
SDAV, the Rat Coronavirus—How Much Do We Know about It in the Light of Potential Zoonoses


Abstract
:1. Introduction
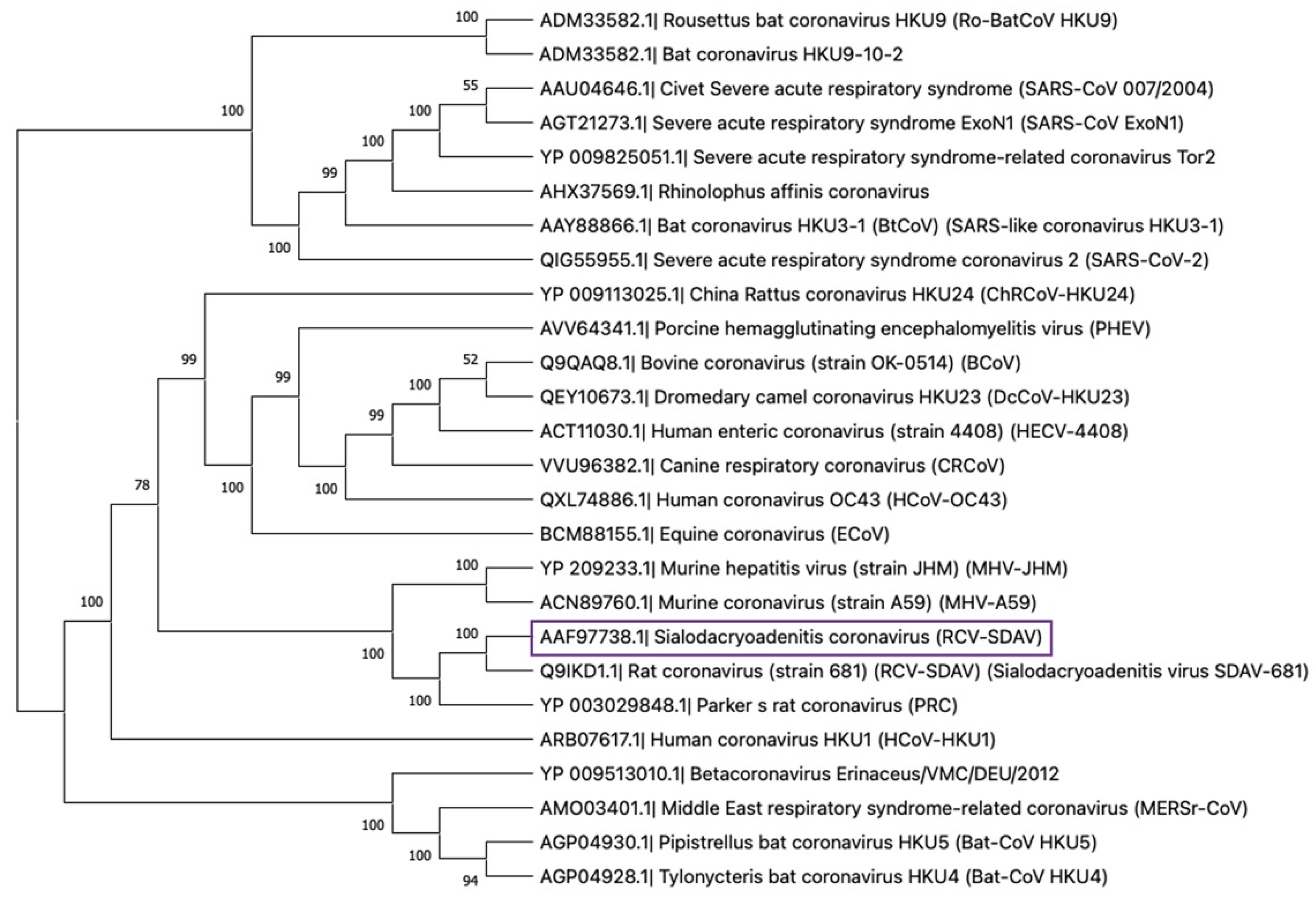
2. Rat Coronaviruses: The Origin
2.1. Discovery
2.2. Sialodacryoadenitis Virus: Transmission, Clinical Signs of Infection, and Diagnosis
3. Characteristics
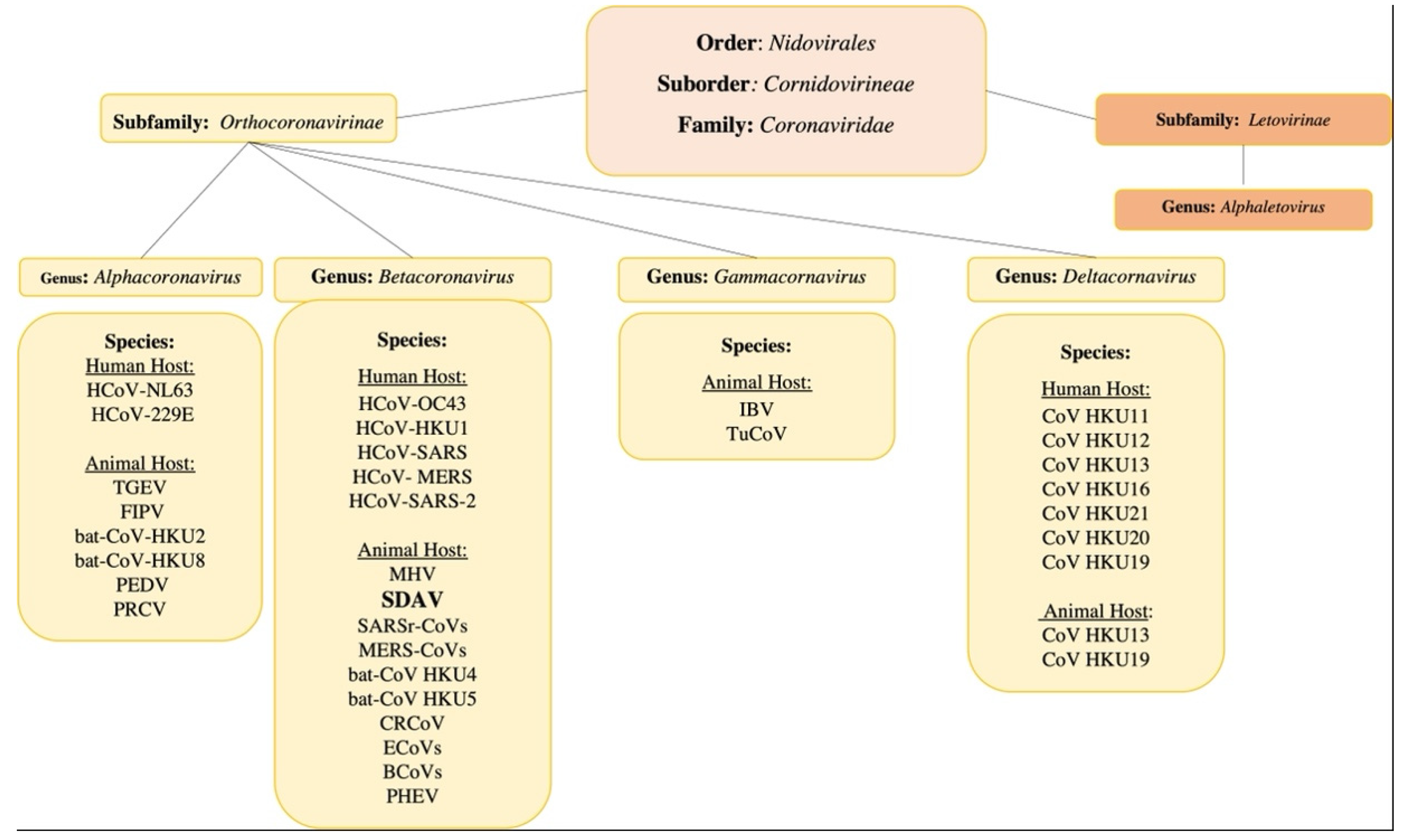
3.1. The General Classification
3.2. The Rat Coronaviruses
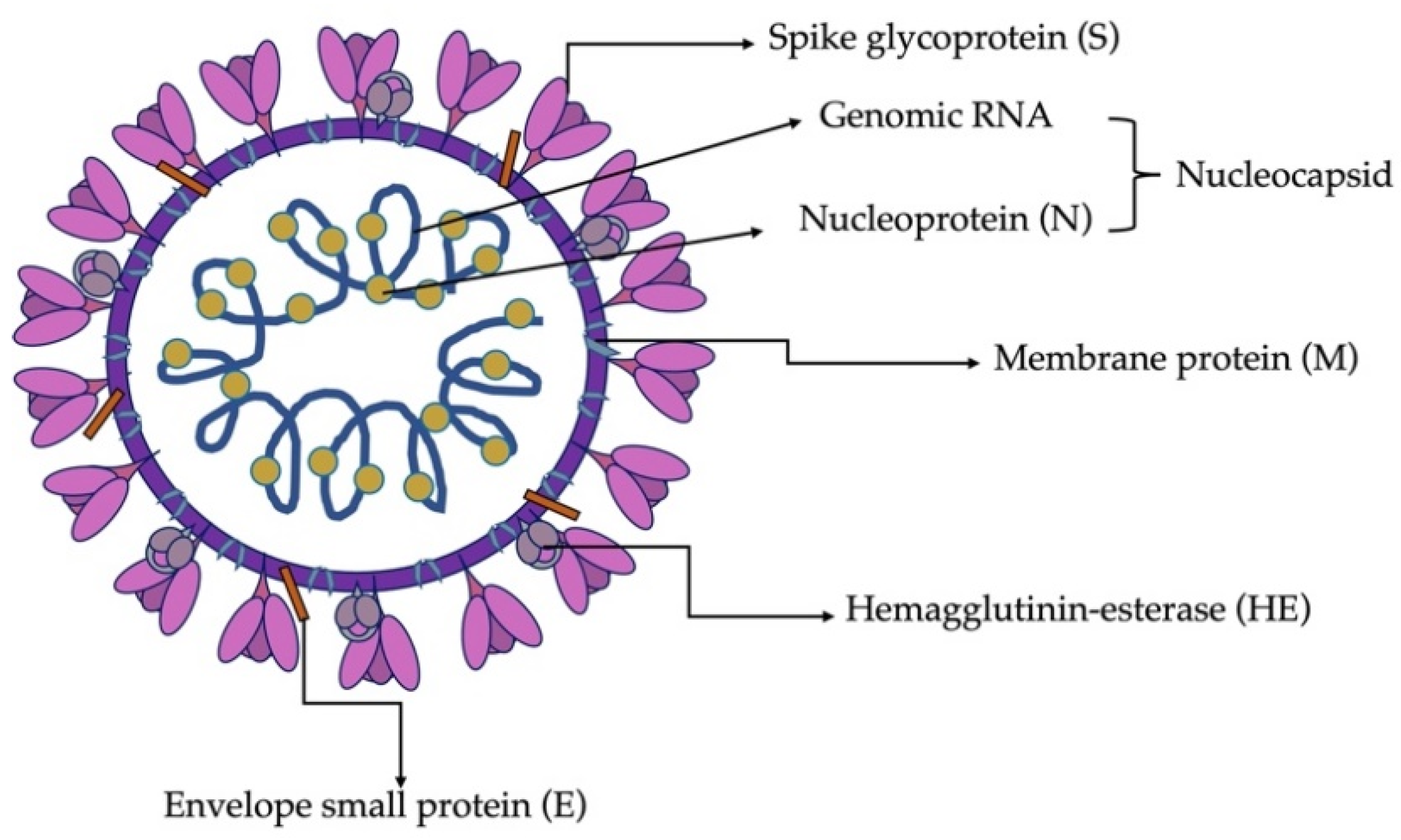
3.3. Virion Structure and Biological Functions of Proteins
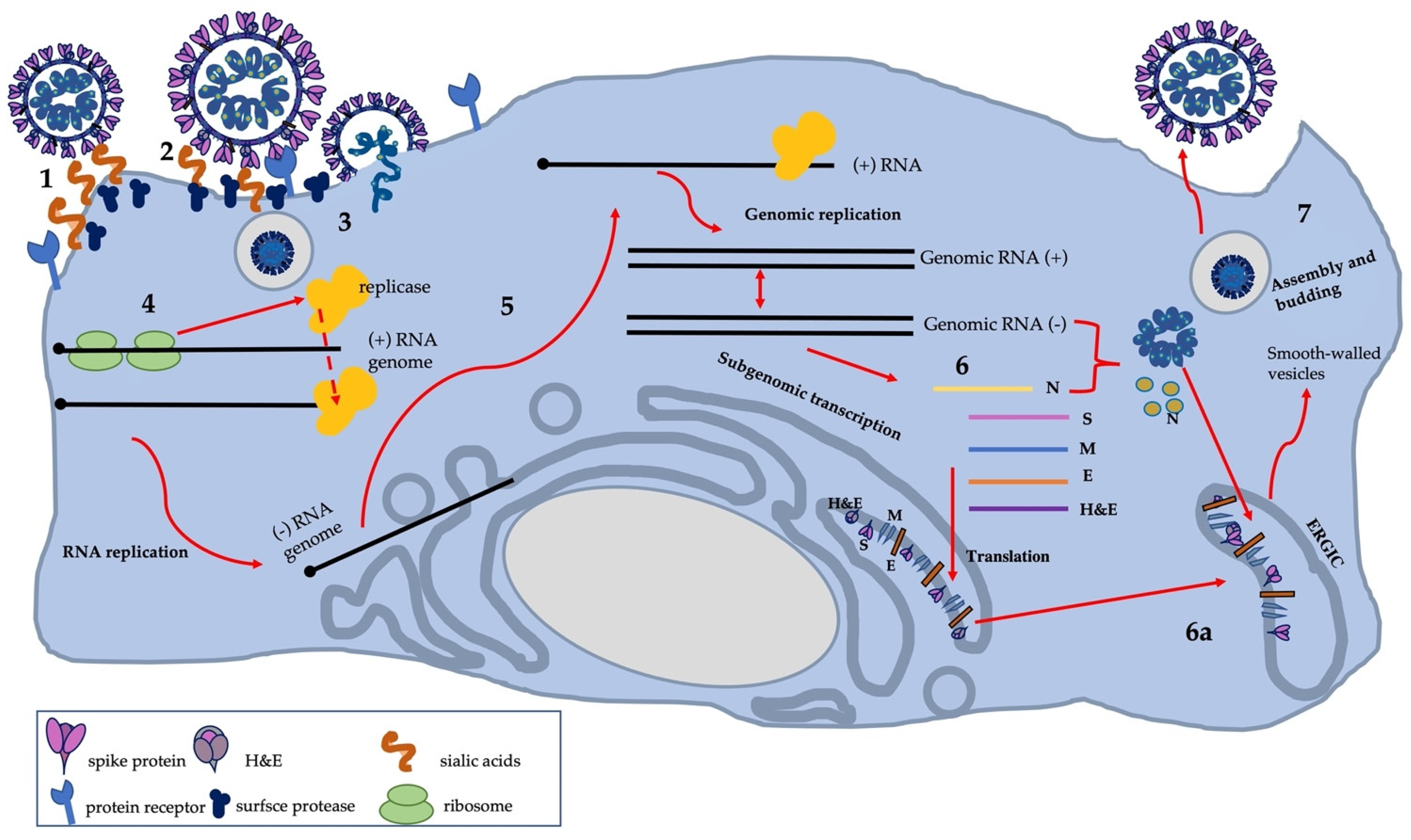
3.4. Cell Infection and Virus Replication
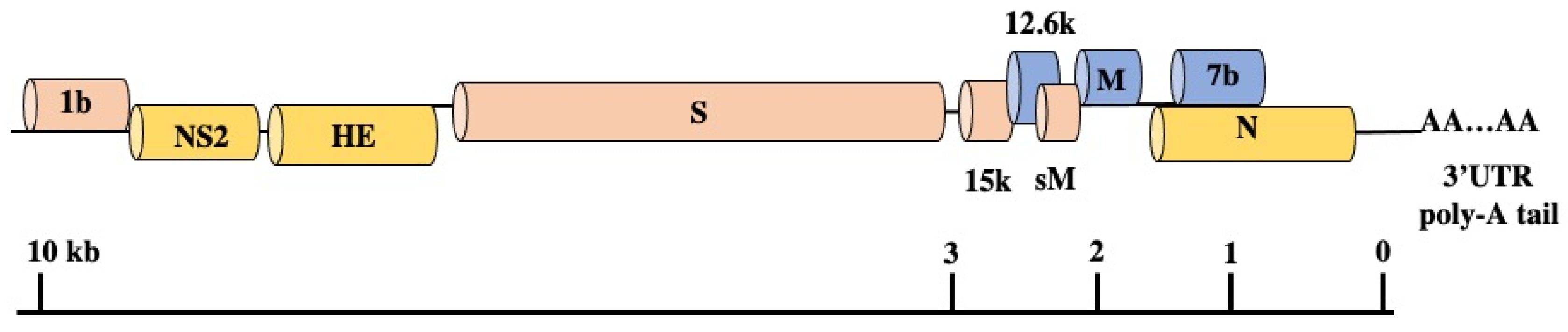
3.5. Genomic Structure and Gene Functions
3.6. SDAV Propagation and Role of In Vitro Studies
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mallapaty, S. The search for animals harbouring coronavirus-and why it matters. Nature 2021, 591, 26–28. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [Green Version]
- Schlottau, K.; Rissmann, M.; Graaf, A.; Schön, J.; Sehl, J.; Wylezich, C.; Höper, D.; Mettenleiter, T.C.; Balkema-Buschmann, A.; Harder, T.; et al. SARS-CoV-2 in fruit bats, ferrets, pigs, and chickens: An experimental transmission study. Lancet Microbe 2020, 1, 218–225. [Google Scholar] [CrossRef]
- Seyran, M.; Takayama, K.; Uversky, V.N.; Lundstrom, K.; Palù, G.; Sherchan, S.P.; Attrish, D.; Rezaei, N.; Aljabali, A.; Ghosh, S.; et al. The structural basis of accelerated host cell entry by SARS-CoV-2†. FEBS J. 2020, 288, 5010–5020. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Moneim, A.S.; Abdelwhab, E.M. Evidence for SARS-CoV-2 Infection of Animal Hosts. Pathogens 2020, 9, 529. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Jiao, H.; Liu, Q.; Zhang, Z.; Xiong, Q.; Wang, B.J.; Wang, X.; Guo, M.; Wang, L.F.; Lan, K.; et al. ACE2 receptor usage reveals variation in susceptibility to SARS-CoV and SARS-CoV-2 infection among bat species. Nat. Ecol. Evol. 2021, 5, 600–608. [Google Scholar] [CrossRef]
- Meerburg, G.B.; Singleton, R.G.; Kijlstra, A. Rodent-borne diseases and their risks for public health. Crit. Rev. Micro. 2009, 35, 221–270. [Google Scholar] [CrossRef]
- Alvarez-Munoz, S.; Upegui-Porras, N.; Gomez, A.P.; Ramirez-Nieto, G. Key Factors That Enable the Pandemic Potential of RNA Viruses and Inter-Species Transmission: A Systematic Review. Viruses 2021, 13, 537. [Google Scholar] [CrossRef]
- Han, B.A.; Schmidt, J.P.; Bowden, S.E.; Drake, J.M. Rodent reservoirs of future zoonotic diseases. Proc. Natl. Acad. Sci. USA 2015, 112, 7039–7044. [Google Scholar] [CrossRef] [Green Version]
- Jonas, A.M.; Craft, J.; Black, L.; Bhatt, P.N.; Hilding, D. Sialodacryoadenitis in the rat. A light and electron microscopic study. Arch. Pathol. 1969, 88, 613–622. [Google Scholar]
- Jacoby, R.O.; Bhatt, P.N.; Jonas, A.M. Pathogenesis of sialodacryoadenitis in gnotobiotic rats. Vet. Pathol. 1975, 12, 196–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Percy, D.H.; Hanna, P.E.; Paturzo, F.; Bhatt, P.N. Comparison of strain susceptibility to experimental sialodacryoadenitis in rats. Lab. Anim. Sci. 1984, 34, 255–260. [Google Scholar] [PubMed]
