
Bovine Leukaemia Virus: Current Epidemiological Circumstance and Future Prospective

 ,
,  ,
,  ,
,  ,
, 

Abstract
:1. Introduction
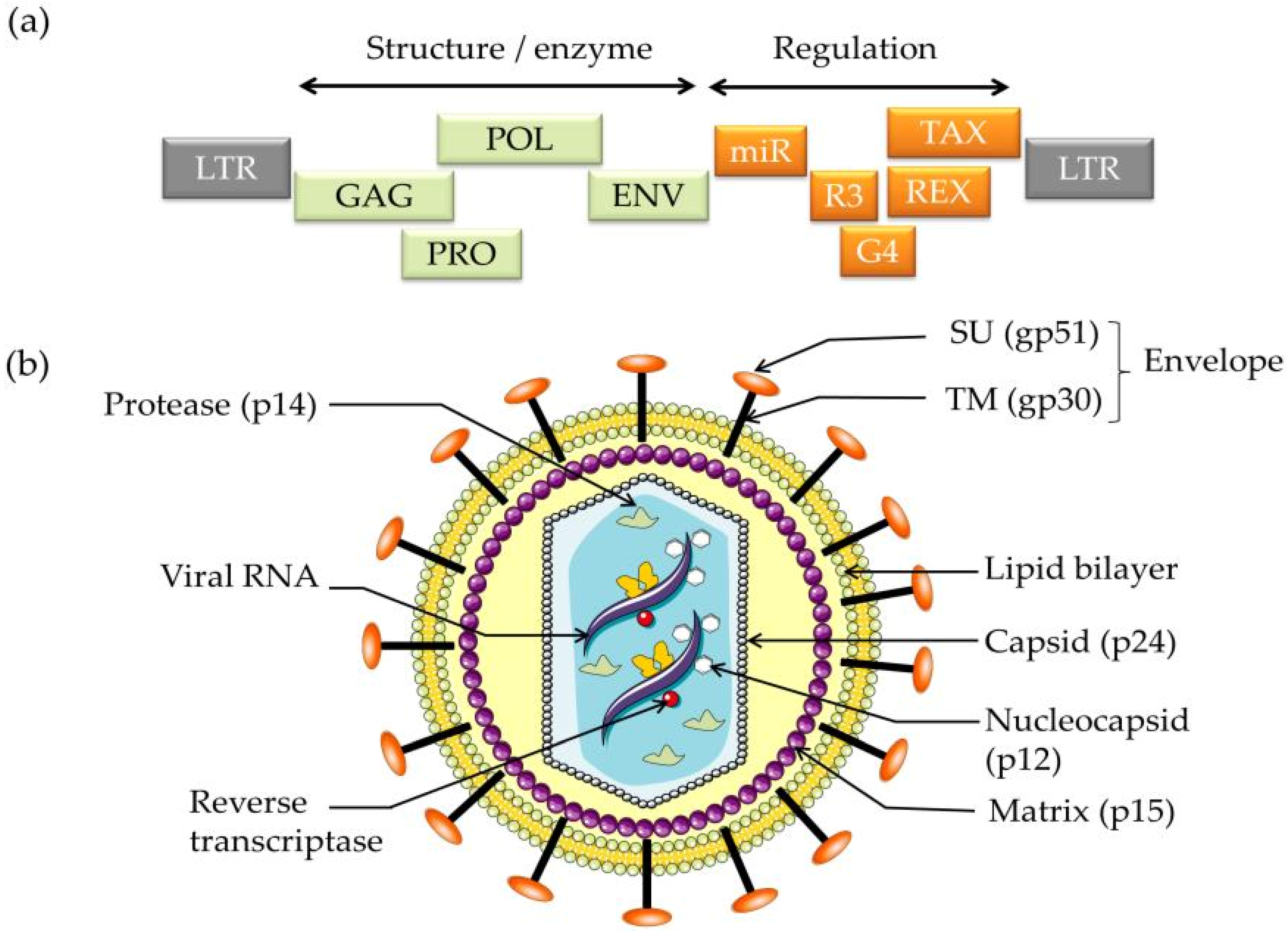
2. BLV Genome
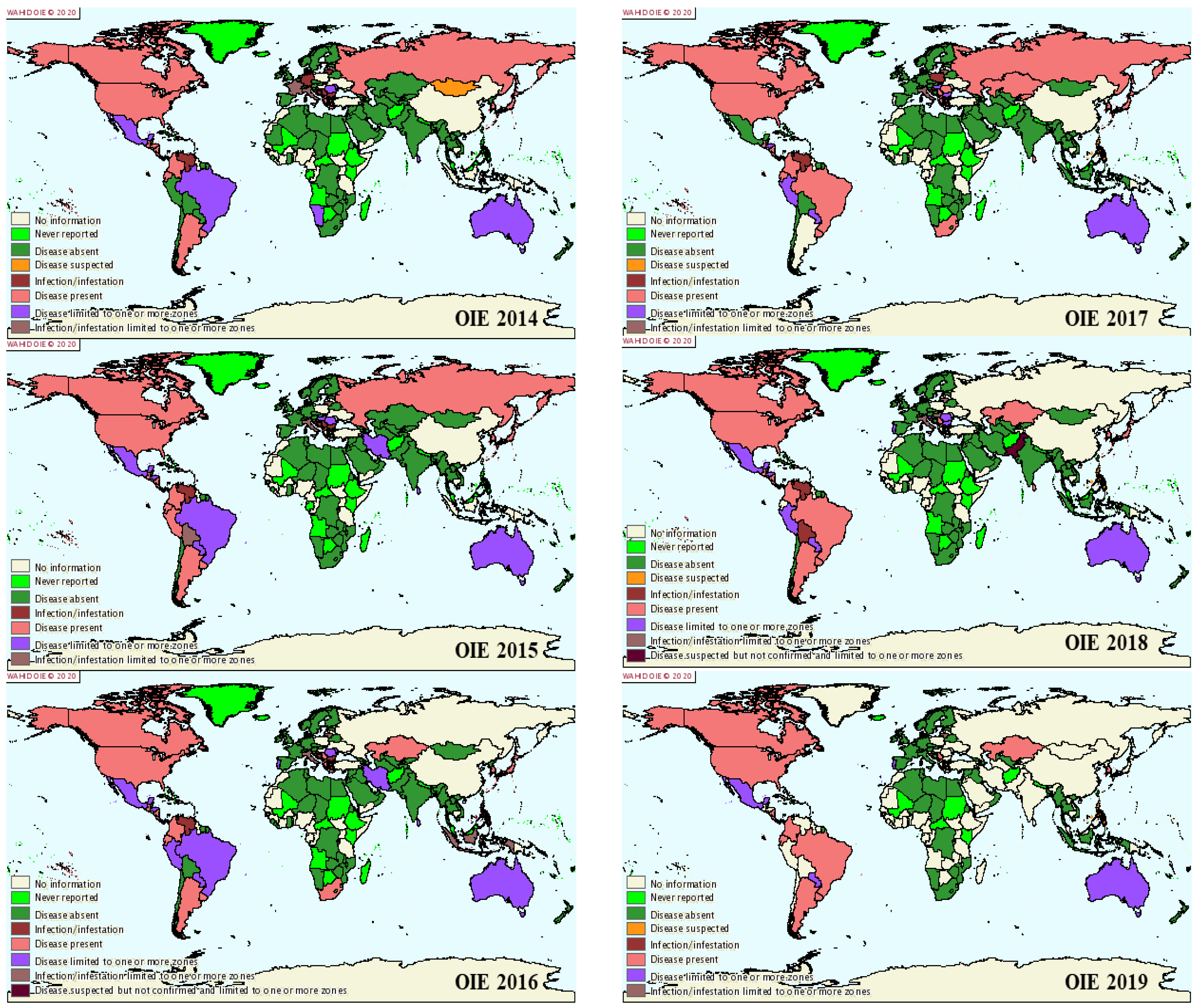
3. BLV Prevalence
4. BLV Susceptibility
5. Transmission
6. BLV Pathogenesis
7. Clinical Outcomes
8. Zoonotic Potential of BLV
9. Diagnosis
10. Strategies for the Control of BLV Infection
11. Conclusions
12. Future Prospective Studies
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aida, Y.; Murakami, H.; Takagashi, M.; Takeshima, S.-N. Mechanisms of pathogenesis induced by bovine leukemia virus as a model for human T-cell leukemia virus. Front. Microbiol. 2013, 4, 328. [Google Scholar] [CrossRef] [Green Version]
- Brujeni, G.N.; Ghorbanpour, R.; Esmailnejad, A. Association of BoLA-DRB3. 2 alleles with BLV infection profiles (persistent lymphocytosis/lymphosarcoma) and lymphocyte subsets in Iranian Holstein cattle. Biochem. Genet. 2016, 54, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Maclachlan, N.J.; Dubovi, E.J. Fenner’s Veterinary Virology; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Martin, F.; Bangham, C.R.; Ciminale, V.; Lairmore, M.D.; Murphy, E.L.; Switzer, W.M.; Mahieux, R. Conference highlights of the 15th international conference on human retrovirology: HTLV and related retroviruses, 4–8 june 2011, Leuven, Gembloux, Belgium. Retrovirology 