PEDV | Enters cell through both clathrin- and caveolae-mediated way, and then participates in the endosome pathway | [33] | |
| VSV | VSV-G is transferred from TGN to the basolateral surface by vesicles consisting of hetero-oligomers of CAV-1 and CAV-2 | [75] | |
| Influenza virus | HA is transported from TGN to the apical cell surface by vesicles consisting of large homo-oligomers of CAV-1 | [75] | |
| EBV | Transports from basolateral to apical by CAV-mediated virus entry | [76] | |
| Replication | DENV | Caveolar microdomains play a role in DENV polyprotein processing and replication | [77] |
| HCV | HCV infection induces autophagy to enhance its own replication, and CAV-1 is found on autophagosomes | [78] | |
| IVA | Interaction between CAV-1 and M2 matrix protein affects H1N1 replication | [79] | |
| HIV-1 | CAV-1 could inhibit the expression of HIV-1 protein | [80] | |
| Assembly and egress | MLV | CAV-1 interacts with Gag, which could drive MLV assembly | [81] |
| DENV NS1 | CAV-1 assists DENV NS1 release from mosquito cells without passing through the Golgi apparatus | [82,83] | |
| RSV | Caveolar morphology changed; CAV-1 is incorporated into the envelope | [84,85,86] | |
| PIV5 | CAV-1 triggers the assembly and/or budding of virus | [87] | |
| NDV | Purified NDV particles contain lipid raft-associated CAV-1 | [88] | |
| TFV | CAV-1 has an inhibitory effect on the egress of TFV virions | [89] | |
| Related signaling pathway | HIV | CAV-1 inhibits the production of proviral DNA | [90] |
| CAV-1 inhibits HIV through lysosomal degradation pathway | [91] | ||
| Tat induces CAV-1 upregulation | [92] | ||
| HIV could cross the BBB owning to CAV-1-needed Tat-induced Ras signal activation | [93] | ||
| Tat interacts with CAV-1, causing Aβ deposit in brain | [94] | ||
| CAV-1 interacts with Nef, activating cholesterol efflux | [95] | ||
| Nef affects the redistribution of CAV-1, thereby preventing cholesterol from transferring to HDL | [96] | ||
| CAV-1 interacts with gp41 and inhibits Env-induced membrane hemifusion | [97] | ||
| CAV-1 is involved in the mutation of UBXN6, which can affect HIV replication | [98] | ||
| HBV | CAV-1 associates with HCC and participates in VEGF-mediated signaling | [99,100] | |
| HBx can downregulate CAV-1 transcription in HCC cell line | [101] | ||
| HBxΔC can upregulate CAV-1 transcription in HCC cell line | [102] | ||
| HSV-1 | CAV-1 is utilized by HSV to escape iNOS-mediated responses | [103] | |
| PRRSV | CAV-1 and HSP90 induce an increased ROS generation and higher endothelial dysfunction | [104] | |
| IVA | The absence of CAV-1 leads to an upregulation of p53 and ROS level | [105] | |
| HPV16 | Viral protein E6 downregulates CAV-1 level by inactivating p53 | [106] | |
| Rotavirus | Rotavirus NSP 4 associates with CAV-1, leading NSP4 intracellular trafficking from the ER to the cell surface | [107,108] | |
| VEEV and WEEV | IFN and RhoA GTPase signaling pathways are involved in CAV-1-mediated transcytosis | [109] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, Y.; Wen, Z.; Gao, W.; Lin, Z.; Zhong, J.; Jiu, Y. Multifaceted Functions of Host Cell Caveolae/Caveolin-1 in Virus Infections. Viruses 2020, 12, 487. https://doi.org/10.3390/v12050487
Xing Y, Wen Z, Gao W, Lin Z, Zhong J, Jiu Y. Multifaceted Functions of Host Cell Caveolae/Caveolin-1 in Virus Infections. Viruses. 2020; 12(5):487. https://doi.org/10.3390/v12050487
Chicago/Turabian StyleXing, Yifan, Zeyu Wen, Wei Gao, Zhekai Lin, Jin Zhong, and Yaming Jiu. 2020. "Multifaceted Functions of Host Cell Caveolae/Caveolin-1 in Virus Infections" Viruses 12, no. 5: 487. https://doi.org/10.3390/v12050487
APA StyleXing, Y., Wen, Z., Gao, W., Lin, Z., Zhong, J., & Jiu, Y. (2020). Multifaceted Functions of Host Cell Caveolae/Caveolin-1 in Virus Infections. Viruses, 12(5), 487. https://doi.org/10.3390/v12050487