Whole Genomic Analysis and Comparison of Two Canine Papillomavirus Type 9 Strains in Malignant and Benign Skin Lesions

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Ethical Statement
- Dog 1: Fresh collected and formalin-fixed paraffin-embedded (FFPE) tissue samples from Dog 1 with recurrent SCCs were obtained from the animal hospitals at National Taiwan University in 2018. The sample collections were performed following the guidelines of and under the permissions of the Institutional Animal Care and Use Committees (IACUCs) of National Taiwan University (NTU) with the permission numbers NTU107-EL-00165.
- Dog 2: The FFPE sample from Dog 2 was obtained from the Department of Veterinary Pathology, The University of Tokyo (UT) in 2018 which was deposited for diagnostic purposes and to enable adequate disease prevention strategies. All surgical procedures were performed by educated veterinarians in accordance with Japanese law.
2.2. Case Information
2.3. Histological Descriptions and Immunohistochemistry Staining (IHC)
2.4. DNA Extraction from FFPE and Fresh Tissue
2.5. Polymerase Chain Reaction (PCR) for the Detection and Full Genome Sequencing of CPV
2.6. Sequence Analysis
2.7. RNA Extraction and Genomic DNA Eradication from FFPE Tissue
2.8. Primer-Specific cDNA Synthesis of E6 and E7
2.9. The Detection of mRNA Expression Level of E6 and E7 by PCR and qPCR
3. Results
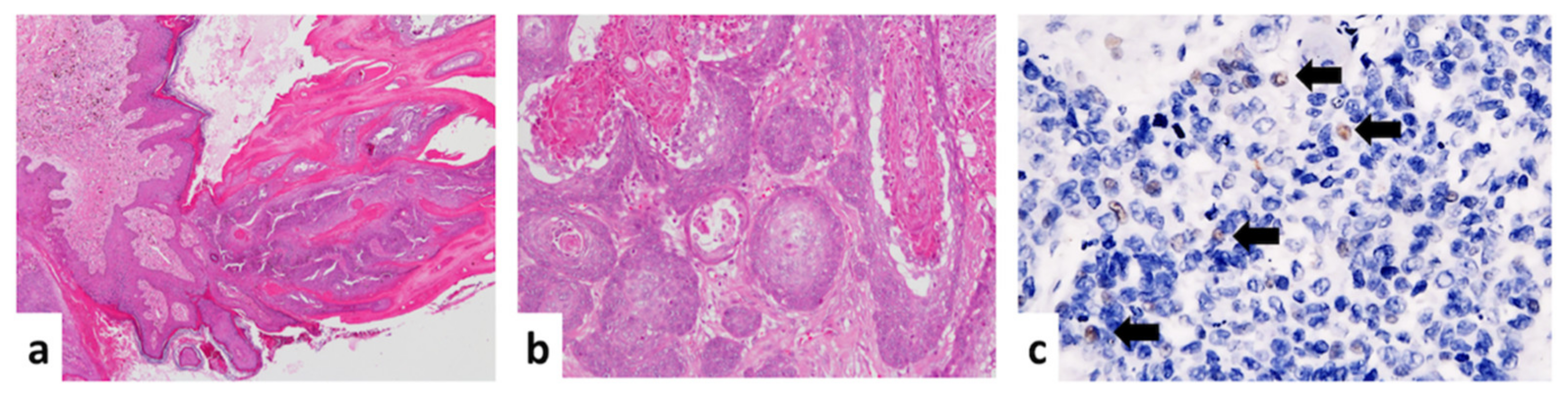
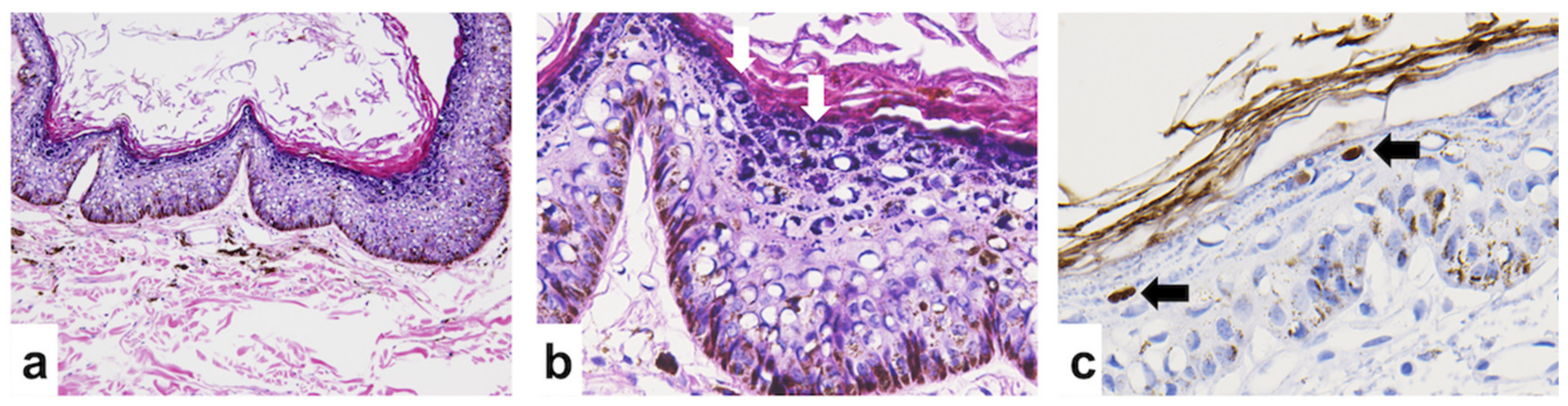
3.1. Histological Descriptions and Immunohistochemistry Staining
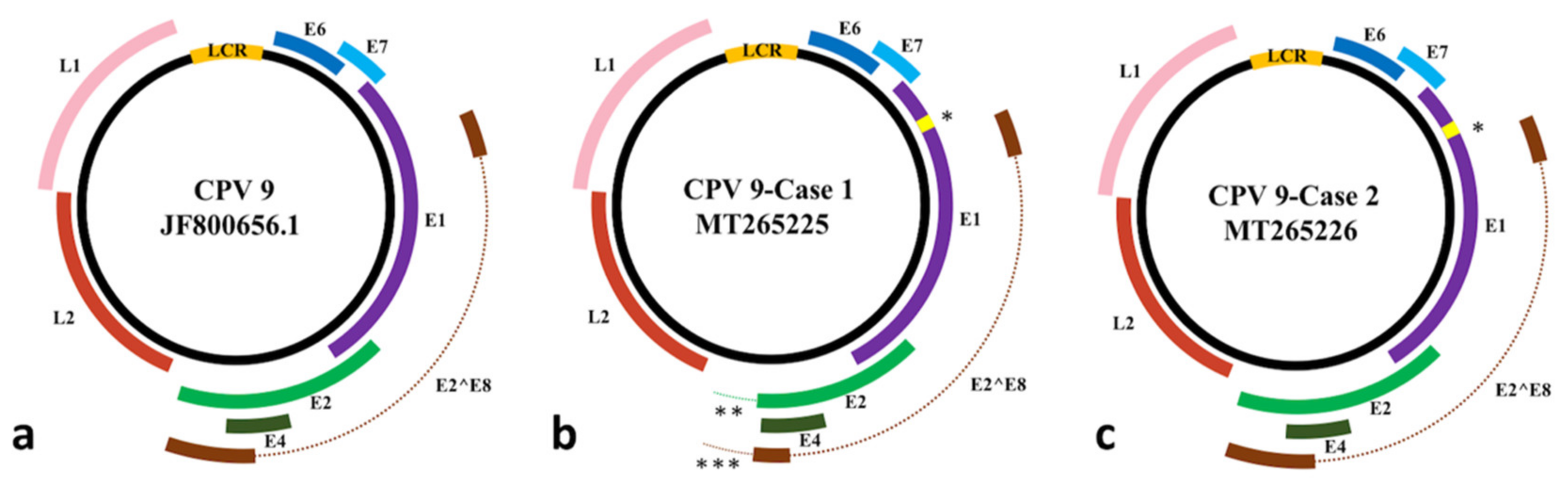
3.2. Complete Nucleotide Sequence of CPV9
3.3. Open Reading Frames (ORFs) and Deduced Amino Acid Comparisons
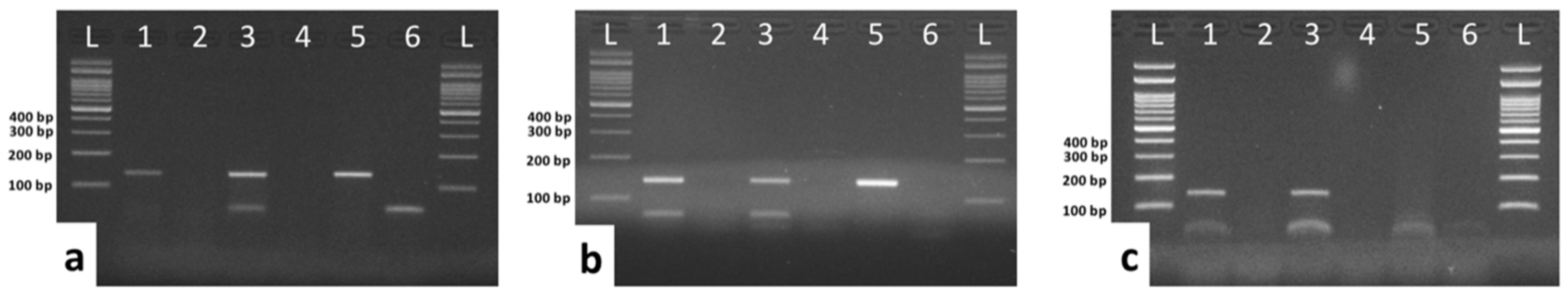
3.4. Detection of E6 and E7 mRNA by conventional RT-PCR
3.5. Comparison of mRNA Expression Levels of E6 and E7 Using Quantitative PCR (qPCR)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Münger, K.; Baldwin, A.; Edwards, K.M.; Hayakawa, H.; Nguyen, C.L.; Owens, M.; Grace, M.; Huh, K. Mechanisms of Human Papillomavirus-Induced Oncogenesis. J. Virol. 2004, 78, 11451–11460. [Google Scholar] [CrossRef] [Green Version]
- Zur Hausen, H.; de Villiers, E.M.; Gissmann, L. Papillomavirus infections and human genital cancer. Gynecol. Oncol. 1981, 12 Pt 2, S124–S128. [Google Scholar] [CrossRef]
- Gil da Costa, R.M.; Peleteiro, M.C.; Pires, M.A.; DiMaio, D. An Update on Canine, Feline and Bovine Papillomaviruses. Transbound. Emerg. Dis. 2017, 64, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Doorbar, J.; Egawa, N.; Griffin, H.; Kranjec, C.; Murakami, I. Human papillomavirus molecular biology and disease association. Rev. Med. Virol. 2015, 25, 2–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munday, J.S.; Thomson, N.A.; Luff, J.A. Papillomaviruses in dogs and cats. Vet. J. 2017, 225, 23–31. [Google Scholar] [CrossRef]
- Hibma, M.H. The immune response to papillomavirus during infection persistence and regression. Open Virol. J. 2012, 6, 241–248. [Google Scholar] [CrossRef]
- Stanley, M.A. Epithelial Cell Responses to Infection with Human Papillomavirus. Clin. Microbiol. Rev. 2012, 25, 215–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, C.E.; Favrot, C. Canine Papillomaviruses. Vet. Clin. N. Am. Small Anim. Pract. 2011, 41, 1183–1195. [Google Scholar] [CrossRef]
- Smith, J.S.; Lindsay, L.; Hoots, B.; Keys, J.; Franceschi, S.; Winer, R.; Clifford, G.M. Human papillomavirus type distribution in invasive cervical cancer and high-grade cervical lesions: A meta-analysis update. Int. J. Cancer 2007, 121, 621–632. [Google Scholar] [CrossRef]
- Doorbar, J. The papillomavirus life cycle. J. Clin. Virol. 2005, 32 (Suppl. 1), S7–S15. [Google Scholar] [CrossRef]
- Kirnbauer, R.; Booy, F.; Cheng, N.; Lowy, D.R.; Schiller, J.T. Papillomavirus L1 major capsid protein self-assembles into virus-like particles that are highly immunogenic. Proc. Natl. Acad. Sci. USA 1992, 89, 12180–12184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnen, R.L.; Erickson, K.D.; Chen, X.S.; Garcea, R.L. Interactions between Papillomavirus L1 and L2 Capsid Proteins. J. Virol. 2003, 77, 4818–4826. [Google Scholar] [CrossRef] [Green Version]
- Hamid, N.A.; Brown, C.; Gaston, K. The regulation of cell proliferation by the papillomavirus early proteins. Cell. Mol. Life Sci. 2009, 66, 1700–1717. [Google Scholar] [CrossRef]
- Hegde, R.S. The Papillomavirus E2 Proteins: Structure, Function, and Biology. Annu. Rev. Biophys. Biomol. Struct. 2002, 31, 343–360. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Yue, Y. Human papillomavirus oncoproteins and apoptosis. Exp. Ther. Med. 2014, 7, 3–7. [Google Scholar] [CrossRef] [Green Version]
- McBride, A.A. The papillomavirus E2 proteins. Virology 2013, 445, 57–79. [Google Scholar] [CrossRef] [Green Version]
- Wilson, V.G.; West, M.; Woytek, K.; Rangasamy, D. Papillomavirus E1 proteins: Form, function, and features. Virus Genes 2002, 24, 275–290. [Google Scholar] [CrossRef] [PubMed]
- Bergvall, M.; Melendy, T.; Archambault, J. The E1 proteins. Virology 2013, 445, 35–56. [Google Scholar] [CrossRef] [Green Version]
- Graham, S.V. Human Papillomavirus E2 Protein: Linking Replication, Transcription, and RNA Processing. J. Virol. 2016, 90, 8384–8388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, T.; Brandsma, J.L.; Peng, X.; Srimatkandada, S.; Li, L.; Canaan, A.; Deisseroth, A.B. High and low levels of cottontail rabbit papillomavirus E2 protein generate opposite effects on gene expression. J. Biol. Chem. 2001, 276, 867–874. [Google Scholar] [CrossRef] [Green Version]
- Puustusmaa, M.; Abroi, A. Conservation of the E8 CDS of the E8^E2 protein among mammalian papillomaviruses. J. Gen. Virol. 2016, 97, 2333–2345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yajid, A.I.; Zakariah, M.A.; Zin, A.A.M.; Othman, N.H. Potential Role of E4 Protein in Human Papillomavirus Screening: A Review. Asian Pac. J. Cancer Prev. 2017, 18, 315–319. [Google Scholar] [PubMed]
- Suarez, I.; Trave, G. Structural Insights in Multifunctional Papillomavirus Oncoproteins. Viruses 2018, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Westrich, J.A.; Warren, C.J.; Pyeon, D. Evasion of host immune defenses by human papillomavirus. Virus Res. 2017, 231, 21–33. [Google Scholar] [CrossRef]
- Rector, A.; Van Ranst, M. Animal papillomaviruses. Virology 2013, 445, 213–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, C.E.; Tobler, K.; Markau, T.; Alhaidari, Z.; Bornand, V.; Stockli, R.; Trussel, M.; Ackermann, M.; Favrot, C. Sequence and classification of FdPV2, a papillomavirus isolated from feline Bowenoid in situ carcinomas. Vet. Microbiol. 2009, 137, 60–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, C.G.; Dunowska, M.; Munday, J.S.; Peters-Kennedy, J.; Rosa, B.V. Comparison of the levels of Equus caballus papillomavirus type 2 (EcPV-2) DNA in equine squamous cell carcinomas and non-cancerous tissues using quantitative PCR. Vet. Microbiol. 2013, 166, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.E.; Diallo, A.; Zewe, C.; Ferrer, L. Novel canine papillomavirus type 18 found in pigmented plaques. Papillomavirus Res. 2016, 2, 159–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tisza, M.J.; Yuan, H.; Schlegel, R.; Buck, C.B. Genomic Sequence of Canine Papillomavirus 19. Genome Announc. 2016, 4. [Google Scholar] [CrossRef]
- Thaiwong, T.; Sledge, D.G.; Wise, A.G.; Olstad, K.; Maes, R.K.; Kiupel, M. Malignant transformation of canine oral papillomavirus (CPV1)-associated papillomas in dogs: An emerging concern? Papillomavirus Res. 2018, 6, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Munday, J.S.; Dunowska, M.; Laurie, R.E.; Hills, S. Genomic characterisation of canine papillomavirus type 17, a possible rare cause of canine oral squamous cell carcinoma. Vet. Microbiol. 2016, 182, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Chen, W.T.; Haga, T.; Yamashita, N.; Lee, C.F.; Tsuzuki, M.; Chang, H.W. The Detection and Association of Canine Papillomavirus with Benign and Malignant Skin Lesions in Dogs. Viruses 2020, 12, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, C.E.; Zollinger, S.; Tobler, K.; Ackermann, M.; Favrot, C. Clinically Healthy Skin of Dogs Is a Potential Reservoir for Canine Papillomaviruses. J. Clin. Microbiol. 2011, 49, 707–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkhof, B.; Spee, B.; Rothuizen, J.; Penning, L.C. Development and evaluation of canine reference genes for accurate quantification of gene expression. Anal. Biochem. 2006, 356, 36–43. [Google Scholar] [CrossRef]
- Nagamine, E.; Hirayama, K.; Matsuda, K.; Okamoto, M.; Ohmachi, T.; Uchida, K.; Kadosawa, T.; Taniyama, H. Invasive Front Grading and Epithelial-Mesenchymal Transition in Canine Oral and Cutaneous Squamous Cell Carcinomas. Vet. Pathol. 2017, 54, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Soeda, E.; Ferran, M.C.; Baker, C.C.; McBride, A.A. Repression of HPV16 early region transcription by the E2 protein. Virology 2006, 351, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Zemlo, T.R.; Lohrbach, B.; Lambert, P.F. Role of transcriptional repressors in transformation by bovine papillomavirus type 1. J. Virol. 1994, 68, 6787–6793. [Google Scholar] [CrossRef] [Green Version]
- Straub, E.; Dreer, M.; Fertey, J.; Iftner, T.; Stubenrauch, F. The viral E8^E2C repressor limits productive replication of human papillomavirus 16. J. Virol. 2014, 88, 937–947. [Google Scholar] [CrossRef] [Green Version]
- Bernard, H.-U. Taxonomy and phylogeny of papillomaviruses: An overview and recent developments. Infect. Genet. Evol. 2013, 18, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Luff, J.; Zhou, D.; Wang, J.; Affolter, V.; Moore, P.; Schlegel, R. Complete Genome Sequence of Canine Papillomavirus Type 9. J. Virol. 2012, 86, 5966. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Luff, J.; Paul, S.; Alkhilaiwi, F.; Usuda, Y.; Wang, N.; Affolter, V.; Moore, P.; Schlegel, R.; Yuan, H. Complete genome sequence of canine papillomavirus virus type 12. Genome Announc. 2015, 3, e00294-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.; Paul, S.; Alkhilaiwi, F.; Clark, M.; Schlegel, R.; Yuan, H. Canine papillomavirus 20 isolate Ada, complete genome. GenBank. 2016. Available online: https://www.ncbi.nlm.nih.gov/nucleotide/1008264056 (accessed on 30 October 2015).
- Narechania, A.; Chen, Z.; DeSalle, R.; Burk, R.D. Phylogenetic Incongruence among Oncogenic Genital Alpha Human Papillomaviruses. J. Virol. 2005, 79, 15503–15510. [Google Scholar] [CrossRef] [Green Version]
- Carvajal-Rodríguez, A. Detecting recombination and diversifying selection in human alpha-papillomavirus. Infect. Genet. Evol. 2008, 8, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Bolatti, E.M.; Chouhy, D.; Casal, P.E.; Pérez, G.R.; Stella, E.J.; Sanchez, A.; Gorosito, M.; Bussy, R.F.; Giri, A.A. Characterization of novel human papillomavirus types 157, 158 and 205 from healthy skin and recombination analysis in genus γ-Papillomavirus. Infect. Genet. Evol. 2016, 42, 20–29. [Google Scholar] [CrossRef]
- Dreer, M.; van de Poel, S.; Stubenrauch, F. Control of viral replication and transcription by the papillomavirus E8^E2 protein. Virus Res. 2017, 231, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Lace, M.J.; Anson, J.R.; Thomas, G.S.; Turek, L.P.; Haugen, T.H. The E8^E2 gene product of human papillomavirus type 16 represses early transcription and replication but is dispensable for viral plasmid persistence in keratinocytes. J. Virol. 2008, 82, 10841–10853. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORF | CPV9 Strain | Location | Gene Length | Protein Length |
|---|---|---|---|---|
| E6 | Dog 1 | 263–718 | 456 bp | 151 a.a. |
| Dog 2 | ||||
| JF800656.1 | ||||
| E7 | Dog 1 | 678–980 | 303 bp | 100 a.a. |
| Dog 2 | ||||
| JF800656.1 | ||||
| E1 | Dog 1 | 970–2862 | 1893 bp | 630 a.a. |
| Dog 2 | 970–2862 | 1893 bp | 630 a.a. | |
| JF800656.1 | 970–2835 | 1866 bp | 621 a.a. | |
| E2 | Dog 1 | 2804–4099 | 1296 bp | 431 a.a. |
| Dog 2 | 2804–4267 | 1464 bp | 487 a.a. | |
| JF800656.1 | 2777–4240 | 1464 bp | 487 a.a. | |
| E8^E2 | Dog 1 | 1352–1389^3409–4099 | 729 bp | 242 a.a. |
| Dog 2 | 1352–1389^3409–4267 | 897 bp | 298 a.a. | |
| JF800656.1 | 1325–1362^3382–4240 | 897 bp | 298 a.a. | |
| E4 | Dog 1 | 3375–4028 | 654 bp | 217 a.a. |
| Dog 2 | 3375–4028 | |||
| JF800656.1 | 3348–4001 | |||
| L2 | Dog 1 | 4180–5697 | 1518 bp | 505 a.a. |
| Dog 2 | 4508–6025 | |||
| JF800656.1 | 4481–5998 | |||
| L1 | Dog 1 | 5750–7255 | 1506 bp | 502 a.a. |
| Dog 2 | 6078–7583 | |||
| JF800656.1 | 6051–7556 |