2011, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nekouei, O.; Stryhn, H.; VanLeeuwen, J.; Kelton, D.; Hanna, P.; Keefe, G. Predicting within-herd prevalence of infection with bovine leukemia virus using bulk-tank milk antibody levels. Prev. Vet. Med. 2015, 122, 53–60. [Google Scholar] [CrossRef]
- Moratorio, G.; Fischer, S.; Bianchi, S.; Tomé, L.; Rama, G.; Obal, G.; Carrión, F.; Pritsch, O.; Cristina, J. A detailed molecular analysis of complete Bovine Leukemia Virus genomes isolated from B-cell lymphosarcomas. Vet. Res. 2013, 44, 19. [Google Scholar] [CrossRef] [Green Version]
- Juliarena, M.A.; Barrios, C.N.; Lützelschwab, C.M.; Esteban, E.N.; Gutiérrez, S.E. Bovine Leukemia Virus: Current Perspectives; Dove Press: Manchester, UK, 2017. [Google Scholar]
- Pandey, G.S.; Simulundu, E.; Mwiinga, D.; Samui, K.L.; Mweene, A.S.; Kajihara, M.; Mangani, A.; Mwenda, R.; Ndebe, J.; Konnai, S. Clinical and subclinical bovine leukemia virus infection in a dairy cattle herd in Zambia. Arch. Virol. 2017, 162, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Aida, Y.; Miyasaka, M.; Okada, K.; Onuma, M.; Kogure, S.; Suzuki, M.; Minoprio, P.; Levy, D.; Ikawa, Y. Further phenotypic characterization of target cells for bovine leukemia virus experimental infection in sheep. Am. J. Vet. Res. 1989, 50, 1946–1951. [Google Scholar] [PubMed]
- Frie, M.C.; Coussens, P.M. Bovine leukemia virus: A major silent threat to proper immune responses in cattle. Vet. Immunol. Immunopathol. 2015, 163, 103–114. [Google Scholar] [CrossRef]