- Wojcinski, Z.W.; Percy, D.H. Sialodacryoadenitis virus-associated lesions in the lower respiratory tract of rats. Vet. Pathol. 1986, 23, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Hartley, J.W.; Rowe, W.P.; Bloom, J.J.; Turner, H.C. Antibodies to mouse hepatitis virus in human sera. Proc. Soc. Exp. Biol. Med. 1964, 115, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Coronaviruses; Nature: London, UK, 1968; pp. 220–650. [Google Scholar]
- Parker, J.C.; Cross, S.S.; Rowe, W.P. Rat coronavirus (RCV): A prevalent, naturally occurring pneumotropic virus of rats. Arch. Gesamte Virusforsch. 1970, 31, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, P.N.; Percy, D.H.; Jonas, A.M. Characterization of the virus of sialodacryoadenitis of rats: A member of the coronvarivus group. J. Infect. Dis. 1972, 126, 123–130. [Google Scholar] [CrossRef]
- Kojima, A.; Fujimani, F.; Doi, K.; Yasoshima, A.; Okaniwa, A. Isolation and properties of sialodacryoadenitis virus of rats. Exp. Anim. 1980, 29, 409–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maru, M.; Sato, K. Characterization of a coronavirus isolated from rats with sialodacryoadenitis. Arch. Virol. 1982, 73, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Compton, S.R.; Smith, A.L.; Gaertner, D.J. Comparison of the pathogenicity in rats of rat coronaviruses of different neutralization groups. Lab. Anim. Sci. 1999, 49, 514–518. [Google Scholar]
- Compton, S.R.; Vivas-Gonzales, B.E.; Macy, J.D. Reverse transcriptase chain reaction-based diagnosis and molecular characterization of a new rat coronavirus strain. Lab. Anim. Sci. 1999, 49, 506–513. [Google Scholar]
- Gaertner, D.J.; Compton, S.R.; Winograd, D.F. Environmental stability of rat coronaviruses (RCVs). Lab. Anim. Sci. 1993, 43, 403–404. [Google Scholar]
- La Regina, M.; Woods, L.; Klender, P.; Gaertner, D.J.; Paturzo, F.X. Transmission of sialodacryoadenitis virus (SDAV) from infected rats to rats and mice through handling, close contact, and soiled bedding. Lab. Anim. Sci. 1992, 42, 344–346. [Google Scholar]
- Bhatt, P.N.; Jacoby, R.O. Epizootiological observations of natural and experimental infection with sialodacryoadenitis virus in rats. Lab. Anim. Sci. 1985, 35, 129–134. [Google Scholar]
- Percy, D.H.; Bond, S.J.; Paturzo, F.X.; Bhatt, P.N. Duration of protection from reinfection following exposure to sialodacryoadenitis virus in Wistar rats. Lab. Anim. Sci. 1990, 40, 144–149. [Google Scholar] [PubMed]
- Weir, E.C.; Jacoby, R.O.; Paturzo, E.X.; Johnson, E.A. Infection of SDAV-immune rats with SDAV and rat coronavirus. Lab. Anim. Sci. 1990, 40, 363–366. [Google Scholar] [PubMed]
- Percy, D.H.; Williams, K.L.; Croy, B.A. Experimental sialodacryoadenitis virus infection in severe combined immunodeficient mice. Can. J. Vet. Res. 1991, 55, 89–90. [Google Scholar] [PubMed]
- Percy, D.H.; Scott, R.A. Coronavirus infection in the laboratory rat: Immunization trials using attenuated virus replicated in L-2 cells. Can. J. Vet. Res. 1991, 55, 60–66. [Google Scholar] [PubMed]
- Sato, T.; Meguid, M.M.; Quinn, R.H.; Zhang, L.; Chen, C. Feeding behavior during sialodacryoadenitis viral infection in rats. Physiol. Behav. 2001, 72, 721–726. [Google Scholar] [CrossRef]
- Barthold, S.W.; Griffey, S.M.; Percy, D.H. Rat, Pathology of Laboratory Rodents and Rabbits, 4th ed.; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2016; pp. 119–172. [Google Scholar]
- Smith, A.L.; Winograd, D.F. Two enzyme immunoassays for the detection of antibody to rodent coronaviruses. J. Virol. Methods 1986, 14, 335–343. [Google Scholar] [CrossRef]
- Homberger, F.R.; Smith, A.L.; Barthold, S.W. Detection of rodent coronaviruses in tissues and cell cultures by using polymerase chain reaction. J. Clin. Microbiol. 1991, 29, 2789–2793. [Google Scholar] [CrossRef] [Green Version]
- Besselsen, D.G.; Wagner, A.M.; Loganbill, J.K. Detection of rodent coronaviruses by use of fluorogenic reverse transcriptase-polymerase chain reaction analysis. Comp. Med. 2002, 52, 111–116. [Google Scholar]
- Gorbalenya, A.E.; Enjuanes, L.; Ziebuhr, J.; Snijder, E.J. Nidovirales: Evolving the largest RNA virus genome. Virol. Res. 2006, 117, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, K.; Mulley, G.; Gulyaeva, A.A.; Zhao, L.Y.; Shu, G.C.; Jiang, J.P.; Neuman, B.W. Description and initial characterization of metatranscriptomic nidovirus-like genomes from the proposed new family Abyssoviridae, and from a sister group to the Coronavirinae, the proposed genus Alphaletovirus. Virol. J. 2018, 524, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; et al. Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses (2020). Arch. Virol. 2020, 165, 2737–2748. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Qiu, Y.; Ge, X. The taxonomy, host range and pathogenicity of coronaviruses and other viruses in the Nidovirales order. Anim. Dis. 2021, 1, 1–28. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Revs. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Lu, L.; Du, J.; Yang, L.; Ren, X.; Liu, B.; Jiang, J.; Yang, J.; Dong, J.; Sun, L.; et al. Comparative analysis of rodent and small mammal viromes to better understand the wildlife origin of emerging infectious diseases. Microbiome 2018, 6, 178. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Woo, P.C.Y.; Li, K.S.M.; Tsang, A.K.L.; Fan, R.Y.Y.; Luk, H.K.H.; Cai, J.-P.; Chan, K.-H.; Zheng, B.-J.; Wang, M.; et al. Discovery of a novel coronavirus, China Rattus coronavirus HKU24, from Norway rats supports the murine origin of Betacoronavirus 1 and has implications for the ancestor of Betacoronavirus lineage A. J. Virol. 2015, 89, 3076–3092. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Grunewald, M.; Perlman, S. Coronaviruses: An Updated Overview of Their Replication and Pathogenesis. Methods Mol. Biol. 2020, 2203, 1–29. [Google Scholar] [CrossRef]
- Barker, M.G.; Percy, D.H.; Hovland, D.J.; MacInnes, J.I. Preliminary characterization of the structural proteins of the coronaviruses, sialodacryoadenitis virus and Parker’s rat coronavirus. Can. J. Vet. Res. 1994, 58, 99–103. [Google Scholar]
- Yoo, D.; Pei, Y.; Christie, N.; Cooper, M. Primary structure of the sialodacryoadenitis virus genome: Sequence of the structural-protein region and its application for differential diagnosis. Clin. Diag. Lab. Immunol. 2000, 7, 568–573. [Google Scholar] [CrossRef] [Green Version]
- Strasser, P.; Unger, U.; Strobl, B.; Vilas, U.; Vlasak, R. Recombinant viral sialate-O-acetylesterases. Glyc. J. 2004, 20, 551–561. [Google Scholar] [CrossRef]
- Compton, S.R.; Kunita, S. Characterization of the S protein of enterotropic murine coronavirus strain-Y. Adv. Exp. Med. Biol. 1995, 380, 23–28. [Google Scholar] [CrossRef]
- Raamsman, M.J.; Locker, J.K.; de Hooge, A.; de Vries, A.A.; Griffiths, G.; Vennema, H.; Rottier, P.J. Characterization of the coronavirus mouse hepatitis virus strain A59 small membrane protein E. J. Virol. 2000, 74, 2333–2342. [Google Scholar] [CrossRef] [Green Version]
- Yadav, R.; Chaudhary, J.K.; Jain, N.; Chaudhary, P.K.; Khanra, S.; Dhamija, P.; Sharma, A.; Kumar, A.; Handu, S. Role of Structural and Non-Structural Proteins and Therapeutic Targets of SARS-CoV-2 for COVID-19. Cells 2021, 10, 821. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.S.; Liu, X.D. Similarities and Dissimilarities of COVID-19 and Other Coronavirus Diseases. Annu. Rev. Microbiol. 2021, 75, 1–29. [Google Scholar] [CrossRef] [PubMed]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Hogue, B.G.; Machamer, C.E. Coronavirus structural proteins and virus assembly. In Nidoviruses; Perlman, S., Ed.; ASM Press: Washington, DC, USA, 2007; pp. 179–200. [Google Scholar]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.M.; Baksh, F.M.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G.; et al. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, K.; Chen, C.J.; Maeda, J.; Makino, S. Nucleocapsid-independent specific viral RNA packaging via viral envelope protein and viral RNA signal. J. Virol. 2003, 77, 2922–2927. [Google Scholar] [CrossRef] [Green Version]
- Masters, P.S. The molecular biology of coronaviruses. Adv. Virus Res. 2006, 66, 193–292. [Google Scholar]
- Wilson, L.; McKinlay, C.; Gage, P.; Ewart, G. SARS coronavirus E protein forms cation- selective ion channels. Virology 2004, 330, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Kunita, S.; Mori, M.; Terada, E. Sequence analysis of the nucleocapsid protein gene of rat coronavirus SDAV-681. Virology 1993, 193, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Payne, S. In Viruses: From Understanding to Investigation, 1st ed.; Payne, S., Ed.; Academic Press: New York, NY, USA, 2017; Volume 1, pp. 149–158. [Google Scholar] [CrossRef]
- Cowley, T.J.; Long, S.Y.; Weiss, S.R. The murine coronavirus nucleocapsid gene is a determinant of virulence. J. Virol. 2010, 84, 1752–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, Y.; Li, W.; Li, Z.; Koerhuis, D.; van den Burg, A.; Rozemuller, E.; Bosch, B.J.; van Kuppeveld, F.; Boons, G.J.; Huizinga, E.G.; et al. Coronavirus hemagglutinin-esterase and spike proteins coevolve for functional balance and optimal virion avidity. Proc. Nat. Acad. Sci. USA 2020, 117, 25759–25770. [Google Scholar] [CrossRef] [PubMed]
- Klausegger, A.; Strobl, B.; Regl, G.; Kaser, A.; Luytjes, W.; Vlasak, R. Identification of a coronavirus hemagglutinin-esterase with a substrate specificity different from those of influenza C virus and bovine coronavirus. J. Virol. 1999, 73, 3737–3743. [Google Scholar] [CrossRef] [Green Version]
- Kienzle, T.E.; Abraham, S.; Hogue, B.G.; Brian, D.A. Structure and orientation of expressed bovine coronavirus hemagglutinin-esterase protein. J. Virol. 1990, 64, 1834–1838. [Google Scholar] [CrossRef] [Green Version]
- Vlasak, R.; Luytjes, W.; Spaan, W.; Palese, P. Human and bovine coronaviruses recognize sialic acid-containing receptors similar to those of influenza C viruses. Proc. Natl. Acad. Sci. USA 1988, 85, 4526–4529. [Google Scholar] [CrossRef] [Green Version]
- Vlasak, R.; Luytjes, W.; Leider, J.; Spaan, W.; Palese, P. The E3 protein of bovine coronavirus is a receptor-destroying enzyme with acetylesterase activity. J. Virol. 1988, 62, 4686–4690. [Google Scholar] [CrossRef] [Green Version]
- Yokomori, K.; Banner, L.R.; Lai, M.M. Heterogeneity of gene expression of the hemagglutinin-esterase (HE) protein of murine coronaviruses. Virology 1991, 183, 647–657. [Google Scholar] [CrossRef]
- Yokomori, K.; Baker, S.C.; Stohlman, S.A.; Lai, M.M. Hemagglutinin-esterase-specific monoclonal antibodies alter the neuropathogenicity of mouse hepatitis virus. J. Virol. 1992, 66, 2865–2874. [Google Scholar] [CrossRef] [Green Version]
- Yokomori, K.; Asanaka, M.; Stohlman, S.A.; Makino, S.; Shubin, R.A.; Gilmore, W.; Weiner, L.P.; Wang, F.I.; Lai, M.M. Neuropathogenicity of mouse hepatitis virus JHM isolates differing in hemagglutinin-esterase protein expression. J. Neurovirol. 1995, 1, 330–339. [Google Scholar] [CrossRef]
- Weiss, S.R.; Leibowitz, J.L. Coronavirus pathogenesis. Adv. Virus Res. 2011, 81, 85–164. [Google Scholar] [CrossRef]
- Percy, D.H.; Lynch, J.A.; Descôteaux, J.P. Central nervous system lesions in suckling mice and rats inoculated intranasally with sialodacryoadenitis virus. Vet. Pathol. 1986, 23, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Funk, C.J.; Manzer, R.; Miura, T.A.; Groshong, S.D.; Ito, Y.; Travanty, E.A.; Leete, J.; Holmes, K.V.; Mason, R.J. Rat respiratory coronavirus infection: Replication in airway and alveolar epithelial cells and the innate immune response. J. Gen. Virol. 2009, 90, 2956–2964. [Google Scholar] [CrossRef]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagneten, S.; Scanga, C.A.; Dveksler, G.S.; Beauchemin, N.; Percy, D.; Holmes, K.V. Attachment glycoproteins and receptor specificity of rat coronaviruses. Lab. Anim. Sci. 1996, 46, 159–166. [Google Scholar]
- King, B.; Potts, B.J.; Brian, D.A. Bovine coronavirus hemagglutinin protein. Virus Res. 1985, 2, 53–59. [Google Scholar] [CrossRef]
- Williams, R.K.; Jiang, G.S.; Holmes, K.V. Receptor for mouse hepatitis virus is a member of the carcinoembryonic antigen family of glycoproteins. Proc. Nat. Acad. Sci. USA 1991, 88, 5533–5536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regl, G.; Kaser, A.; Iwersen, M.; Schmid, H.; Kohla, G.; Strobl, B.; Vilas, U.; Schauer, R.; Vlasak, R. The hemagglutinin-esterase of mouse hepatitis virus strain S is a sialate-4-O-acetylesterase. J. Virol. 1999, 73, 4721–4727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurzer, W.J.; Obojes, K.; Vlasak, R. The sialate-4-O-acetylesterases of coronaviruses related to mouse hepatitis virus: A proposal to reorganize group 2 Coronaviridae. J. Gen. Virol. 2002, 83, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Smits, S.L.; Gerwig, G.J.; van Vliet, A.L.; Lissenberg, A.; Briza, P.; Kamerling, J.P.; Vlask, R.; de Groot, R.J. Nidovirus sialate-O-acetylesterases: Evolution and substrate specificity of corona- and toroviral receptor-destroying enzymes. J. Biol. Chem. 2005, 280, 6933–6941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langereis, M.A.; van Vliet, A.L.; Boot, W.; de Groot, R.J. Attachment of mouse hepatitis virus to O-acetylated sialic acid is mediated by hemagglutinin-esterase and not by the spike protein. J. Virol. 2010, 84, 8970–8974. [Google Scholar] [CrossRef] [Green Version]
- Langereis, M.A.; Zeng, Q.; Heesters, B.A.; Huizinga, E.G.; de Groot, R.J. The murine coronavirus hemagglutinin-esterase receptor-binding site: A major shift in ligand specificity through modest changes in architecture. PLoS Pathog. 2012, 8, 1002492. [Google Scholar] [CrossRef]
- Bakkers, M.J.; Zeng, Q.; Feitsma, L.J.; Hulswit, R.J.; Li, Z.; Westerbeke, A.; van Kuppeveld, F.J.; Boons, G.J.; Langereis, M.A.; Huizinga, E.G.; et al. Coronavirus receptor switch explained from the stereochemistry of protein-carbohydrate interactions and a single mutation. Proc. Natl. Acad. Sci. USA 2016, 113, 3111–3119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulswit, R.J.G.; Lang, Y.; Bakkers, M.J.G.; Li, W.; Li, Z.; Schouten, A.; Ophorst, B.; van Kuppeveld, F.J.M.; Boons, G.J.; Bosch, B.J.; et al. Human coronaviruses OC43 and HKU1 bind to 9-O-acetylated sialic acids via a conserved receptor-binding site in spike protein domain A. Proc. Natl. Acad. Sci. USA 2019, 116, 2681–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lissenberg, A.; Vrolijk, M.M.; van Vliet, A.L.; Langereis, M.A.; de Groot-Mijnes, J.D.; Rottier, P.J.; de Groot, R.J. Luxury at a cost? Recombinant mouse hepatitis viruses expressing the accessory hemagglutinin esterase protein display reduced fitness in vitro. J. Virol. 2005, 79, 15054–15063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olofsson, S.; Bergström, T. Glycoconjugate glycans as viral receptors. Ann. Med. 2005, 37, 154–172. [Google Scholar] [CrossRef] [PubMed]
- Hingley, S.T.; Leparc-Goffart, I.; Weiss, S.R. The spike protein of murine coronavirus mouse hepatitis virus strain A59 is not cleaved in primary glial cells and primary hepatocytes. J. Virol. 1998, 72, 1606–1609. [Google Scholar] [CrossRef] [Green Version]
- White, J.M.; Whittaker, G.R. Fusion of Enveloped Viruses in Endosomes. Traffic 2016, 7, 593–614. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, S.; Taguchi, F. Receptor-induced conformational changes of murine coronavirus spike protein. J. Virol. 2002, 76, 11819–11826. [Google Scholar] [CrossRef] [Green Version]
- Nash, T.C.; Buchmeier, M.J. Entry of mouse hepatitis virus into cells by endosomal and nonendosomal pathways. Virology 1997, 233, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkard, C.; Verheije, M.H.; Wicht, O.; van Kasteren, S.I.; van Kuppeveld, F.J.; Haagmans, B.L.; Pelkmans, L.; Rottier, P.J.; Bosch, B.J.; de Haan, C.A. Coronavirus cell entry occurs through the endo-/lysosomal pathway in a proteolysis-dependent manner. PLoS Pathog. 2014, 10, 1004502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephensen, C.B.; Casebolt, D.B.; Gangopadhyay, N.N. Phylogenetic analysis of a highly conserved region of the polymerase gene from 11 coronaviruses and development of a consensus polymerase chain reaction assay. Virus Res. 1999, 60, 181–189. [Google Scholar] [CrossRef]
- Denison, M.R.; Perlman, S. Translation and processing of mouse hepatitis virus virion RNA in a cell-free system. J. Virol. 1986, 60, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Kanjanahaluethai, A.; Chen, Z.; Jukneliene, D.; Baker, S.C. Membrane topology of murine coronavirus replicase nonstructural protein 3. Virology 2007, 361, 391–401. [Google Scholar] [CrossRef] [Green Version]
- Oostra, M.; Hagemeijer, M.C.; van Gent, M.; Bekker, C.P.; te Lintelo, E.G.; Rottier, P.J.; de Haan, C.A. Topology and membrane anchoring of the coronavirus replication complex: Not all hydrophobic domains of nsp3 and nsp6 are membrane spanning. J. Virol. 2008, 82, 12392–12405. [Google Scholar] [CrossRef] [Green Version]
- Neuman, B.W.; Chamberlain, P.; Bowden, F.; Joseph, J. Atlas of coronavirus replicase structure. Virus Res. 2014, 194, 49–66. [Google Scholar] [CrossRef]
- Sawicki, S.G.; Sawicki, D.L. Coronaviruses use discontinuous extension for synthesis of subgenome-length negative strands. Adv. Exp. Med. Biol. 1995, 380, 499–506. [Google Scholar]
- Sola, I.; Almazán, F.; Zúñiga, S.; Enjuanes, L. Continuous and discontinuous RNA synthesis in coronaviruses. Annu. Rev. Virol. 2015, 2, 265–288. [Google Scholar] [CrossRef] [Green Version]
- Di, H.; McIntyre, A.A.; Brinton, M.A. New insights about the regulation of Nidovirus subgenomic mRNA synthesis. Virology 2008, 517, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Viehweger, A.; Krautwurst, S.; Lamkiewicz, K.; Madhugiri, R.; Ziebuhr, J.; Hölzer, M.; Marz, M. Direct RNA nanopore sequencing of full-length coronavirus genomes provides novel insights into structural variants and enables modification analysis. Gen. Res. 2019, 29, 1545–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stertz, S.; Reichelt, M.; Spiegel, M.; Kuri, T.; Martínez-Sobrido, L.; García-Sastre, A.; Weber, F.; Kochs, G. The intracellular sites of early replication and budding of SARS-coronavirus. Virology 2007, 361, 304–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Haan, C.A.; Rottier, P.J. Molecular interactions in the assembly of coronaviruses. Adv. Vir. Res. 2005, 64, 165–230. [Google Scholar] [CrossRef]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Meth. Mol. Biol. 2015, 1282, 1–23. [Google Scholar] [CrossRef]
- Visser, E.A.; Moons, S.J.; Timmermans, S.; de Jong, H.; Boltje, T.J.; Büll, C. Sialic acid O-acetylation: From biosynthesis to roles in health and disease. J. Biol. Chem. 2021, 297, 100906. [Google Scholar] [CrossRef]
- Miura, T.A.; Wang, J.; Holmes, K.V.; Mason, R.J. Rat coronaviruses infect rat alveolar type I epithelial cells and induce expression of CXC chemokines. Virology 2007, 369, 288–298. [Google Scholar] [CrossRef] [Green Version]
- Gaertner, D.J.; Compton, S.R.; Winograd, D.F.; Smith, A.L. Growth characteristics and protein profiles of prototype and wild-type rat coronavirus isolates grown in a cloned subline of mouse fibroblasts (L2p.176 cells). Virus Res. 1996, 41, 55–68. [Google Scholar] [CrossRef]
- Percy, D.; Bond, S.; MacInnes, J. Replication of sialodacryoadenitis virus in mouse L-2 cells. Arch. Virol. 1989, 104, 323–333. [Google Scholar] [CrossRef]
- Hirano, N.; Suzuki, Y.; Ono, K.; Murakami, T.; Fujiwara, K. Growth of rat sialodacryoadenitis viruses in LBC cell culture. Jap. J. Vet. Sci. 1986, 48, 193–195. [Google Scholar] [CrossRef] [Green Version]
- Ohsawa, K.; Watanabe, Y.; Takakura, A.; Itoh, T.; Sato, H. Replication of rat coronaviruses in intestinal cell line, RCN-9, derived from F344 rats. J. Exp. Anim. 1996, 45, 389–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macy, J.D.; Compton, S.R. Viral diseases. In Laboratory Rat, 3rd ed.; Suckow, M.A., Hankenson, F.C., Wilson, R.P., Foley, P.L., Eds.; Academic Press: Washington, DC, USA, 2020; Chapter 131; pp. 541–568. ISBN 9780128143384. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Huang, Y.; Yuen, K.-Y. Coronavirus Diversity, Phylogeny and Interspecies Jumping. Exp. Biol. Med. 2009, 234, 1117–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhama, K.; Patel, S.K.; Sharun, K.; Pathak, M.; Tiwari, R.; Yatoo, M.I.; Malik, Y.S.; Sah, R.; Rabaan, A.A.; Panwar, P.K.; et al. SARS-CoV-2 jumping the species barrier: Zoonotic lessons from SARS, MERS and recent advances to combat this pandemic virus. Travel Med. Infect. Dis. 2020, 37, 101830. [Google Scholar] [CrossRef] [PubMed]
- de Bruin, W.C.; van de Ven, E.M.; Hooijmans, C.R. Efficacy of Soiled Bedding Transfer for Transmission of Mouse and Rat Infections to Sentinels: A Systematic Review. PLoS ONE 2016, 11, e0158410. [Google Scholar] [CrossRef]
- Mishra, R.; Banerjea, A.C. Neurological Damage by Coronaviruses: A Catastrophe in the Queue! Front. Immunol. 2020, 11, 565521. [Google Scholar] [CrossRef] [PubMed]
- Palmer, S.; Brown, D.; Morgan, D. Early qualitative risk assessment of the emerging zoonotic potential of animal diseases. BMJ 2005, 331, 1256–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Genus | Species | Substrate * |
|---|---|---|---|
| Coronaviridae | Coronavirus | Human coronavirus OC43 | Neu5,9Ac2 |
| Bovine coronavirus | |||
| Hemagglutinating encephalomyelitis virus | |||
| Mouse hepatitis virus | Neu4,5Ac2 | ||
| Puffinosis coronavirus | Neu4,5Ac2 | ||
| Sialodacryoadenitis virus | Neu4,5Ac2 | ||
| Torovirus | Porcine torovirus | n.d. | |
| Bovine torovirus | |||
| Orthomyxoviridae | Isavirus | Infectious salmon anemia virus | Neu4,5Ac2 |
| Influenza C virus | Influenza C virus | Neu5,9Ac2 (α2-8-linked, GL) | |
| Influenza D virus | Influenza D virus | Neu5,9Ac2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartak, M.; Słońska, A.; Bańbura, M.W.; Cymerys, J. SDAV, the Rat Coronavirus—How Much Do We Know about It in the Light of Potential Zoonoses. Viruses 2021, 13, 1995. https://doi.org/10.3390/v13101995
Bartak M, Słońska A, Bańbura MW, Cymerys J. SDAV, the Rat Coronavirus—How Much Do We Know about It in the Light of Potential Zoonoses. Viruses. 2021; 13(10):1995. https://doi.org/10.3390/v13101995
Chicago/Turabian StyleBartak, Michalina, Anna Słońska, Marcin W Bańbura, and Joanna Cymerys. 2021. "SDAV, the Rat Coronavirus—How Much Do We Know about It in the Light of Potential Zoonoses" Viruses 13, no. 10: 1995. https://doi.org/10.3390/v13101995
APA StyleBartak, M., Słońska, A., Bańbura, M. W., & Cymerys, J. (2021). SDAV, the Rat Coronavirus—How Much Do We Know about It in the Light of Potential Zoonoses. Viruses, 13(10), 1995. https://doi.org/10.3390/v13101995