Isolation and Characterization of Bacteriophages That Infect Citrobacter rodentium, a Model Pathogen for Intestinal Diseases

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culturing
2.2. Phage Isolation, Cultivation and Purification
2.3. Transmission Electron Microscopy
2.4. DNA Extraction and Genome Sequencing
2.5. Genome Annotations, Alignments, and Phylogenetic Analyses
2.6. Phage Adsorption and Growth
2.7. Phage Lysis Curves
2.8. Phage Host Range
2.9. Statistics
2.10. Accession Numbers
3. Results
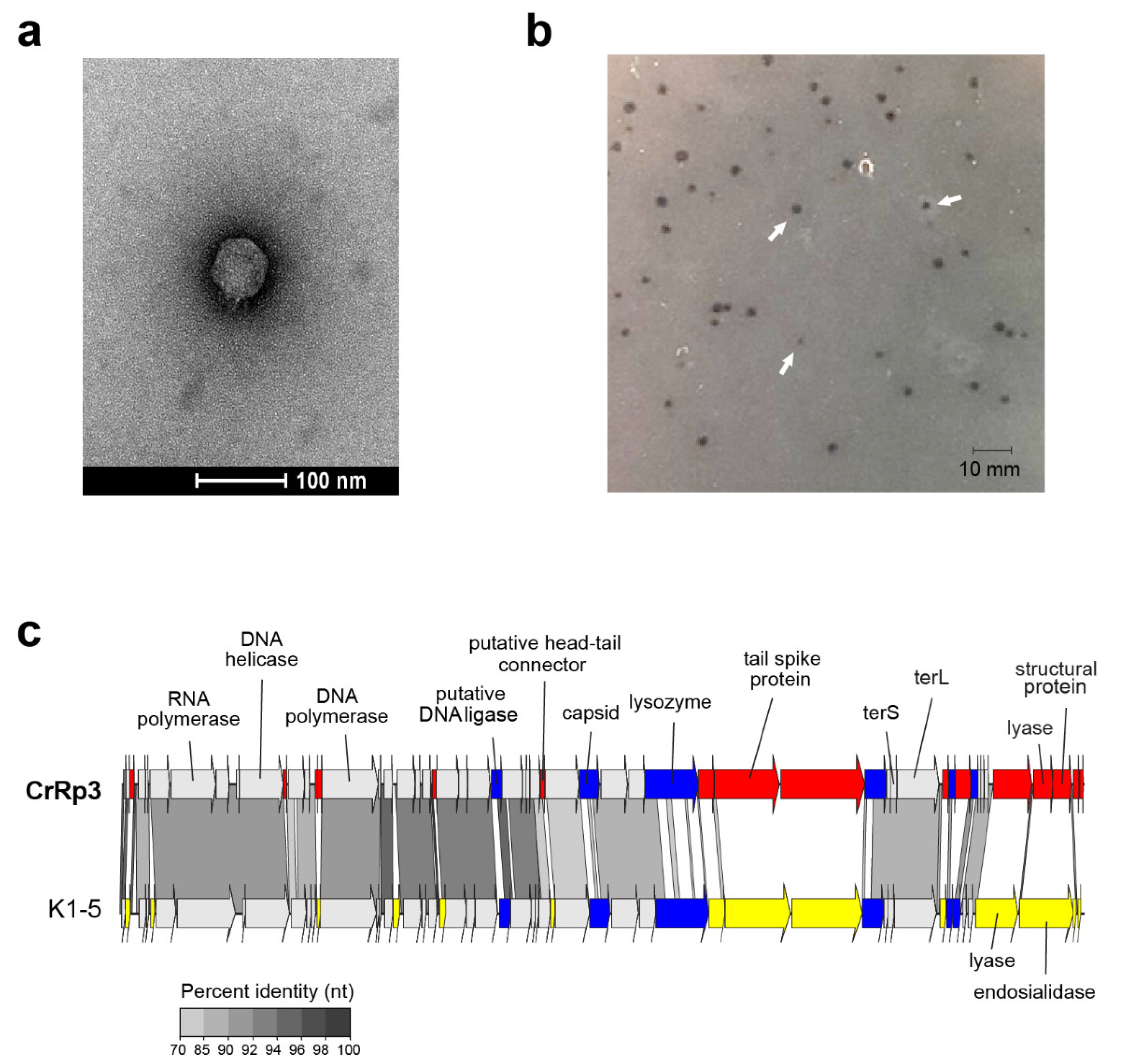
3.1. Isolation, Morphology, Sequencing, and Genome Annotation of CrRp3
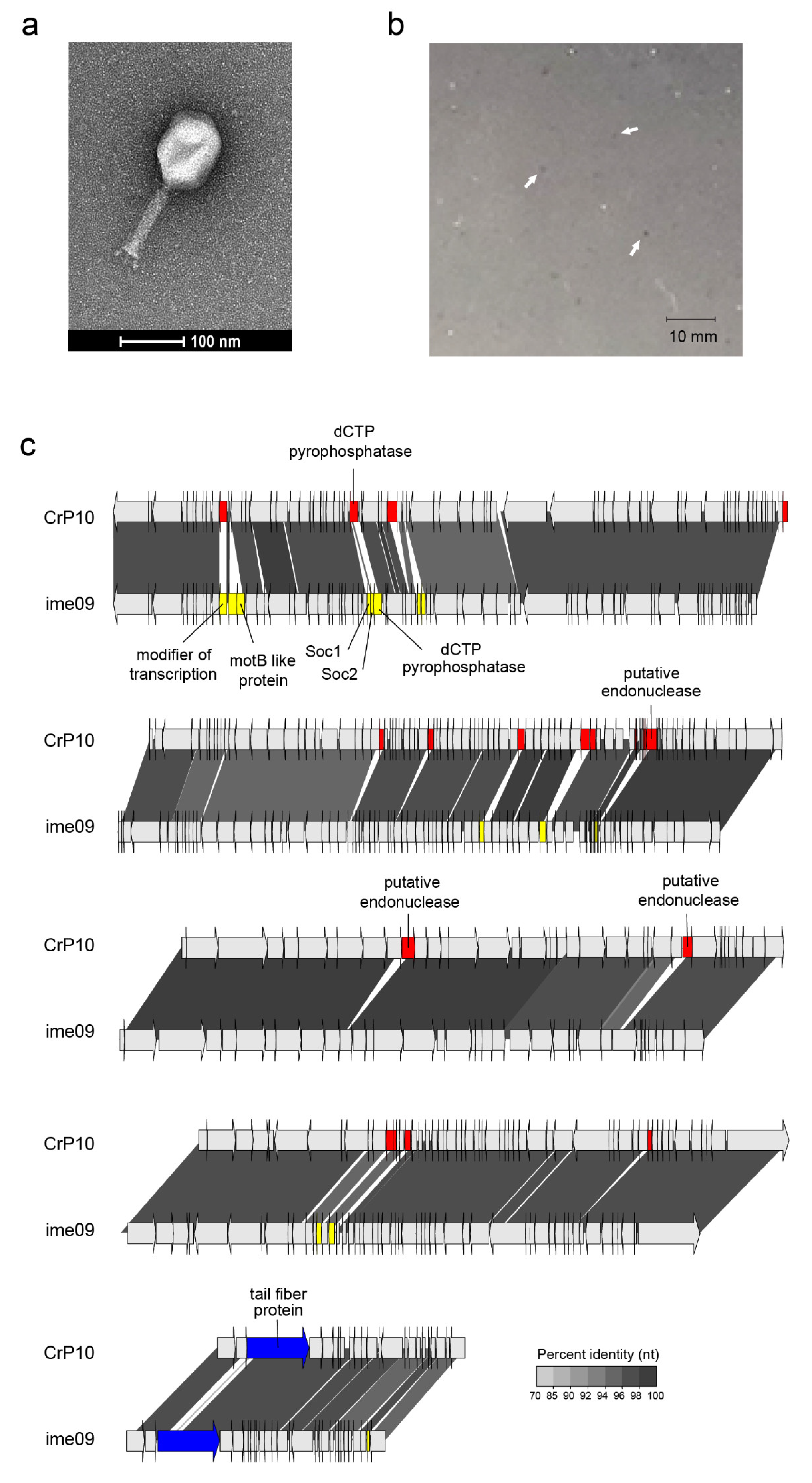
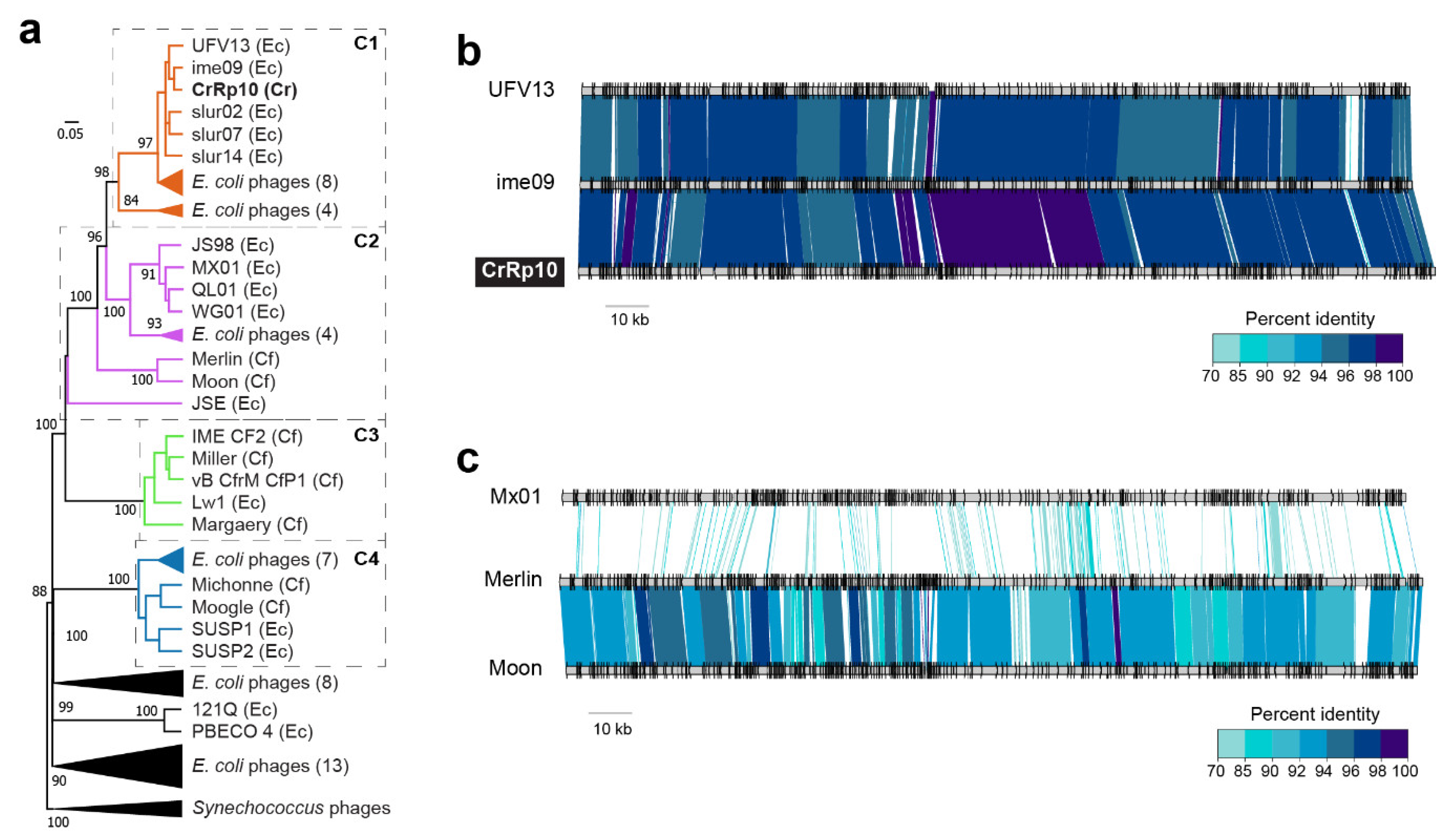
3.2. Morphology and Genome of Phage CrRp10
3.3. Comparison of Citrobacter Phage Proteins
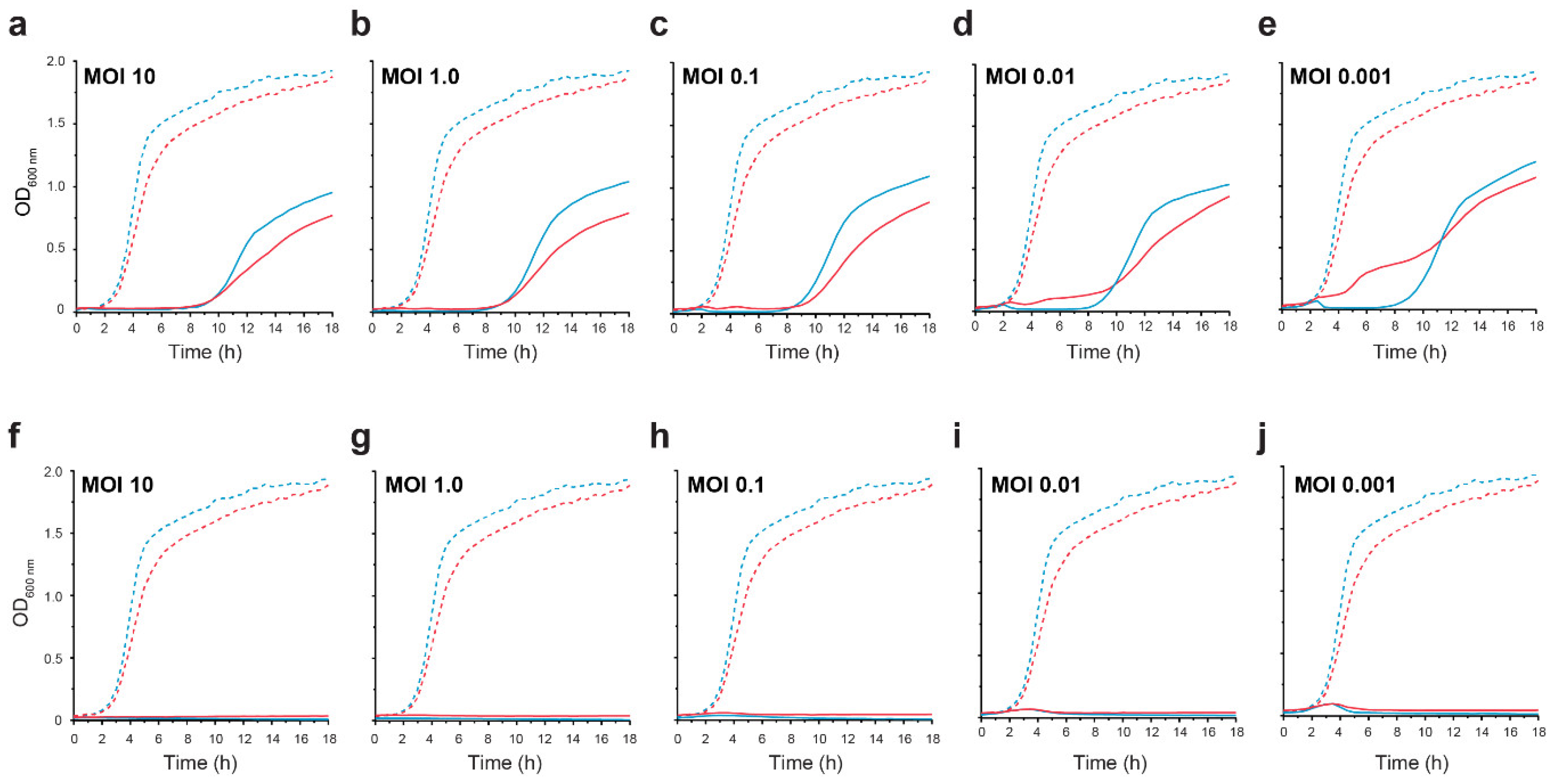
3.4. CrRp3 Is Faster at Infecting a Host Cell, but Has a Smaller Burst Size
3.5. Hypoxia Reduces Lysis at Low MOIs but Not Resistance
3.6. Host Range
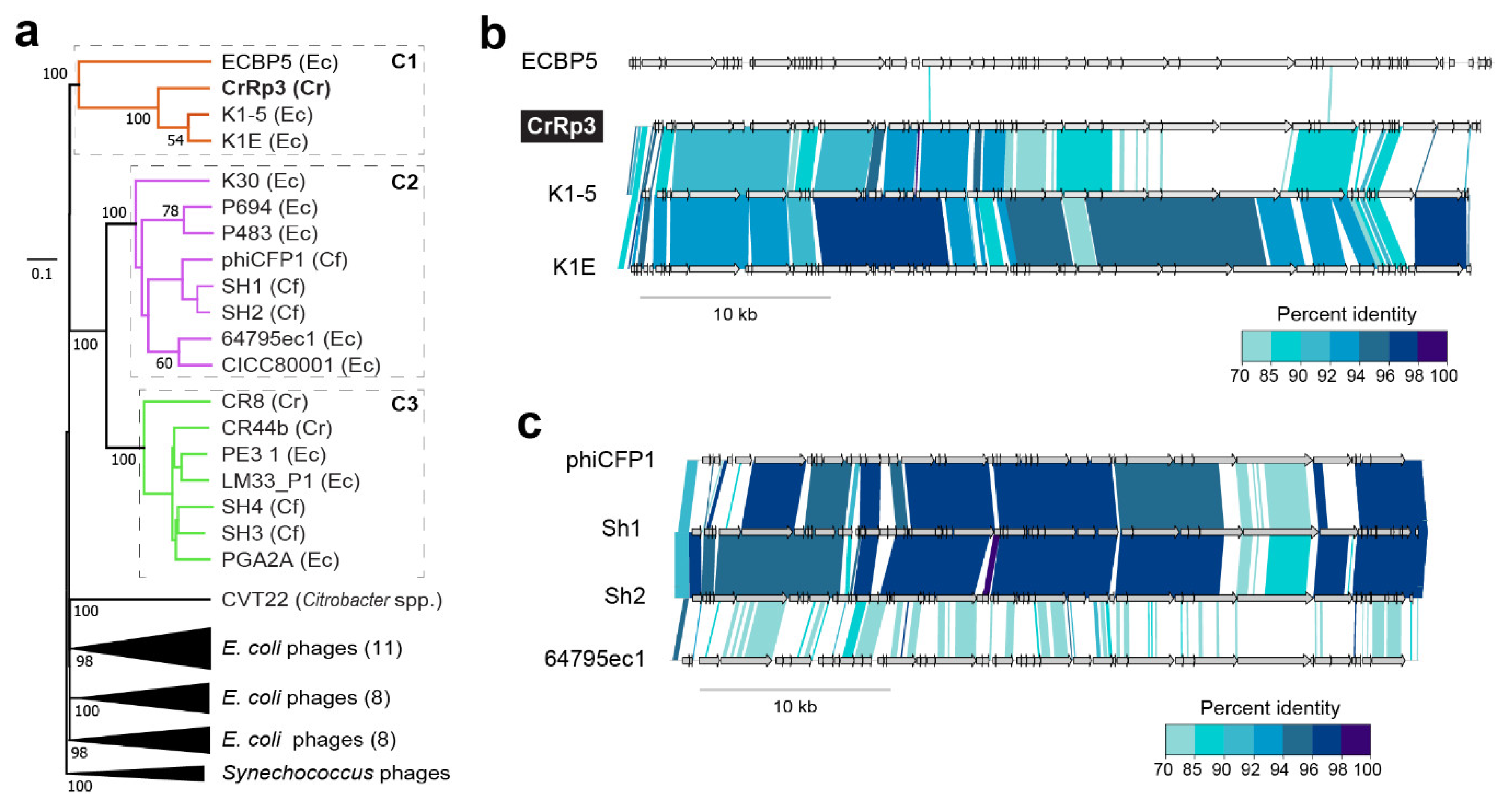
3.7. Phage Phylogenetic Relationships
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- GBD 2016 Diarrhoeal Disease Collaborators. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of diarrhoea in 195 countries: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1211–1228. [Google Scholar] [CrossRef] [Green Version]
- Qu, M.; Lv, B.; Zhang, X.; Yan, H.; Huang, Y.; Qian, H.; Pang, B.; Jia, L.; Kan, B.; Wang, Q. Prevalence and antibiotic resistance of bacterial pathogens isolated from childhood diarrhea in Beijing, China (2010–2014). Gut Pathog. 2016, 8, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, B.; Ran, L.; Wu, S.; Deng, X.; Ke, C.; Feng, Z.; Ma, L.; Varma, J.K. Survey of physician diagnostic and treatment practices for patients with acute ciarrhea in Guangdong province, China. Foodborne Pathog. Dis. 2012, 9, 47–53. [Google Scholar] [CrossRef]
- Buffie, C.G.; Pamer, E.G. Microbiota-mediated colonization resistance against intestinal pathogens. Nat. Rev. Immunol. 2013, 13, 790–801. [Google Scholar] [CrossRef] [Green Version]
- Maura, D.; Galtier, M.; Le Bouguénec, C.; Debarbieux, L. Virulent bacteriophages can target O104:H4 enteroaggregative Escherichia coli in the mouse intestine. Antimicrob. Agents Chemother. 2012, 56, 6235–6242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galtier, M.; De Sordi, L.; Sivignon, A.; De Vallée, A.; Maura, D.; Christel, N.; Rahmouni, O.; Wannerberger, K.; Darfeuille-Michaud, A.; Desreumaux, P.; et al. Bacteriophages targeting adherent invasive Escherichia coli strains as a promising new treatment for Crohn’s disease. J. Crohn’s Colitis 2017, 11, 840–847. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, U.; Ukhanova, M.; Moye, Z.D.; Sulakvelidze, A.; Mai, V. Bacteriophages reduce pathogenic Escherichia coli counts in mice without distorting gut microbiota. Front. Microbiol. 2019, 10, 1984. [Google Scholar] [CrossRef]
- Cepko, L.C.S.; Garling, E.E.; Dinsdale, M.J.; Scott, W.P.; Bandy, L.; Nice, T.J.; Faber-Hammond, J.; Mellies, J.L. Myoviridae phage PDX kills enteroaggregative Escherichia coli without human microbiome dysbiosis. J. Med. Microbiol. 2020, 69, 309–323. [Google Scholar] [CrossRef]
- Crepin, V.F.; Collins, J.W.; Habibzay, M.; Frankel, G. Citrobacter rodentium mouse model of bacterial infection. Nat. Protoc. 2016, 11, 1851–1876. [Google Scholar] [CrossRef]
- Hu, J.; Torres, A.G. Enteropathogenic Escherichia coli: Foe or innocent bystander? Clin. Microbiol. Infect. 2015, 21, 729–734. [Google Scholar] [CrossRef] [Green Version]
- Bouladoux, N.; Harrison, O.; Belkaid, Y. The mouse model of infection with Citrobacter rodentium. Curr. Protoc. Immunol. 2017, 119, 19.15.1–19.15.25. [Google Scholar] [CrossRef] [PubMed]
- Roach, D.R.; Debarbieux, L. Phage therapy: Awakening a sleeping giant. Emerg. Top. Life Sci. 2017, 1, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Altamirano, F.L.G.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef] [Green Version]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat. Med. 2019, 25, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, S.; Hamilton, T.; Biswas, B.; Kumaraswamy, M.; Schooley, R.T.; Wooten, D. Phage therapy for a multidrug-resistant Acinetobacter baumannii craniectomy site infection. Open Forum Infect. Dis. 2018, 5, ofy064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nir-Paz, R.; Gelman, D.; Khouri, A.; Sisson, B.M.; Fackler, J.; Alkalay-Oren, S.; Khalifa, L.; Rimon, A.; Yerushalmy, O.; Bader, R.; et al. Successful treatment of antibiotic-resistant, poly-microbial bone infection with bacteriophages and antibiotics combination. Clin. Infect. Dis. 2019, 69, 2015–2018. [Google Scholar] [CrossRef]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef]
- Segall, A.M.; Roach, D.R.; Strathdee, S.A. Stronger together? Perspectives on phage-antibiotic synergy in clinical applications of phage therapy. Curr. Opin. Microbiol. 2019, 51, 46–50. [Google Scholar] [CrossRef]
- Petty, N.K.; Toribio, A.; Goulding, D.; Foulds, I.; Thomson, N.R.; Dougan, G.; Salmond, G.P.C. A generalized transducing phage for the murine pathogen Citrobacter rodentium. Microbiology 2007, 153, 2984–2988. [Google Scholar] [CrossRef] [Green Version]
- Sváb, D.; Horváth, B.; Rohde, M.; Maróti, G.; Tóth, I. R18C is a new viable P2-like bacteriophage of rabbit origin infecting Citrobacter rodentium and Shigella sonnei strains. Arch. Virol. 2019, 164, 3157–3160. [Google Scholar] [CrossRef] [Green Version]
- Toribio, A.; Pickard, D.; Cerdeño-Tárraga, A.M.; Petty, N.K.; Thomson, N.R.; Salmond, G.; Dougan, G. Complete genome sequences of two Citrobacter rodentium bacteriophages, CR8 and CR44b. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiles, S.; Clare, S.; Harker, J.A.; Huett, A.; Young, D.; Dougan, G.; Frankel, G. Organ specificity, colonization and clearance dynamics in vivo following oral challenges with the murine pathogen Citrobacter rodentium. Cell Microbiol. 2004, 6, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Forti, F.; Roach, D.R.; Cafora, M.; Pasini, M.E.; Horner, D.S.; Fiscarelli, E.V.; Rossitto, M.; Cariani, L.; Briani, F.; Debarbieux, L.; et al. Design of a broad-range bacteriophage cocktail that reduces Pseudomonas aeruginosa biofilms and treats acute infections in two animal models. Antimicrob. Agents Chemother. 2018, 62, e02573-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickard, D.J.J. Preparation of bacteriophage lysates and pure DNA. Methods Mol. Biol. 2009, 502, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A Program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, D.; Chen, G.; Locascio, P.F.; Land, M.; Larimer, F.W.; Hauser, L. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soeding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haft, D.H. TIGRFAMs: A protein family resource for the functional identification of proteins. Nucleic Acids Res. 2001, 29, 41–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2016, 45, D566–D573. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.-H.; McDermott, P.F.; et al. Validating the AMRFinder tool and resistance gene database by using antimicrobial resistance genotype-phenotype correlations in a collection of isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.-M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2013, 58, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis-10 years on. Nucleic Acids Res. 2015, 44, D694–D697. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. VICTOR: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A comprehensive, accurate, and fast distance-based phylogeny inference program. Mol. Boil. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef] [Green Version]
- Kropinski, A.M. Measurement of the rate of attachment of bacteriophage to cells. Methods Mol. Biol. 2009, 501, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Kropinski, A.M. Practical advice on the one-step growth curve. Adv. Struct. Saf. Stud. 2017, 1681, 41–47. [Google Scholar] [CrossRef]
- Basset, A.; Khush, R.S.; Braun, A.; Gardan, L.; Boccard, F.; Hoffmann, J.A.; Lemaitre, B. The phytopathogenic bacteria Erwinia carotovora infects Drosophila and activates an immune response. Proc. Natl. Acad. Sci. USA 2000, 97, 3376–3381. [Google Scholar] [CrossRef] [PubMed]
- Le Flèche-Matéos, A.; Levast, M.; Lomprez, F.; Arnoux, Y.; Andonian, C.; Perraud, M.; Vincent, V.; Gouilh, M.A.; Thiberge, J.-M.; Vandenbogaert, M.; et al. Rouxiella chamberiensis gen nov., sp. nov., a member of the family Enterobacteriaceae isolated from parenteral nutrition bags. Int. J. Syst. Evol. Microbiol. 2015, 65, 1812–1818. [Google Scholar] [CrossRef] [Green Version]
- Flyg, C.; Kenne, K.; Boman, H.G. Insect pathogenic properties of Serratia marcescens: Phage-resistant mutants with a decreased resistance to Cecropia immunity and a decreased virulence to Drosophila. Microbiology 1980, 120, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Sapriel, G.; Wandersman, C.; Delepelaire, P. The SecB chaperone is bifunctional in Serratia marcescens: SecB is involved in the Sec pathway and required for HasA secretion by the ABC transporter. J. Bacteriol. 2003, 185, 80–88. [Google Scholar] [CrossRef] [Green Version]
- De Sordi, L.; Khanna, V.; Debarbieux, L. The gut microbiota facilitates drifts in the genetic diversity and infectivity of bacterial viruses. Cell Host Microbe 2017, 22, 801–808. [Google Scholar] [CrossRef] [Green Version]
- Dufour, N.; Clermont, O.; La Combe, B.; Messika, J.; Dion, S.; Khanna, V.; Denamur, E.; Ricard, J.-D.; Debarbieux, L.; on behalf of the ColoColi group. Bacteriophage LM33_P1, a fast-acting weapon against the pandemic ST131-O25b:H4 Escherichia coli clonal complex. J. Antimicrob. Chemother. 2016, 71, 3072–3080. [Google Scholar] [CrossRef] [Green Version]
- Galtier, M.; De Sordi, L.; Maura, D.; Arachchi, H.; Volant, S.; Dillies, M.-A.; Debarbieux, L. Bacteriophages to reduce gut carriage of antibiotic resistant uropathogens with low impact on microbiota composition. Environ. Microbiol. 2016, 18, 2237–2245. [Google Scholar] [CrossRef] [Green Version]
- Maura, D.; Morello, E.; Du Merle, L.; Bomme, P.; Le Bouguénec, C.; Debarbieux, L. Intestinal colonization by enteroaggregative Escherichia coli supports long-term bacteriophage replication in mice. Environ. Microbiol. 2011, 14, 1844–1854. [Google Scholar] [CrossRef]
- Dufour, N.; Debarbieux, L.; Fromentin, M.; Ricard, J.-D. Treatment of highly virulent extraintestinal pathogenic Escherichia coli pneumonia with bacteriophages. Crit. Care Med. 2015, 43, e190–e198. [Google Scholar] [CrossRef] [Green Version]
- Tikhe, C.; Martin, T.M.; Gissendanner, C.R.; Husseneder, C. Complete genome sequence of Citrobacter phage CVT22 isolated from the gut of the formosan subterranean termite, Coptotermes formosanus Shiraki. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Lesage, K.C.; Hargrove, E.C.; Cahill, J.; Rasche, E.S.; Everett, G.F.K. Complete genome sequence of Citrobacter freundii myophage Merlin. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, K.; Luna, A.J.; Hernandez, A.C.; Everett, G.F.K. Complete genome sequence of Citrobacter freundii myophage Miller. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, G.B.; Luna, A.J.; Hernández, A.C.; Everett, G.F.K. Complete genome sequence of Citrobacter freundii myophage Moon. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal, C.L.; Berkowitz, V.E.; Cahill, J.; Rasche, E.S.; Everett, G.F.K. Complete genome sequence of Citrobacter freundii myophage Michonne. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, Q.T.; Luna, A.J.; Hernández, A.C.; Everett, G.F.K. Complete genome sequence of Citrobacter freundii myophage Moogle. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.; Medina, C.A.A.; Chen, Y.; Luna, A.J.; Hernández, A.C.; Everett, G.F.K. Complete genome of Citrobacter freundii siphophage Stevie. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Adriaenssens, E.M.; Sullivan, M.B.; Knezevic, P.; Van Zyl, L.J.; Sarkar, B.L.; Dutilh, B.E.; Alfenas-Zerbini, P.; Lobocka, M.; Tong, Y.; Brister, J.R.; et al. Taxonomy of prokaryotic viruses: 2018–2019 update from the ICTV Bacterial and Archaeal Viruses Subcommittee. Arch. Virol. 2020, 165, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Fan, H.; An, X.; Fan, H.; Jiang, H.; Chen, Y.; Tong, Y. Scrutinizing virus genome termini by high-throughput sequencing. PLoS ONE 2014, 9, e85806. [Google Scholar] [CrossRef] [Green Version]
- Rivas, L.; Coffey, B.; McAuliffe, O.; McDonnell, M.J.; Burgess, C.M.; Coffey, A.; Ross, R.P.; Duffy, G. In vivo and ex vivo evaluations of bacteriophages e11/2 and e4/1c for use in the control of Escherichia coli O157:H7. Appl. Environ. Microbiol. 2010, 76, 7210–7216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albenberg, L.; Esipova, T.V.; Judge, C.P.; Bittinger, K.; Chen, J.; Laughlin, A.; Grunberg, S.; Baldassano, R.N.; Lewis, J.D.; Li, H.; et al. Correlation between intraluminal oxygen gradient and radial partitioning of intestinal microbiota. Gastroenterology 2014, 147, 1055–1063. [Google Scholar] [CrossRef] [Green Version]
- Enault, F.; Briet, A.; Bouteille, L.; Roux, S.; Sullivan, M.B.; Petit, M.-A. Phages rarely encode antibiotic resistance genes: A cautionary tale for virome analyses. ISME J. 2017, 11, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Guan, J.; Snowden, J.D.; Cahill, J.; Rasche, E.S.; Everett, G.F.K. Complete genome sequence of Citrobacter freundii myophage Mordin. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Huang, L.; Zhao, J.; He, X.; Li, E.; Li, H.; Liu, W.; Zou, D.; Wei, X.; Wang, X.; et al. Characterization of phiCFP-1, a virulent bacteriophage specific for Citrobacter freundii. J. Med. Virol. 2015, 88, 895–905. [Google Scholar] [CrossRef]
- Hamdi, S.; Rousseau, G.M.; Labrie, S.J.; Kourda, R.S.; Tremblay, D.M.; Moineau, S.; Slama, K.B. characterization of five Podoviridae phages infecting Citrobacter freundii. Front. Microbiol. 2016, 7, 1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipsky, B.A.; Hook, E.W.; Smith, A.A.; Plorde, J.J. Citrobacter infections in humans: Experience at the Seattle Veterans Administration Medical Center and a review of the literature. Rev. Infect. Dis. 1980, 2, 746–760. [Google Scholar] [CrossRef]
- Schauer, D.B.; Zabel, B.A.; Pedraza, I.F.; O’Hara, C.M.; Steigerwalt, A.G.; Brenner, D.J. Genetic and biochemical characterization of Citrobacter rodentium sp. nov. J. Clin. Microbiol. 1995, 33, 2064–2068. [Google Scholar] [CrossRef] [Green Version]
- Petty, N.K.; Bulgin, R.; Crepin, V.F.; Cerdeño-Tárraga, A.M.; Schroeder, G.; Quail, M.A.; Lennard, N.; Corton, C.; Barron, A.; Clark, L.; et al. The Citrobacter rodentium genome sequence reveals convergent evolution with human pathogenic Escherichia coli. J. Bacteriol. 2009, 192, 525–538. [Google Scholar] [CrossRef] [Green Version]
- Petty, N.K.; Feltwell, T.; Pickard, D.; Clare, S.; Toribio, A.; Fookes, M.; Roberts, K.; Monson, R.; Nair, S.; Kingsley, R.A.; et al. Citrobacter rodentium is an unstable pathogen showing evidence of significant genomic flux. PLoS Pathog. 2011, 7, e1002018. [Google Scholar] [CrossRef] [PubMed]
- Scholl, D.; Adhya, S.; Merril, C.R. Bacteriophage SP6 is closely related to phages K1-5, K5, and K1E but encodes a tail protein very similar to that of the distantly related P22. J. Bacteriol. 2002, 184, 2833–2836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholl, D.; Kieleczawa, J.; Kemp, P.; Rush, J.; Richardson, C.; Merril, C.; Adhya, S.; Molineux, I. Genomic analysis of bacteriophages SP6 and K1-5, an estranged subgroup of the T7 supergroup. J. Mol. Boil. 2004, 335, 1151–1171. [Google Scholar] [CrossRef] [PubMed]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Genet. 2010, 8, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.C.T.; Friman, V.-P.; Smith, M.C.M.; Brockhurst, M.A. Resistance evolution against phage combinations depends on the timing and order of exposure. MBio 2019, 10, e01652-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Haddad, L.; Harb, C.P.; Gebara, M.A.; Stibich, M.A.; Chemaly, R. A systematic and critical review of bacteriophage therapy against multidrug-resistant ESKAPE organisms in humans. Clin. Infect. Dis. 2019, 69, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.; Hawkins, C.; Änggård, E.; Harper, D. A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa; a preliminary report of efficacy. J. Clin. Otolaryngol. 2009, 34, 349–357. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Carmeliet, P. Hypoxia and inflammation. N. Engl. J. Med. 2011, 364, 656–665. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Liu, X.; Zha, H.; Fan, S.; Zhang, D.; Li, S.; Xiao, W. A pathogen-derived effector modulates host glucose metabolism by arginine GlcNAcylation of HIF-1α protein. PLoS Pathog. 2018, 14, e1007259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenco, M.; De Sordi, L.; Debarbieux, L. The diversity of bacterial lifestyles hampers bacteriophage tenacity. Viruses 2018, 10, 327. [Google Scholar] [CrossRef] [Green Version]
- You, L.; Suthers, P.F.; Yin, J. Effects of Escherichia coli physiology on growth of phage T7 in vivo and in silico. J. Bacteriol. 2002, 184, 1888–1894. [Google Scholar] [CrossRef] [Green Version]
- McDaniel, L.E.; Bailey, E.G.; Zimmerli, A. Effect of oxygen-supply rates on growth of Escherichia coli. Ii. Comparison of results in shake flasks and 50-liter fermenter. Appl. Microbiol. 1965, 13, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Swift, B.M.C.; Gerrard, Z.E.; Huxley, J.; Rees, C. Factors affecting phage D29 infection: A tool to investigate different growth states of mycobacteria. PLoS ONE 2014, 9, e106690. [Google Scholar] [CrossRef] [PubMed]
- Hadas, H.; Einav, M.; Fishov, I.; Zaritsky, A. Bacteriophage T4 development depends on the physiology of its host Escherichia coli. Microbiology 1997, 143, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Hyman, P.; Abedon, S.T. Bacteriophage host range and bacterial resistance. Adv. Appl. Microbiol. 2010, 70, 217–248. [Google Scholar] [CrossRef] [PubMed]
- Szajewska, H.; Canani, R.B.; Guarino, A.; Hojsak, I.; Indrio, F.; Kolaček, S.; Orel, R.; Shamir, R.; Vandenplas, Y.; Van Goudoever, J.B.; et al. Probiotics for the prevention of antibiotic-associated diarrhea in children. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Servick, K. Beleaguered phage therapy trial presses on. Science 2016, 352, 1506. [Google Scholar] [CrossRef]
- Croxen, M.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, B.K.; Abedon, S.; Loc-Carrillo, C. Phage cocktails and the future of phage therapy. Future Microbiol. 2013, 8, 769–783. [Google Scholar] [CrossRef]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and use of personalized bacteriophage-based therapeutic cocktails to treat a patient with a disseminated resistant Acinetobacter baumannii infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage | Host | Source | Phage Family * | Size (kb) | GC% | Accession No. | Ref. |
|---|---|---|---|---|---|---|---|
| CrRp3 | C. rodentium | wastewater | A | 44.3 | 45.1 | MG775042 | This study |
| CR44b | C. rodentium | sewage effluent | A | 39.2 | 50.5 | NC_023576 | [21] |
| CR8 | C. rodentium | sewage effluent | A | 39.7 | 49.7 | NC_023548 | [21] |
| CVT22 | Citrobacter sp. | termite gut | A | 47.6 | 41.6 | NC_027988 | [51] |
| phiCFP-1 | C. freundii | seawater | A | 38.6 | 50.3 | NC_028880 | N/A |
| SH1 | C. freundii | seawater | A | 39.4 | 51 | NC_031066 | N/A |
| SH2 | C. freundii | seawater | A | 39.2 | 50.7 | NC_031092 | N/A |
| SH3 | C. freundii | seawater | A | 39.4 | 50.6 | NC_031123 | N/A |
| SH4 | C. freundii | seawater | A | 39.3 | 52.6 | NC_031018 | N/A |
| CrRp10 | C. rodentium | municipal wastewater | M | 171.5 | 35.5 | MG775043 | This study |
| R18C | C. rodentium | rabbit feces | M | 31.8 | 51.6 | MN016939 | [20] |
| IME-CF2 | C. freundii | hospital wastewater | M | 177.7 | 43.2 | NC_029013 | N/A |
| Margaery | C. freundii | wastewater | M | 178.2 | 44.9 | NC_028755 | N/A |
| Merlin | C. freundii | wastewater | M | 172.7 | 38.8 | NC_028857 | [52] |
| Miller | C. freundii | wastewater | M | 178.2 | 43.1 | NC_025414 | [53] |
| Moon | C. freundii | wastewater | M | 170.3 | 38.9 | NC_027331 | [54] |
| Michonne | C. freundii | wastewater | M | 90.0 | 38.8 | NC_028247 | [55] |
| Moogle | C. freundii | wastewater | M | 88.0 | 39 | NC_027293 | [56] |
| CfP1 | C. freundii | sewage effluent | M | 180.2 | 43.1 | NC_031057 | N/A |
| Stevie | C. freundii | soil | D | 49.8 | 42.8 | NC_027350 | [57] |
| CrRp3 | CrRp10 | |
|---|---|---|
| Family | Autographiviridae | Myoviridae |
| Adsorption constant 1 (mL−1 min−1) | k =3.50 × 10−10 (±3.2 × 10−10) | k =8.52 × 10−11 (±2.8 × 10−11) |
| Latent period 2 | 15 (±2) min | 17 (±2) min |
| Mean burst size | 43 (±22) PFU | 85 (±16) PFU |
 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mizuno, C.M.; Luong, T.; Cederstrom, R.; Krupovic, M.; Debarbieux, L.; Roach, D.R. Isolation and Characterization of Bacteriophages That Infect Citrobacter rodentium, a Model Pathogen for Intestinal Diseases. Viruses 2020, 12, 737. https://doi.org/10.3390/v12070737
Mizuno CM, Luong T, Cederstrom R, Krupovic M, Debarbieux L, Roach DR. Isolation and Characterization of Bacteriophages That Infect Citrobacter rodentium, a Model Pathogen for Intestinal Diseases. Viruses. 2020; 12(7):737. https://doi.org/10.3390/v12070737
Chicago/Turabian StyleMizuno, Carolina M., Tiffany Luong, Robert Cederstrom, Mart Krupovic, Laurent Debarbieux, and Dwayne R. Roach. 2020. "Isolation and Characterization of Bacteriophages That Infect Citrobacter rodentium, a Model Pathogen for Intestinal Diseases" Viruses 12, no. 7: 737. https://doi.org/10.3390/v12070737
APA StyleMizuno, C. M., Luong, T., Cederstrom, R., Krupovic, M., Debarbieux, L., & Roach, D. R. (2020). Isolation and Characterization of Bacteriophages That Infect Citrobacter rodentium, a Model Pathogen for Intestinal Diseases. Viruses, 12(7), 737. https://doi.org/10.3390/v12070737