| Point Mutations | ||||||
|---|---|---|---|---|---|---|
| Position | Reference Strain (JF800656.1) | Dog 1 (MT265225) | Dog 2 (MT265226) | |||
| Nucleotide | Amino Acid | Nucleotide | Amino Acid | Nucleotide | Amino Acid | |
| 3806 | C | D | T | D | T | D |
| 4579 | T | I | T | I | C | I |
| 4927 | C | S | C | S | T | S |
| 4987 | C | S | C | S | T | S |
| 5011 | A | V | G | V | A | V |
| 5080 | T | T | A | T | T | T |
| 5166 | T | I | T | I | C | T* |
| 5228 | C | R | A | R | A | R |
| 5233 | T | R | G | R | T | R |
| 5281 | C | P | C | P | T | P |
| 5910 | A | D | G | G* | A | D |
| 6125 | C | D | C | D | T | D |
| 6128 | T | S | T | S | A | S |
| 6722 | A | V | T | V | A | V |
| 6779 | G | G | A | G | G | G |
| 7098 | A | T | A | T | T | S* |
| 7343 | G | P | A | P | G | P |
| 7542 | C | R | C | R | A | R |
| Insertion | ||||||
| 1113 | Reference Strain | G | ||||
| Dog 1 | TAGCTTGGAGGGATCAA ATAGTCAGGGG | |||||
| Dog 2 | CAGCTTGGAGGGATCAAATA GTCAGGGG | |||||
| Deletion | ||||||
| 4037–4364 | Dog 1 | 328 bp deletion | ||||
| Dog 2 | No deletion | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, C.-Y.; Yamashita-Kawanishi, N.; Tomizawa, S.; Liu, I.-L.; Chen, W.-T.; Chang, Y.-C.; Huang, W.-H.; Tsai, P.-S.; Shirota, K.; Chambers, J.K.; et al. Whole Genomic Analysis and Comparison of Two Canine Papillomavirus Type 9 Strains in Malignant and Benign Skin Lesions. Viruses 2020, 12, 736. https://doi.org/10.3390/v12070736
Chang C-Y, Yamashita-Kawanishi N, Tomizawa S, Liu I-L, Chen W-T, Chang Y-C, Huang W-H, Tsai P-S, Shirota K, Chambers JK, et al. Whole Genomic Analysis and Comparison of Two Canine Papillomavirus Type 9 Strains in Malignant and Benign Skin Lesions. Viruses. 2020; 12(7):736. https://doi.org/10.3390/v12070736
Chicago/Turabian StyleChang, Chia-Yu, Nanako Yamashita-Kawanishi, Sonoka Tomizawa, I-Li Liu, Wei-Tao Chen, Yen-Chen Chang, Wei-Hsiang Huang, Pei-Shiue Tsai, Kinji Shirota, James K Chambers, and et al. 2020. "Whole Genomic Analysis and Comparison of Two Canine Papillomavirus Type 9 Strains in Malignant and Benign Skin Lesions" Viruses 12, no. 7: 736. https://doi.org/10.3390/v12070736
APA StyleChang, C. -Y., Yamashita-Kawanishi, N., Tomizawa, S., Liu, I. -L., Chen, W. -T., Chang, Y. -C., Huang, W. -H., Tsai, P. -S., Shirota, K., Chambers, J. K., Uchida, K., Haga, T., & Chang, H. -W. (2020). Whole Genomic Analysis and Comparison of Two Canine Papillomavirus Type 9 Strains in Malignant and Benign Skin Lesions. Viruses, 12(7), 736. https://doi.org/10.3390/v12070736