- Nishimori, A.; Konnai, S.; Ikebuchi, R.; Okagawa, T.; Nakahara, A.; Murata, S.; Ohashi, K. Direct polymerase chain reaction from blood and tissue samples for rapid diagnosis of bovine leukemia virus infection. J. Vet. Med. Sci. 2016, 78, 791–796. [Google Scholar] [CrossRef] [Green Version]
- Panel, E.A. Scientific opinion on enzootic bovine leukosis. EFSA J. 2015, 13, 4188. [Google Scholar]
- Erskine, R.; Bartlett, P.; Byrem, T.; Render, C.; Febvay, C.; Houseman, J. Association between bovine leukemia virus, production, and population age in Michigan dairy herds. J. Dairy Sci. 2012, 95, 727–734. [Google Scholar] [CrossRef]
- Norby, B.; Bartlett, P.; Byrem, T.; Erskine, R. Effect of infection with bovine leukemia virus on milk production in Michigan dairy cows. J. Dairy Sci. 2016, 99, 2043–2052. [Google Scholar] [CrossRef] [PubMed]
- Manaa, E.; Marawan, M.; Abdelhady, A.; Selim, A. Association between bovine leukemia virus infection, reproductive performance and milk production in water buffaloes and dairy cattle in Egypt. Adv. Anim. Vet. Sci. 2020, 8, 1109–1113. [Google Scholar] [CrossRef]
- OIE. Enzootic bovine leukosis. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health: Paris, France, 2018; pp. 1113–1124. Available online: https://www.oie.int/standard-setting/terrestrial-manual/access-online/ (accessed on 11 August 2020).
- LaDronka, R.M.; Ainsworth, S.; Wilkins, M.J.; Norby, B.; Byrem, T.M.; Bartlett, P.C. Prevalence of bovine leukemia virus antibodies in US dairy cattle. Vet. Med. Int. 2018, 2018, 5831278. [Google Scholar] [CrossRef] [Green Version]
- Ruggiero, V.; Norby, B.; Benitez, O.; Hutchinson, H.; Sporer, K.; Droscha, C.; Swenson, C.; Bartlett, P. Controlling bovine leukemia virus in dairy herds by identifying and removing cows with the highest proviral load and lymphocyte counts. J. Dairy Sci. 2019, 102, 9165–9175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, S.L.; Johnson, R.; Wells, S.J. Association between bovine-leukosis virus seroprevalence and herd-level productivity on US dairy farms. Prev. Vet. Med. 2003, 61, 249–262. [Google Scholar] [CrossRef]
- Olaya-Galan, N.; Corredor-Figueroa, A.; Guzman-Garzon, T.; Rios-Hernandez, K.; Salas-Cardenas, S.; Patarroyo, M.; Gutierrez, M. Bovine leukaemia virus DNA in fresh milk and raw beef for human consumption. Epidemiol. Infect. 2017, 145, 3125–3130. [Google Scholar] [CrossRef] [Green Version]
- Buehring, G.C.; Shen, H.M.; Jensen, H.M.; Choi, K.Y.; Sun, D.; Nuovo, G. Bovine leukemia virus DNA in human breast tissue. Emerg. Infect. Dis. 2014, 20, 772. [Google Scholar] [CrossRef] [Green Version]
- Buehring, G.C.; Shen, H.M.; Jensen, H.M.; Jin, D.L.; Hudes, M.; Block, G. Exposure to bovine leukemia virus is associated with breast cancer: A case-control study. PLoS ONE 2015, 10, e0134304. [Google Scholar] [CrossRef] [Green Version]
- Giovanna, M.; Carlos, U.J.; María, U.A.; Gutierrez, M.F. Bovine Leukemia Virus Gene Segment Detected in Human Breast Tissue. Open J. Med. J. 2013, 3, 28694. [Google Scholar] [CrossRef] [Green Version]
- Buehring, G.C.; Shen, H.; Schwartz, D.A.; Lawson, J.S. Bovine leukemia virus linked to breast cancer in Australian women and identified before breast cancer development. PLoS ONE 2017, 12, e0179367. [Google Scholar] [CrossRef] [Green Version]
- Olaya-Galán, N.N.; Salas-Cárdenas, S.P.; Rodriguez-Sarmiento, J.L.; Ibáñez-Pinilla, M.; Monroy, R.; Corredor-Figueroa, A.P.; Rubiano, W.; de la Peña, J.; Shen, H.; Buehring, G.C. Risk factor for breast cancer development under exposure to bovine leukemia virus in Colombian women: A case-control study. PLoS ONE 2021, 16, e0257492. [Google Scholar] [CrossRef] [PubMed]
- Zur Hausen, H.; de Villiers, E.M. Dairy cattle serum and milk factors contributing to the risk of colon and breast cancers. Int. J. Cancer 2015, 137, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Lawson, J.S.; Salmons, B.; Glenn, W.K. Oncogenic Viruses and Breast Cancer: Mouse Mammary Tumor Virus (MMTV), Bovine Leukemia Virus (BLV), Human Papilloma Virus (HPV), and Epstein-Barr Virus (EBV). Front. Oncol. 2018, 8, 1. [Google Scholar] [CrossRef]
- Boris-Lawrie, K.; Altanerova, V.; Altaner, C.; Kucerova, L.; Temin, H.M. In vivo study of genetically simplified bovine leukemia virus derivatives that lack tax and rex. J. Virol. 1997, 71, 1514–1520. [Google Scholar] [CrossRef] [Green Version]
- Pătraşcu, I.; Coman, S.; Sandu, I.; Stiube, P.; Munteanu, I.; Coman, T.; Ionescu, M.; Popescu, D.; Mihăilescu, D. Specific protection against bovine leukemia virus infection conferred on cattle by the Romanian inactivated vaccine BL-VACC-RO. Virologie 1980, 31, 95–102. [Google Scholar]
- Ristau, E.; Beier, D.; Wittmann, W.; Klima, F. Protection of sheep against infection with bovine leukemia virus by vaccination with tumor cells or tumor cell preparations from lymph nodes of leukemic cattle. Arch. Exp. Vet. 1987, 41, 185–196. [Google Scholar]
- Sagata, N.; Yasunaga, T.; Ohishi, K.; Tsuzuku-Kawamura, J.; Onuma, M.; Ikawa, Y. Comparison of the entire genomes of bovine leukemia virus and human T-cell leukemia virus and characterization of their unidentified open reading frames. EMBO J. 1984, 3, 3231–3237. [Google Scholar] [CrossRef]
- Zhao, X.; Buehring, G.C. Natural genetic variations in bovine leukemia virus envelope gene: Possible effects of selection and escape. Virology 2007, 366, 150–165. [Google Scholar] [CrossRef] [Green Version]
- Donnik, I. Productivity and health markers for large cattle. Int. J. Green Pharm. (IJGP) 2017, 11, S620–S625. [Google Scholar]
- Derse, D. Bovine leukemia virus transcription is controlled by a virus-encoded trans-acting factor and by cis-acting response elements. J. Virol. 1987, 61, 2462–2471. [Google Scholar] [CrossRef] [Green Version]
- Blazhko, N.; Vyshegurov, S.K.; Donchenko, A.; Shatokhin, K.; Krytsyna, T.; Ryabinina, V. Association of haplotypes for SNPs in the LTR regions of bovine leukemia virus with hematological indices of cattle. Vavilov J. Genet. Breed. 2019, 23, 162–169. [Google Scholar] [CrossRef] [Green Version]
- