
Cell Entry of Animal Coronaviruses


Abstract
:1. Introduction
2. Overview of Coronaviral S Proteins
3. Cell Binding
3.1. Cellular Glycoprotein Receptor
3.2. Receptor-Binding Domains and Residues in the Coronaviral S Proteins
3.3. Attachment Factors
3.3.1. Sialic Acid (SA)
3.3.2. Heparan Sulfate (HS)
3.3.3. C-Type Lectins
4. Membrane Fusion
4.1. Structure and Function of the S2 Domain
4.2. Cleavage of S Proteins by Host Proteases
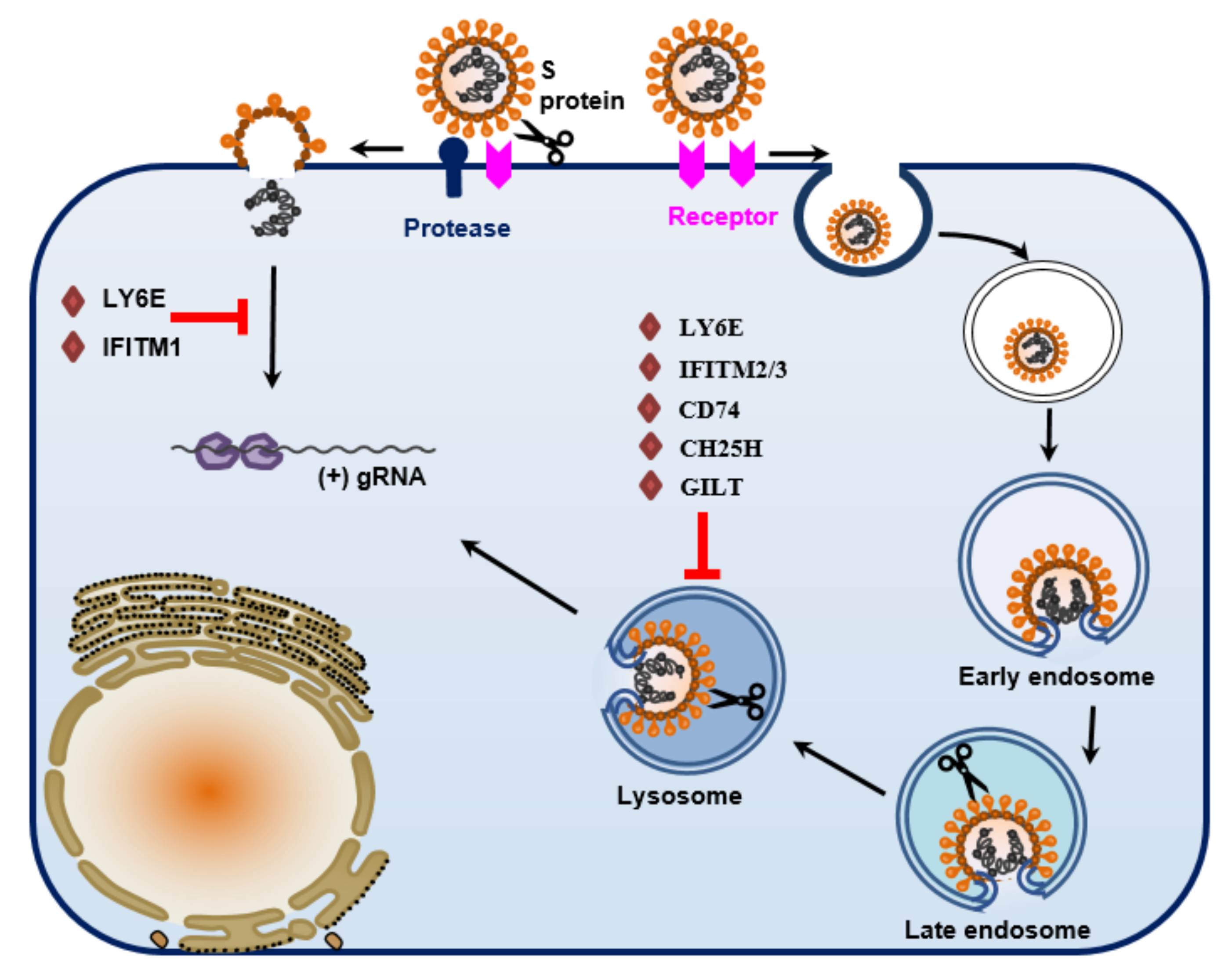
5. Host Restriction Factors for Coronavirus Entry
5.1. IFITM
5.2. LY6E
5.3. CD74
5.4. Ezrin
5.5. GILT
5.6. CH25H
6. Perspective of Future Study
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lai, M.M.; Holmes, K.V. Coronaviridae: The Viruses and Their Replication. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 2001; pp. 1163–1186. [Google Scholar]
- Cavanagh, D. Coronavirus Avian Infectious Bronchitis Virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laude, H.; Rasschaert, D.; Delmas, B.; Godet, M.; Gelfi, J.; Charley, B. Molecular Biology of Transmissible Gastroenteritis Virus. Vet. Microbiol. 1990, 23, 147–154. [Google Scholar] [CrossRef]
- Gallagher, T.M.; Buchmeier, M.J. Coronavirus Spike Proteins in Viral Entry and Pathogenesis. Virology 2001, 279, 371–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millet, J.K.; Whittaker, G.R. Host Cell Proteases: Critical Determinants of Coronavirus Tropism and Pathogenesis. Virus Res. 2015, 202, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Wentworth, D.E.; Holmes, K.V. Coronavirus Binding and Entry. In Coronaviruses: Molecular and Cellular Biology; Thiel, V., Ed.; Caister Academic Press: Norfolk, UK, 2007; pp. 3–30. [Google Scholar]
- Kuo, L.; Godeke, G.J.; Raamsman, M.J.; Masters, P.S.; Rottier, P.J. Retargeting of Coronavirus by Substitution of the Spike Glycoprotein Ectodomain: Crossing the Host Cell Species Barrier. J. Virol. 2000, 74, 1393–1406. [Google Scholar] [CrossRef] [Green Version]
- Haijema, B.J.; Volders, H.; Rottier, P.J.M. Switching Species Tropism: An Effective Way to Manipulate the Feline Coronavirus Genome. J. Virol. 2003, 77, 4528–4538. [Google Scholar] [CrossRef] [Green Version]
- Masters, P.S. The Molecular Biology of Coronaviruses. Adv. Virus Res. 2006, 66, 193–292. [Google Scholar] [CrossRef]
- Du, L.; He, Y.; Zhou, Y.; Liu, S.; Zheng, B.-J.; Jiang, S. The Spike Protein of SARS-CoV-a Target for Vaccine and Therapeutic Development. Nat. Rev. Microbiol. 2009, 7, 226–236. [Google Scholar] [CrossRef]
- Ballesteros, M.L.; Sánchez, C.M.; Enjuanes, L. Two Amino Acid Changes at the N-Terminus of Transmissible Gastroenteritis Coronavirus Spike Protein Result in the Loss of Enteric Tropism. Virology 1997, 227, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Krempl, C.; Schultze, B.; Laude, H.; Herrler, G. Point Mutations in the S Protein Connect the Sialic Acid Binding Activity with the Enteropathogenicity of Transmissible Gastroenteritis Coronavirus. J. Virol. 1997, 71, 3285–3287. [Google Scholar] [CrossRef] [Green Version]
- Dveksler, G.S.; Dieffenbach, C.W.; Cardellichio, C.B.; McCuaig, K.; Pensiero, M.N.; Jiang, G.S.; Beauchemin, N.; Holmes, K.V. Several Members of the Mouse Carcinoembryonic Antigen-Related Glycoprotein Family Are Functional Receptors for the Coronavirus Mouse Hepatitis Virus-A59. J. Virol. 1993, 67, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dveksler, G.S.; Pensiero, M.N.; Cardellichio, C.B.; Williams, R.K.; Jiang, G.S.; Holmes, K.V.; Dieffenbach, C.W. Cloning of the Mouse Hepatitis Virus (MHV) Receptor: Expression in Human and Hamster Cell Lines Confers Susceptibility to MHV. J. Virol. 1991, 65, 6881–6891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.K.; Jiang, G.S.; Holmes, K.V. Receptor for Mouse Hepatitis Virus Is a Member of the Carcinoembryonic Antigen Family of Glycoproteins. Proc. Natl. Acad. Sci. USA 1991, 88, 5533–5536. [Google Scholar] [CrossRef] [Green Version]
- Pensiero, M.N.; Dveksler, G.S.; Cardellichio, C.B.; Jiang, G.S.; Elia, P.E.; Dieffenbach, C.W.; Holmes, K.V. Binding of the Coronavirus Mouse Hepatitis Virus A59 to Its Receptor Expressed from a Recombinant Vaccinia Virus Depends on Posttranslational Processing of the Receptor Glycoprotein. J. Virol. 1992, 66, 4028–4039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dveksler, G.S.; Pensiero, M.N.; Dieffenbach, C.W.; Cardellichio, C.B.; Basile, A.A.; Elia, P.E.; Holmes, K.V. Mouse Hepatitis Virus Strain A59 and Blocking Antireceptor Monoclonal Antibody Bind to the N-Terminal Domain of Cellular Receptor. Proc. Natl. Acad. Sci. USA 1993, 90, 1716. [Google Scholar] [CrossRef] [Green Version]
- Rao, P.V.; Kumari, S.; Gallagher, T.M. Identification of a Contiguous 6-Residue Determinant in the MHV Receptor That Controls the Level of Virion Binding to Cells. Virology 1997, 229, 336–348. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.; Zelus, B.D.; Meijers, R.; Liu, J.; Bergelson, J.M.; Duke, N.; Zhang, R.; Joachimiak, A.; Holmes, K.V.; Wang, J. Crystal Structure of Murine SCEACAM1a [1,4]: A Coronavirus Receptor in the CEA Family. EMBO J. 2002, 21, 2076–2086. [Google Scholar] [CrossRef] [Green Version]
- Wessner, D.R.; Shick, P.C.; Lu, J.H.; Cardellichio, C.B.; Gagneten, S.E.; Beauchemin, N.; Holmes, K.V.; Dveksler, G.S. Mutational Analysis of the Virus and Monoclonal Antibody Binding Sites in MHVR, the Cellular Receptor of the Murine Coronavirus Mouse Hepatitis Virus Strain A59. J. Virol. 1998, 72, 1941–1948. [Google Scholar] [CrossRef] [Green Version]
- Peng, G.; Sun, D.; Rajashankar, K.R.; Qian, Z.; Holmes, K.V.; Li, F. Crystal Structure of Mouse Coronavirus Receptor-Binding Domain Complexed with Its Murine Receptor. Proc. Natl. Acad. Sci. USA 2011, 108, 10696–10701. [Google Scholar] [CrossRef] [Green Version]
- Gagneten, S.; Scanga, C.A.; Dveksler, G.S.; Beauchemin, N.; Percy, D.; Holmes, K.V. Attachment Glycoproteins and Receptor Specificity of Rat Coronaviruses. Lab. Anim. Sci. 1996, 46, 159–166. [Google Scholar]
- Godet, M.; Grosclaude, J.; Delmas, B.; Laude, H. Major Receptor-Binding and Neutralization Determinants Are Located within the Same Domain of the Transmissible Gastroenteritis Virus (Coronavirus) Spike Protein. J. Virol. 1994, 68, 8008–8016. [Google Scholar] [CrossRef] [Green Version]
- Schultze, B.; Krempl, C.; Ballesteros, M.L.; Shaw, L.; Schauer, R.; Enjuanes, L.; Herrler, G. Transmissible Gastroenteritis Coronavirus, but Not the Related Porcine Respiratory Coronavirus, Has a Sialic Acid (N-Glycolylneuraminic Acid) Binding Activity. J. Virol. 1996, 70, 5634–5637. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Xu, L.-D.; Zhang, Y.; Cao, H.; Chen, R.; Wang, B.; Huang, Y.-W. Expression of the Human or Porcine C-Type Lectins DC-SIGN/L-SIGN Confers Susceptibility to Porcine Epidemic Diarrhea Virus Entry and Infection in Otherwise Refractory Cell Lines. Microb. Pathog. 2021, 157, 104956. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Ye, G.; Liu, Q.; Navid, M.T.; Zhong, X.; Li, Y.; Wan, C.; Xiao, S.; He, Q.; Fu, Z.F.; et al. Identification and Comparison of Receptor Binding Characteristics of the Spike Protein of Two Porcine Epidemic Diarrhea Virus Strains. Viruses 2016, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.-L. The Role of Cell Surface Sialic Acids for SARS-CoV-2 Infection. Glycobiology 2021. [Google Scholar] [CrossRef]
- Huan, C.; Wang, Y.; Ni, B.; Wang, R.; Huang, L.; Ren, X.; Tong, G.; Ding, C.; Fan, H.; Mao, X. Porcine Epidemic Diarrhea Virus Uses Cell-Surface Heparan Sulfate as an Attachment Factor. Arch. Virol. 2015, 160, 1621–1628. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-X.; Feng, Y.; Wong, G.; Wang, L.; Li, B.; Zhao, X.; Li, Y.; Smaill, F.; Zhang, C. 2008 Identification of Residues in the Receptor-Binding Domain (RBD) of the Spike Protein of Human Coronavirus NL63 That Are Critical for the RBD–ACE2 Receptor Interaction. J. Gen. Virol. 2008, 89, 1015–1024. [Google Scholar] [CrossRef]
- Milewska, A.; Zarebski, M.; Nowak, P.; Stozek, K.; Potempa, J.; Pyrc, K. Human Coronavirus NL63 Utilizes Heparan Sulfate Proteoglycans for Attachment to Target Cells. J. Virol. 2014, 88, 13221–13230. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, H.; Simmons, G.; Rennekamp, A.J.; Chaipan, C.; Gramberg, T.; Heck, E.; Geier, M.; Wegele, A.; Marzi, A.; Bates, P.; et al. Highly Conserved Regions within the Spike Proteins of Human Coronaviruses 229E and NL63 Determine Recognition of Their Respective Cellular Receptors. J. Virol. 2006, 80, 8639–8652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonavia, A.; Zelus, B.D.; Wentworth, D.E.; Talbot, P.J.; Holmes, K.V. Identification of a Receptor-Binding Domain of the Spike Glycoprotein of Human Coronavirus HCoV-229E. J. Virol. 2003, 77, 2530–2538. [Google Scholar] [CrossRef] [Green Version]
- Breslin, J.J.; Mørk, I.; Smith, M.K.; Vogel, L.K.; Hemmila, E.M.; Bonavia, A.; Talbot, P.J.; Sjöström, H.; Norén, O.; Holmes, K.V. Human Coronavirus 229E: Receptor Binding Domain and Neutralization by Soluble Receptor at 37 Degrees C. J. Virol. 2003, 77, 4435–4438. [Google Scholar] [CrossRef] [Green Version]
- Jeffers, S.A.; Hemmila, E.M.; Holmes, K.V. Human Coronavirus 229E Can Use CD209L (L-SIGN) to Enter Cells. Adv. Exp. Med. Biol. 2006, 581, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Haan, C.; Haijema, B.; Schellen, P.; Wichgers Schreur, P.; Lintelo, E.; Vennema, H.; Rottier, P. Cleavage of Group 1 Coronavirus Spike Proteins: How Furin Cleavage Is Traded Off against Heparan Sulfate Binding upon Cell Culture Adaptation. J. Virol. 2008, 82, 6078–6083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regan, A.D.; Whittaker, G.R. Utilization of DC-SIGN for Entry of Feline Coronaviruses into Host Cells. J. Virol. 2008, 82, 11992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regan, A.D.; Millet, J.K.; Tse, L.P.V.; Chillag, Z.; Rinaldi, V.D.; Licitra, B.N.; Dubovi, E.J.; Town, C.D.; Whittaker, G.R. Characterization of a Recombinant Canine Coronavirus with a Distinct Receptor-Binding (S1) Domain. Virology 2012, 430, 90–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, H.; Yamada, Y.K.; Taguchi, F. Localization of Neutralizing Epitopes and the Receptor-Binding Site within the Amino-Terminal 330 Amino Acids of the Murine Coronavirus Spike Protein. J. Virol. 1994, 68, 5403–5410. [Google Scholar] [CrossRef] [Green Version]
- Szczepanski, A.; Owczarek, K.; Bzowska, M.; Gula, K.; Drebot, I.; Ochman, M.; Maksym, B.; Rajfur, Z.; Mitchell, J.A.; Pyrc, K. Canine Respiratory Coronavirus, Bovine Coronavirus, and Human Coronavirus OC43: Receptors and Attachment Factors. Viruses 2019, 11, 328. [Google Scholar] [CrossRef] [Green Version]
- Borucki, M.K.; Allen, J.E.; Chen-Harris, H.; Zemla, A.; Vanier, G.; Mabery, S.; Torres, C.; Hullinger, P.; Slezak, T. The Role of Viral Population Diversity in Adaptation of Bovine Coronavirus to New Host Environments. PLoS ONE 2013, 8, e52752. [Google Scholar] [CrossRef]
- Schultze, B.; Herrler, G. Bovine Coronavirus Uses N-Acetyl-9-O-Acetylneuraminic Acid as a Receptor Determinant to Initiate the Infection of Cultured Cells. J. Gen. Virol. 1992, 73 Pt 4, 901–906. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Lee, P.; Tsang, A.K.L.; Yip, C.C.Y.; Tse, H.; Lee, R.A.; So, L.-Y.; Lau, Y.-L.; Chan, K.-H.; Woo, P.C.Y.; et al. Molecular Epidemiology of Human Coronavirus OC43 Reveals Evolution of Different Genotypes over Time and Recent Emergence of a Novel Genotype Due to Natural Recombination. J. Virol. 2011, 85, 11325–11337. [Google Scholar] [CrossRef] [Green Version]
- Vlasak, R.; Luytjes, W.; Spaan, W.; Palese, P. Human and Bovine Coronaviruses Recognize Sialic Acid-Containing Receptors Similar to Those of Influenza C Viruses. Proc. Natl. Acad. Sci. USA 1988, 85, 4526–4529. [Google Scholar] [CrossRef] [Green Version]
- Mou, H.; Raj, V.S.; van Kuppeveld, F.J.M.; Rottier, P.J.M.; Haagmans, B.L.; Bosch, B.J. The Receptor Binding Domain of the New Middle East Respiratory Syndrome Coronavirus Maps to a 231-Residue Region in the Spike Protein That Efficiently Elicits Neutralizing Antibodies. J. Virol. 2013, 87, 9379–9383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Song, Y.; Xia, K.; He, P.; Zhang, F.; Chen, S.; Pouliot, R.; Weiss, D.J.; Tandon, R.; Bates, J.T.; et al. Heparan Sulfates from Bat and Human Lung and Their Binding to the Spike Protein of SARS-CoV-2 Virus. Carbohydr. Polym. 2021, 260, 117797. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Li, W.; Moore, M.J.; Choe, H.; Farzan, M. A 193-Amino Acid Fragment of the SARS Coronavirus S Protein Efficiently Binds Angiotensin-Converting Enzyme 2. J. Biol. Chem. 2004, 279, 3197–3201. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Chen, C.Z.; Swaroop, M.; Xu, M.; Wang, L.; Lee, J.; Wang, A.Q.; Pradhan, M.; Hagen, N.; Chen, L.; et al. Heparan Sulfate Assists SARS-CoV-2 in Cell Entry and Can Be Targeted by Approved Drugs in Vitro. Cell Discov. 2020, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Marzi, A.; Gramberg, T.; Simmons, G.; Möller, P.; Rennekamp, A.; Krumbiegel, M.; Geier, M.; Eisemann, J.; Turza, N.; Saunier, B.; et al. DC-SIGN and DC-SIGNR Interact with the Glycoprotein of Marburg Virus and the S Protein of Severe Acute Respiratory Syndrome Coronavirus. J. Virol. 2004, 78, 12090–12095. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.-Y.; Huang, Y.; Ganesh, L.; Leung, K.; Kong, W.-P.; Schwartz, O.; Subbarao, K.; Nabel, G.J. PH-Dependent Entry of Severe Acute Respiratory Syndrome Coronavirus Is Mediated by the Spike Glycoprotein and Enhanced by Dendritic Cell Transfer through DC-SIGN. J. Virol. 2004, 78, 5642–5650. [Google Scholar] [CrossRef] [Green Version]
- Tai, W.; He, L.; Zhang, X.; Pu, J.; Voronin, D.; Jiang, S.; Zhou, Y.; Du, L. Characterization of the Receptor-Binding Domain (RBD) of 2019 Novel Coronavirus: Implication for Development of RBD Protein as a Viral Attachment Inhibitor and Vaccine. Cell. Mol. Immunol. 2020, 17, 613–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thépaut, M.; Luczkowiak, J.; Vivès, C.; Labiod, N.; Bally, I.; Lasala, F.; Grimoire, Y.; Fenel, D.; Sattin, S.; Thielens, N.; et al. DC/L-SIGN Recognition of Spike Glycoprotein Promotes SARS-CoV-2 Trans-Infection and Can Be Inhibited by a Glycomimetic Antagonist. PLoS Pathog. 2021, 17, e1009576. [Google Scholar] [CrossRef]
- Huang, X.; Dong, W.; Milewska, A.; Golda, A.; Qi, Y.; Zhu, Q.K.; Marasco, W.A.; Baric, R.S.; Sims, A.C.; Pyrc, K.; et al. Human Coronavirus HKU1 Spike Protein Uses O-Acetylated Sialic Acid as an Attachment Receptor Determinant and Employs Hemagglutinin-Esterase Protein as a Receptor-Destroying Enzyme. J. Virol. 2015, 89, 7202–7213. [Google Scholar] [CrossRef] [Green Version]
- Promkuntod, N.; van Eijndhoven, R.E.W.; de Vrieze, G.; Gröne, A.; Verheije, M.H. Mapping of the Receptor-Binding Domain and Amino Acids Critical for Attachment in the Spike Protein of Avian Coronavirus Infectious Bronchitis Virus. Virology 2014, 448, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Winter, C.; Schwegmann-Weßels, C.; Cavanagh, D.; Neumann, U.; Herrler, G. Sialic Acid Is a Receptor Determinant for Infection of Cells by Avian Infectious Bronchitis Virus. J. Gen. Virol. 2006, 87, 1209–1216. [Google Scholar] [CrossRef]
- Madu, I.G.; Chu, V.C.; Lee, H.; Regan, A.D.; Bauman, B.E.; Whittaker, G.R. Heparan Sulfate Is a Selective Attachment Factor for the Avian Coronavirus Infectious Bronchitis Virus Beaudette. Avian. Dis. 2007, 51, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Buckles, E.; Whittaker, G.R. Expression of the C-Type Lectins DC-SIGN or L-SIGN Alters Host Cell Susceptibility for the Avian Coronavirus, Infectious Bronchitis Virus. Vet. Microbiol. 2012, 157, 285–293. [Google Scholar] [CrossRef]
- Yuan, P.; Yang, Z.; Song, H.; Wang, K.; Yang, Y.; Xie, L.; Huang, S.; Liu, J.; Ran, L.; Song, Z. Three Main Inducers of Alphacoronavirus Infection of Enterocytes: Sialic Acid, Proteases, and Low PH. INT 2018, 61, 53–63. [Google Scholar] [CrossRef]
- Reguera, J.; Santiago, C.; Mudgal, G.; Ordoño, D.; Enjuanes, L.; Casasnovas, J.M. Structural Bases of Coronavirus Attachment to Host Aminopeptidase N and Its Inhibition by Neutralizing Antibodies. PLoS Pathog. 2012, 8, e1002859. [Google Scholar] [CrossRef] [Green Version]
- Tresnan, D.B.; Levis, R.; Holmes, K.V. Feline Aminopeptidase N Serves as a Receptor for Feline, Canine, Porcine, and Human Coronaviruses in Serogroup I. J. Virol. 1996, 70, 8669–8674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, Y.; Xu, W. The Structure and Main Functions of Aminopeptidase N. Curr. Med. Chem. 2007, 14, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Delmas, B.; Gelfi, J.; Kut, E.; Sjöström, H.; Noren, O.; Laude, H. Determinants Essential for the Transmissible Gastroenteritis Virus-Receptor Interaction Reside within a Domain of Aminopeptidase-N That Is Distinct from the Enzymatic Site. J. Virol. 1994, 68, 5216–5224. [Google Scholar] [CrossRef] [Green Version]
- Benbacer, L.; Kut, E.; Besnardeau, L.; Laude, H.; Delmas, B. Interspecies Aminopeptidase-N Chimeras Reveal Species-Specific Receptor Recognition by Canine Coronavirus, Feline Infectious Peritonitis Virus, and Transmissible Gastroenteritis Virus. J. Virol. 1997, 71, 734–737. [Google Scholar] [CrossRef] [Green Version]
- Hegyi, A.; Kolb, A.F. Characterization of Determinants Involved in the Feline Infectious Peritonitis Virus Receptor Function of Feline Aminopeptidase N. J. Gen. Virol. 1998, 79 Pt 6, 1387–1391. [Google Scholar] [CrossRef] [Green Version]
- Tusell, S.M.; Schittone, S.A.; Holmes, K.V. Mutational Analysis of Aminopeptidase N, a Receptor for Several Group 1 Coronaviruses, Identifies Key Determinants of Viral Host Range. J. Virol. 2007, 81, 1261–1273. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, H.; Pyrc, K.; van der Hoek, L.; Geier, M.; Berkhout, B.; Pöhlmann, S. Human Coronavirus NL63 Employs the Severe Acute Respiratory Syndrome Coronavirus Receptor for Cellular Entry. Proc. Natl. Acad. Sci. USA 2005, 102, 7988–7993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-Converting Enzyme 2 Is a Functional Receptor for the SARS Coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhang, C.; Sui, J.; Kuhn, J.H.; Moore, M.J.; Luo, S.; Wong, S.-K.; Huang, I.-C.; Xu, K.; Vasilieva, N.; et al. Receptor and Viral Determinants of SARS-Coronavirus Adaptation to Human ACE2. EMBO J. 2005, 24, 1634–1643. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, L.; Wei, Q.; Zhu, H.; Jiang, H.; Tu, X.; Qin, C.; Chen, Z. Rhesus Angiotensin Converting Enzyme 2 Supports Entry of Severe Acute Respiratory Syndrome Coronavirus in Chinese Macaques. Virology 2008, 381, 89–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Zhang, Y.; Liu, Y.; Chen, Z.; Deng, H.; Ma, Z.; Wang, H.; Hu, Z.; Deng, F. Angiotensin-Converting Enzyme 2 (ACE2) from Raccoon Dog Can Serve as an Efficient Receptor for the Spike Protein of Severe Acute Respiratory Syndrome Coronavirus. J. Gen. Virol. 2009, 90, 2695–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamoto, A.; Taguchi, F.; Fukushi, S.; Morikawa, S.; Yamada, Y.K. Identification of Ferret ACE2 and Its Receptor Function for SARS-Coronavirus. Adv. Exp. Med. Biol. 2006, 581, 519–522. [Google Scholar] [CrossRef] [Green Version]
- Heller, L.K.; Gillim-Ross, L.; Olivieri, E.R.; Wentworth, D.E. Mustela Vison ACE2 Functions as a Receptor for Sars-Coronavirus. Nidoviruses 2006, 581, 507–510. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Chen, D.; Szabla, R.; Zheng, M.; Li, G.; Du, P.; Zheng, S.; Li, X.; Song, C.; Li, R.; et al. Broad and Differential Animal Angiotensin-Converting Enzyme 2 Receptor Usage by SARS-CoV-2. J. Virol. 2020, 94, e00940-20. [Google Scholar] [CrossRef]
- Ge, X.-Y.; Li, J.-L.; Yang, X.-L.; Chmura, A.A.; Zhu, G.; Epstein, J.H.; Mazet, J.K.; Hu, B.; Zhang, W.; Peng, C.; et al. Isolation and Characterization of a Bat SARS-like Coronavirus That Uses the ACE2 Receptor. Nature 2013, 503, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Zhao, X.; Zheng, S.; Chen, D.; Du, P.; Li, X.; Jiang, D.; Guo, J.-T.; Zeng, H.; Lin, H. Bat SARS-Like WIV1 Coronavirus Uses the ACE2 of Multiple Animal Species as Receptor and Evades IFITM3 Restriction via TMPRSS2 Activation of Membrane Fusion. Emerg. Microbes Infect. 2020, 9, 1567–1579. [Google Scholar] [CrossRef]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.W.; Müller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.A.; Zaki, A.; Fouchier, R.A.M.; et al. Dipeptidyl Peptidase 4 Is a Functional Receptor for the Emerging Human Coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babcock, G.J.; Esshaki, D.J.; Thomas, W.D.; Ambrosino, D.M. Amino Acids 270 to 510 of the Severe Acute Respiratory Syndrome Coronavirus Spike Protein Are Required for Interaction with Receptor. J. Virol. 2004, 78, 4552–4560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F. Receptor Recognition Mechanisms of Coronaviruses: A Decade of Structural Studies. J. Virol. 2014, 89, 1954–1964. [Google Scholar] [CrossRef] [Green Version]
- Peng, G.; Xu, L.; Lin, Y.-L.; Chen, L.; Pasquarella, J.R.; Holmes, K.V.; Li, F. Crystal Structure of Bovine Coronavirus Spike Protein Lectin Domain. J. Biol. Chem. 2012, 287, 41931–41938. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Taguchi, F. Analysis of the Receptor-Binding Site of Murine Coronavirus Spike Protein. J. Virol. 1996, 70, 2632–2636. [Google Scholar] [CrossRef] [Green Version]
- Thackray, L.B.; Turner, B.C.; Holmes, K.V. Substitutions of Conserved Amino Acids in the Receptor-Binding Domain of the Spike Glycoprotein Affect Utilization of Murine CEACAM1a by the Murine Coronavirus MHV-A59. Virology 2005, 334, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Mi, D.; Ou, X.; Li, P.; Peng, G.; Liu, Y.; Guo, R.; Mu, Z.; Li, F.; Holmes, K.; Qian, Z. Glycine 29 Is Critical for Conformational Changes of the Spike Glycoprotein of Mouse Hepatitis Virus A59 Triggered by Either Receptor Binding or High PH. J. Virol. 2019, 93, e01046-19. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS Coronavirus Spike Receptor-Binding Domain Complexed with Receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef] [PubMed]
- Hulswit, R.J.G.; Lang, Y.; Bakkers, M.J.G.; Li, W.; Li, Z.; Schouten, A.; Ophorst, B.; Kuppeveld, F.J.M.; van Boons, G.-J.; Bosch, B.-J.; et al. Human Coronaviruses OC43 and HKU1 Bind to 9-O-Acetylated Sialic Acids via a Conserved Receptor-Binding Site in Spike Protein Domain A. Proc. Natl. Acad. Sci. USA 2019, 116, 2681–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tortorici, M.A.; Walls, A.C.; Lang, Y.; Wang, C.; Li, Z.; Koerhuis, D.; Boons, G.-J.; Bosch, B.-J.; Rey, F.A.; de Groot, R.J.; et al. Structural Basis for Human Coronavirus Attachment to Sialic Acid Receptors. Nat. Struct. Mol. Biol. 2019, 26, 481–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, C.M.; Gebauer, F.; Suñé, C.; Mendez, A.; Dopazo, J.; Enjuanes, L. Genetic Evolution and Tropism of Transmissible Gastroenteritis Coronaviruses. Virology 1992, 190, 92–105. [Google Scholar] [CrossRef]
- Krempl, C.; Ballesteros, M.L.; Zimmer, G.; Enjuanes, L.; Klenk, H.D.; Herrler, G. Characterization of the Sialic Acid Binding Activity of Transmissible Gastroenteritis Coronavirus by Analysis of Haemagglutination-Deficient Mutants. J. Gen. Virol. 2000, 81, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Schwegmann-Weßels, C.; Zimmer, G.; Laude, H.; Enjuanes, L.; Herrler, G. Binding of Transmissible Gastroenteritis Coronavirus to Cell Surface Sialoglycoproteins. J. Virol. 2002, 76, 6037–6043. [Google Scholar] [CrossRef] [Green Version]
- Schwegmann-Wessels, C.; Zimmer, G.; Schröder, B.; Breves, G.; Herrler, G. Binding of Transmissible Gastroenteritis Coronavirus to Brush Border Membrane Sialoglycoproteins. J. Virol. 2003, 77, 11846–11848. [Google Scholar] [CrossRef] [Green Version]
- Künkel, F.; Herrler, G. Structural and Functional Analysis of the Surface Protein of Human Coronavirus OC43. Virology 1993, 195, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R. Human Coronavirus OC43 Interacts with Major Histocompatibility Complex Class I Molecules at the Cell Surface to Establish Infection. Immunol. Investig. 1994, 23, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Winter, C.; Herrler, G.; Neumann, U. Infection of the Tracheal Epithelium by Infectious Bronchitis Virus Is Sialic Acid Dependent. Microbes Infect. 2008, 10, 367. [Google Scholar] [CrossRef]
- Li, W.; Hulswit, R.J.G.; Widjaja, I.; Raj, V.S.; McBride, R.; Peng, W.; Widagdo, W.; Tortorici, M.A.; van Dieren, B.; Lang, Y.; et al. Identification of Sialic Acid-Binding Function for the Middle East Respiratory Syndrome Coronavirus Spike Glycoprotein. Proc. Natl. Acad. Sci. USA 2017, 114, E8508–E8517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Thorp, S.C. Cell Surface Heparan Sulfate and Its Roles in Assisting Viral Infections. Med. Res. Rev. 2002, 22, 1–25. [Google Scholar] [CrossRef]
- De Haan, C.A.M.; Li, Z.; Lintelo, E.; te Bosch, B.J.; Haijema, B.J.; Rottier, P.J.M. Murine Coronavirus with an Extended Host Range Uses Heparan Sulfate as an Entry Receptor. J. Virol. 2005, 79, 14451. [Google Scholar] [CrossRef] [Green Version]
- Schickli, J.H.; Zelus, B.D.; Wentworth, D.E.; Sawicki, S.G.; Holmes, K.V. The Murine Coronavirus Mouse Hepatitis Virus Strain A59 from Persistently Infected Murine Cells Exhibits an Extended Host Range. J. Virol. 1997, 71. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, R.; Sawicki, S.G.; Taguchi, F. Heparan Sulfate Is a Binding Molecule but Not a Receptor for CEACAM1-Independent Infection of Murine Coronavirus. Virology 2007, 366, 16–22. [Google Scholar] [CrossRef]
- Baribaud, F.; Pöhlmann, S.; Doms, R.W. The Role of DC-SIGN and DC-SIGNR in HIV and SIV Attachment, Infection, and Transmission. Virology 2001, 286, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, P.; Manuel, S.L.; Khan, Z.K.; Ahuja, J.; Quann, K.; Wigdahl, B. DC-SIGN Mediates Cell-Free Infection and Transmission of Human T-Cell Lymphotropic Virus Type 1 by Dendritic Cells. J. Virol. 2009, 83, 10908–10921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozach, P.-Y.; Burleigh, L.; Staropoli, I.; Amara, A. The C Type Lectins DC-SIGN and L-SIGN. Glycovirology Protoc. 2007, 379, 51–68. [Google Scholar] [CrossRef]
- Schneider-Schaulies, J.; Schneider-Schaulies, S. Receptor Interactions, Tropism, and Mechanisms Involved in Morbillivirus-Induced Immunomodulation. Adv. Virus Res. 2008, 71, 173–205. [Google Scholar] [CrossRef] [PubMed]
- Cambi, A.; Koopman, M.; Figdor, C.G. How C-Type Lectins Detect Pathogens. Cell. Microbiol. 2005, 7, 481–488. [Google Scholar] [CrossRef]
- Geijtenbeek, T.B.H.; Gringhuis, S.I. Signalling through C-Type Lectin Receptors: Shaping Immune Responses. Nat. Rev. Immunol. 2009, 9, 465–479. [Google Scholar] [CrossRef]
- Cambi, A.; de Lange, F.; van Maarseveen, N.M.; Nijhuis, M.; Joosten, B.; van Dijk, E.M.H.P.; de Bakker, B.I.; Fransen, J.A.M.; Bovee-Geurts, P.H.M.; van Leeuwen, F.N.; et al. Microdomains of the C-Type Lectin DC-SIGN Are Portals for Virus Entry into Dendritic Cells. J. Cell Biol. 2004, 164, 145–155. [Google Scholar] [CrossRef]
- Han, D.P.; Lohani, M.; Cho, M.W. Specific Asparagine-Linked Glycosylation Sites Are Critical for DC-SIGN- and L-SIGN-Mediated Severe Acute Respiratory Syndrome Coronavirus Entry. J. Virol. 2007, 81, 12029–12039. [Google Scholar] [CrossRef] [Green Version]
- Jeffers, S.A.; Tusell, S.M.; Gillim-Ross, L.; Hemmila, E.M.; Achenbach, J.E.; Babcock, G.J.; Thomas, W.D.; Thackray, L.B.; Young, M.D.; Mason, R.J.; et al. CD209L (L-SIGN) Is a Receptor for Severe Acute Respiratory Syndrome Coronavirus. Proc. Natl. Acad. Sci. USA 2004, 101, 15748–15753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gramberg, T.; Hofmann, H.; Möller, P.; Lalor, P.F.; Marzi, A.; Geier, M.; Krumbiegel, M.; Winkler, T.; Kirchhoff, F.; Adams, D.H.; et al. LSECtin Interacts with Filovirus Glycoproteins and the Spike Protein of SARS Coronavirus. Virology 2005, 340, 224–236. [Google Scholar] [CrossRef] [Green Version]
- Guillén, J.; Pérez-Berná, A.J.; Moreno, M.R.; Villalaín, J. Identification of the Membrane-Active Regions of the Severe Acute Respiratory Syndrome Coronavirus Spike Membrane Glycoprotein Using a 16/18-Mer Peptide Scan: Implications for the Viral Fusion Mechanism. J. Virol. 2005, 79, 1743–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.M.; Rottier, P.J.M. The Coronavirus Spike Protein Is a Class I Virus Fusion Protein: Structural and Functional Characterization of the Fusion Core Complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Weiss, S.R. Roles in Cell-to-Cell Fusion of Two Conserved Hydrophobic Regions in the Murine Coronavirus Spike Protein. Virology 1998, 244, 483–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petit, C.M.; Melancon, J.M.; Chouljenko, V.N.; Colgrove, R.; Farzan, M.; Knipe, D.M.; Kousoulas, K.G. Genetic Analysis of the SARS-Coronavirus Spike Glycoprotein Functional Domains Involved in Cell-Surface Expression and Cell-to-Cell Fusion. Virology 2005, 341, 215–230. [Google Scholar] [CrossRef] [Green Version]
- De Groot, R.J.; Luytjes, W.; Horzinek, M.C.; van der Zeijst, B.A.; Spaan, W.J.; Lenstra, J.A. Evidence for a Coiled-Coil Structure in the Spike Proteins of Coronaviruses. J. Mol. Biol. 1987, 196, 963–966. [Google Scholar] [CrossRef]
- Tripet, B.; Howard, M.W.; Jobling, M.; Holmes, R.K.; Holmes, K.V.; Hodges, R.S. Structural Characterization of the SARS-Coronavirus Spike S Fusion Protein Core. J. Biol. Chem. 2004, 279, 20836–20849. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Liu, Y.; Lou, Z.; Qin, L.; Li, X.; Bai, Z.; Pang, H.; Tien, P.; Gao, G.F.; Rao, Z. Structural Basis for Coronavirus-Mediated Membrane Fusion: Crystal structure of mouse hepatitis virus spike protein fusion core. J. Biol. Chem. 2004, 279, 30514–30522. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Deng, Y.; Liu, J.; van der Hoek, L.; Berkhout, B.; Lu, M. Core Structure of S2 from the Human Coronavirus NL63 Spike Glycoprotein. Biochemistry 2006, 45, 15205–15215. [Google Scholar] [CrossRef] [PubMed]
- Eckert, D.M.; Kim, P.S. Mechanisms of Viral Membrane Fusion and Its Inhibition. Annu. Rev. Biochem. 2001, 70, 777–810. [Google Scholar] [CrossRef] [Green Version]
- Epand, R.M. Fusion Peptides and the Mechanism of Viral Fusion. Biochim. Biophys. Acta 2003, 1614, 116–121. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Xiao, G.; Chen, Y.; He, Y.; Niu, J.; Escalante, C.R.; Xiong, H.; Farmar, J.; Debnath, A.K.; Tien, P.; et al. Interaction between Heptad Repeat 1 and 2 Regions in Spike Protein of SARS-Associated Coronavirus: Implications for Virus Fusogenic Mechanism and Identification of Fusion Inhibitors. Lancet 2004, 363, 938–947. [Google Scholar] [CrossRef] [Green Version]
- Sainz, B.; Mossel, E.C.; Gallaher, W.R.; Wimley, W.C.; Peters, C.J.; Wilson, R.B.; Garry, R.F. Inhibition of Severe Acute Respiratory Syndrome-Associated Coronavirus (SARS-CoV) Infectivity by Peptides Analogous to the Viral Spike Protein. Virus Res. 2006, 120, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Tripet, B.; Kao, D.; Jeffers, S.; Holmes, K.; Hodges, R. Template-Based Coiled-Coil Antigens Elicit Neutralizing Antibodies to the SARS-Coronavirus. J. Struct. Biol. 2006, 155, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Howard, M.W.; Travanty, E.A.; Jeffers, S.A.; Smith, M.K.; Wennier, S.T.; Thackray, L.B.; Holmes, K.V. Aromatic Amino Acids in the Juxtamembrane Domain of Severe Acute Respiratory Syndrome Coronavirus Spike Glycoprotein Are Important for Receptor-Dependent Virus Entry and Cell-Cell Fusion. J. Virol. 2008, 82, 2883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bos, E.C.; Heijnen, L.; Luytjes, W.; Spaan, W.J. Mutational Analysis of the Murine Coronavirus Spike Protein: Effect on Cell-to-Cell Fusion. Virology 1995, 214, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Broer, R.; Boson, B.; Spaan, W.; Cosset, F.-L.; Corver, J. Important Role for the Transmembrane Domain of Severe Acute Respiratory Syndrome Coronavirus Spike Protein during Entry. J. Virol. 2006, 80, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.W.; Sheng, Y.; Gombold, J.L. Coronavirus-Induced Membrane Fusion Requires the Cysteine-Rich Domain in the Spike Protein. Virology 2000, 269, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Hogue, B.G.; Machamer, C.E. Coronavirus Structural Proteins and Virus Assembly. In Nidoviruses; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2007; pp. 179–200. [Google Scholar]
- Youn, S.; Collisson, E.W.; Machamer, C.E. Contribution of Trafficking Signals in the Cytoplasmic Tail of the Infectious Bronchitis Virus Spike Protein to Virus Infection. J. Virol. 2005, 79, 13209–13217. [Google Scholar] [CrossRef] [Green Version]
- Bonifacino, J.S.; Jackson, C.L. Endosome-Specific Localization and Function of the ARF Activator GNOM. Cell 2003, 112, 141–142. [Google Scholar] [CrossRef] [Green Version]
- Schwegmann-Wessels, C.; Glende, J.; Ren, X.; Qu, X.; Deng, H.; Enjuanes, L.; Herrler, G. Comparison of Vesicular Stomatitis Virus Pseudotyped with the S Proteins from a Porcine and a Human Coronavirus. J. Gen. Virol. 2009, 90, 1724–1729. [Google Scholar] [CrossRef]
- Lin, H.-X.; Feng, Y.; Tu, X.; Zhao, X.; Hsieh, C.-H.; Griffin, L.; Junop, M.; Zhang, C. Characterization of the Spike Protein of Human Coronavirus NL63 in Receptor Binding and Pseudotype Virus Entry. Virus Res. 2011, 160, 283–293. [Google Scholar] [CrossRef]
- Giroglou, T.; Cinatl, J.; Rabenau, H.; Drosten, C.; Schwalbe, H.; Doerr, H.W.; von Laer, D. Retroviral Vectors Pseudotyped with Severe Acute Respiratory Syndrome Coronavirus S Protein. J. Virol. 2004, 78, 9007–9015. [Google Scholar] [CrossRef] [Green Version]
- Petit, C.M.; Chouljenko, V.N.; Iyer, A.; Colgrove, R.; Farzan, M.; Knipe, D.M.; Kousoulas, K.G. Palmitoylation of the Cysteine-Rich Endodomain of the SARS-Coronavirus Spike Glycoprotein Is Important for Spike-Mediated Cell Fusion. Virology 2007, 360, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Thorp, E.B.; Boscarino, J.A.; Logan, H.L.; Goletz, J.T.; Gallagher, T.M. Palmitoylations on Murine Coronavirus Spike Proteins Are Essential for Virion Assembly and Infectivity. J. Virol. 2006, 80, 1280–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Haan, C.A.M.; Rottier, P.J.M. Molecular Interactions in the Assembly of Coronaviruses. Adv. Virus Res. 2005, 64, 165–230. [Google Scholar] [CrossRef]
- De Haan, C.A.M.; Stadler, K.; Godeke, G.-J.; Bosch, B.J.; Rottier, P.J.M. Cleavage Inhibition of the Murine Coronavirus Spike Protein by a Furin-like Enzyme Affects Cell-Cell but Not Virus-Cell Fusion. J. Virol. 2004, 78, 6048–6054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stauber, R.; Pfleiderera, M.; Siddell, S. Proteolytic Cleavage of the Murine Coronavirus Surface Glycoprotein Is Not Required for Fusion Activity. J. Gen. Virol. 1993, 74 Pt 2, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, F. Fusion Formation by the Uncleaved Spike Protein of Murine Coronavirus JHMV Variant Cl-2. J. Virol. 1993, 67, 1195. [Google Scholar] [CrossRef] [Green Version]
- Wicht, O.; Burkard, C.; de Haan, C.A.M.; van Kuppeveld, F.J.M.; Rottier, P.J.M.; Bosch, B.J. Identification and Characterization of a Proteolytically Primed Form of the Murine Coronavirus Spike Proteins after Fusion with the Target Cell. J. Virol. 2014, 88, 4943–4952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, Y.; Liu, D.X. Proteolytic Activation of the Spike Protein at a Novel RRRR/S Motif Is Implicated in Furin-Dependent Entry, Syncytium Formation, and Infectivity of Coronavirus Infectious Bronchitis Virus in Cultured Cells. J. Virol. 2009, 83, 8744–8758. [Google Scholar] [CrossRef] [Green Version]
- Wicht, O.; Li, W.; Willems, L.; Meuleman, T.J.; Wubbolts, R.W.; van Kuppeveld, F.J.M.; Rottier, P.J.M.; Bosch, B.J. Proteolytic Activation of the Porcine Epidemic Diarrhea Coronavirus Spike Fusion Protein by Trypsin in Cell Culture. J. Virol. 2014, 88, 7952–7961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-E.; Cruz, D.J.M.; Shin, H.-J. Clathrin- and Serine Proteases-Dependent Uptake of Porcine Epidemic Diarrhea Virus into Vero Cells. Virus Res. 2014, 191, 21–29. [Google Scholar] [CrossRef]
- Regan, A.D.; Shraybman, R.; Cohen, R.D.; Whittaker, G.R. Differential Role for Low PH and Cathepsin-Mediated Cleavage of the Viral Spike Protein during Entry of Serotype II Feline Coronaviruses. Vet. Microbiol. 2008, 132, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Glowacka, I.; Bertram, S.; Müller, M.A.; Allen, P.; Soilleux, E.; Pfefferle, S.; Steffen, I.; Tsegaye, T.S.; He, Y.; Gnirss, K.; et al. Evidence That TMPRSS2 Activates the Severe Acute Respiratory Syndrome Coronavirus Spike Protein for Membrane Fusion and Reduces Viral Control by the Humoral Immune Response. J. Virol. 2011, 85, 4122–4134. [Google Scholar] [CrossRef] [Green Version]
- Shirato, K.; Kawase, M.; Matsuyama, S. Middle East Respiratory Syndrome Coronavirus Infection Mediated by the Transmembrane Serine Protease TMPRSS2. J. Virol. 2013, 87, 12552–12561. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Kao, R.Y.; Zhou, Y.; He, Y.; Zhao, G.; Wong, C.; Jiang, S.; Yuen, K.-Y.; Jin, D.-Y.; Zheng, B.-J. Cleavage of Spike Protein of SARS Coronavirus by Protease Factor Xa Is Associated with Viral Infectivity. Biochem. Biophys. Res. Commun. 2007, 359, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Ujike, M.; Morikawa, S.; Tashiro, M.; Taguchi, F. Protease-Mediated Enhancement of Severe Acute Respiratory Syndrome Coronavirus Infection. Proc. Natl. Acad. Sci. USA 2005, 102, 12543–12547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, G.; Gosalia, D.N.; Rennekamp, A.J.; Reeves, J.D.; Diamond, S.L.; Bates, P. Inhibitors of Cathepsin L Prevent Severe Acute Respiratory Syndrome Coronavirus Entry. Proc. Natl. Acad. Sci. USA 2005, 102, 11876–11881. [Google Scholar] [CrossRef] [Green Version]
- Kawase, M.; Shirato, K.; Matsuyama, S.; Taguchi, F. Protease-Mediated Entry via the Endosome of Human Coronavirus 229E. J. Virol. 2009, 83, 712–721. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, R.; Matsuyama, S.; Shirato, K.; Maejima, M.; Fukushi, S.; Morikawa, S.; Taguchi, F. Entry from the Cell Surface of Severe Acute Respiratory Syndrome Coronavirus with Cleaved S Protein as Revealed by Pseudotype Virus Bearing Cleaved S Protein. J. Virol. 2008, 82, 11985–11991. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Hingley, S.; Simmons, G.; Yu, C.; Sarma, J.; Bates, P.; Weiss, S. Endosomal Proteolysis by Cathepsins Is Necessary for Murine Coronavirus Mouse Hepatitis Virus Type 2 Spike-Mediated Entry. J. Virol. 2006, 80, 5768–5776. [Google Scholar] [CrossRef] [Green Version]
- Bailey, C.C.; Zhong, G.; Huang, I.-C.; Farzan, M. IFITM-Family Proteins: The Cell’s First Line of Antiviral Defense. Annu. Rev. Virol. 2014, 1, 261–283. [Google Scholar] [CrossRef] [Green Version]
- Perreira, J.M.; Chin, C.R.; Feeley, E.M.; Brass, A.L. IFITMs Restrict the Replication of Multiple Pathogenic Viruses. J. Mol. Biol. 2013, 425, 4937–4955. [Google Scholar] [CrossRef]
- Zhao, X.; Li, J.; Winkler, C.A.; An, P.; Guo, J.-T. IFITM Genes, Variants, and Their Roles in the Control and Pathogenesis of Viral Infections. Front. Microbiol. 2019, 9, 3228. [Google Scholar] [CrossRef] [Green Version]
- Huang, I.-C.; Bailey, C.C.; Weyer, J.L.; Radoshitzky, S.R.; Becker, M.M.; Chiang, J.J.; Brass, A.L.; Ahmed, A.A.; Chi, X.; Dong, L.; et al. Distinct Patterns of IFITM-Mediated Restriction of Filoviruses, SARS Coronavirus, and Influenza A Virus. PLoS Pathog 2011, 7, e1001258. [Google Scholar] [CrossRef]
- Winstone, H.; Lista, M.J.; Reid, A.C.; Bouton, C.; Pickering, S.; Galao, R.P.; Kerridge, C.; Doores, K.J.; Swanson, C.M.; Neil, S.J.D. The Polybasic Cleavage Site in SARS-CoV-2 Spike Modulates Viral Sensitivity to Type I Interferon and IFITM2. J. Virol. 2021, 95, e02422-20. [Google Scholar] [CrossRef]
- Bertram, S.; Dijkman, R.; Habjan, M.; Heurich, A.; Gierer, S.; Glowacka, I.; Welsch, K.; Winkler, M.; Schneider, H.; Hofmann-Winkler, H.; et al. TMPRSS2 Activates the Human Coronavirus 229E for Cathepsin-Independent Host Cell Entry and Is Expressed in Viral Target Cells in the Respiratory Epithelium. J. Virol. 2013, 87, 6150–6160. [Google Scholar] [CrossRef] [Green Version]
- Wrensch, F.; Winkler, M.; Pöhlmann, S. IFITM Proteins Inhibit Entry Driven by the MERS-Coronavirus Spike Protein: Evidence for Cholesterol-Independent Mechanisms. Viruses 2014, 6, 3683–3698. [Google Scholar] [CrossRef] [Green Version]
- Shi, G.; Kenney, A.D.; Kudryashova, E.; Zani, A.; Zhang, L.; Lai, K.K.; Hall-Stoodley, L.; Robinson, R.T.; Kudryashov, D.S.; Compton, A.A.; et al. Opposing Activities of IFITM Proteins in SARS-CoV-2 Infection. EMBO J. 2021, 40, e106501. [Google Scholar] [CrossRef] [PubMed]
- Buchrieser, J.; Dufloo, J.; Hubert, M.; Monel, B.; Planas, D.; Rajah, M.M.; Planchais, C.; Porrot, F.; Guivel-Benhassine, F.; van der Werf, S.; et al. Syncytia Formation by SARS-CoV-2-Infected Cells. EMBO J. 2020, 39, e106267. [Google Scholar] [CrossRef]
- Zhao, X.; Zheng, S.; Chen, D.; Zheng, M.; Li, X.; Li, G.; Lin, H.; Chang, J.; Zeng, H.; Guo, J.-T. LY6E Restricts Entry of Human Coronaviruses, Including Currently Pandemic SARS-CoV-2. J. Virol. 2020, 94, e00562-20. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Sehgal, M.; Hou, Z.; Cheng, J.; Shu, S.; Wu, S.; Guo, F.; Le Marchand, S.J.; Lin, H.; Chang, J.; et al. Identification of Residues Controlling Restriction versus Enhancing Activities of IFITM Proteins on Entry of Human Coronaviruses. J. Virol. 2018, 92, e01535-17. [Google Scholar] [CrossRef] [Green Version]
- Pfaender, S.; Mar, K.B.; Michailidis, E.; Kratzel, A.; Boys, I.N.; V’kovski, P.; Fan, W.; Kelly, J.N.; Hirt, D.; Ebert, N.; et al. LY6E Impairs Coronavirus Fusion and Confers Immune Control of Viral Disease. Nat. Microbiol. 2020, 5, 1330–1339. [Google Scholar] [CrossRef]
- Bruchez, A.; Sha, K.; Johnson, J.; Chen, L.; Stefani, C.; McConnell, H.; Gaucherand, L.; Prins, R.; Matreyek, K.A.; Hume, A.J.; et al. MHC Class II Transactivator CIITA Induces Cell Resistance to Ebola Virus and SARS-like Coronaviruses. Science 2020, 370, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Millet, J.K.; Kien, F.; Cheung, C.-Y.; Siu, Y.-L.; Chan, W.-L.; Li, H.; Leung, H.-L.; Jaume, M.; Bruzzone, R.; Peiris, J.S.M.; et al. Ezrin Interacts with the SARS Coronavirus Spike Protein and Restrains Infection at the Entry Stage. PLoS ONE 2012, 7, e49566. [Google Scholar] [CrossRef]
- Chen, D.; Hou, Z.; Jiang, D.; Zheng, M.; Li, G.; Zhang, Y.; Li, R.; Lin, H.; Chang, J.; Zeng, H.; et al. GILT Restricts the Cellular Entry Mediated by the Envelope Glycoproteins of SARS-CoV, Ebola Virus and Lassa Fever Virus. Emerg. Microbes Infect. 2019, 8, 1511–1523. [Google Scholar] [CrossRef] [Green Version]
- Zang, R.; Case, J.B.; Yutuc, E.; Ma, X.; Shen, S.; Gomez Castro, M.F.; Liu, Z.; Zeng, Q.; Zhao, H.; Son, J.; et al. Cholesterol 25-Hydroxylase Suppresses SARS-CoV-2 Replication by Blocking Membrane Fusion. Proc. Natl. Acad. Sci. USA 2020, 117, 32105–32113. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, W.; Hui, H.; Tiwari, S.K.; Zhang, Q.; Croker, B.A.; Rawlings, S.; Smith, D.; Carlin, A.F.; Rana, T.M. Cholesterol 25-Hydroxylase Inhibits SARS-CoV-2 and Other Coronaviruses by Depleting Membrane Cholesterol. EMBO J. 2020, 39, e106057. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J. Interferon-Stimulated Genes: What Do They All Do? Annu. Rev. Virol. 2019, 6, 567–584. [Google Scholar] [CrossRef]
- Zhou, P.; Fan, H.; Lan, T.; Yang, X.-L.; Shi, W.-F.; Zhang, W.; Zhu, Y.; Zhang, Y.-W.; Xie, Q.-M.; Mani, S.; et al. Fatal Swine Acute Diarrhoea Syndrome Caused by an HKU2-Related Coronavirus of Bat Origin. Nature 2018, 556, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Edwards, C.E.; Yount, B.L.; Graham, R.L.; Leist, S.R.; Hou, Y.J.; Dinnon, K.H.; Sims, A.C.; Swanstrom, J.; Gully, K.; Scobey, T.D.; et al. Swine Acute Diarrhea Syndrome Coronavirus Replication in Primary Human Cells Reveals Potential Susceptibility to Infection. Proc. Natl. Acad. Sci. USA 2020, 117, 26915–26925. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genera | Species * | Receptor | Receptor Binding Domain (RBD) | Attachment Factor |
|---|---|---|---|---|
| Alphacoronaviruses | TGEV | Porcine APN | C-terminal aa 506–655 [23] | Sialic acid [24], DC/L-SIGN [25] |
| PEDV | Porcine APN | C-terminal [26] | Sialic acid [27], heparan sulfate [28], DC/L-SIGN [25] | |
| HCoV-NL63 | ACE2 | C-terminal aa 476–616 [29] | Heparan sulfate [30], DC/L-SIGN [31] | |
| HCoV-229E | Human APN (also called CD13) | C-terminal aa 407–547 [32,33] | DC/L-SIGN [34] | |
| FCoV | Feline APN | Unknown | Heparan sulfate [35], DC/L-SIGN [36] | |
| CCoV | Canine APN | C-terminal aa 526–676 [37] | ||
| Betacoronavirus | MHV | CEACAM1 (also called CD66a) | N-terminal aa 1–330 [38] | Sialic acid [27], Heparan sulfate [39] |
| BCoV | Unknown | Not yet determined but falls within the N-terminal domain [40] | Sialic acid [41], heparan sulfate [39] | |
| HCoV-OC43 | Unknown | aa 339–549 [42] | Sialic acid [43], heparan sulfate [35] | |
| MERS-CoV | DPP4 (also called CD26) | aa 358–588 [44] | Sialic acid [27], heparan sulfate [45] | |
| SARS-CoV | ACE2 | aa 318–510 [46] | Heparan sulfate [47], DC/L-SIGN [48,49] | |
| SARS-CoV-2 | ACE2 | aa 331–524 [50] | Heparan sulfate [47], DC/L-SIGN [51] | |
| HCoV-HKU1 | Unknown | N/A | O-Acetylated Sialic Acid [52] | |
| Gammacoronavirus | IBV | Unknown | N-terminal residues 19–272 [53] | Sialic acid [54], Heparan sulfate [55], DC/L-SIGN [56] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Y.-R.; Li, X.; Zhao, X.; Lin, H. Cell Entry of Animal Coronaviruses. Viruses 2021, 13, 1977. https://doi.org/10.3390/v13101977
Cheng Y-R, Li X, Zhao X, Lin H. Cell Entry of Animal Coronaviruses. Viruses. 2021; 13(10):1977. https://doi.org/10.3390/v13101977
Chicago/Turabian StyleCheng, Yang-Ran, Xinglin Li, Xuesen Zhao, and Hanxin Lin. 2021. "Cell Entry of Animal Coronaviruses" Viruses 13, no. 10: 1977. https://doi.org/10.3390/v13101977
APA StyleCheng, Y.-R., Li, X., Zhao, X., & Lin, H. (2021). Cell Entry of Animal Coronaviruses. Viruses, 13(10), 1977. https://doi.org/10.3390/v13101977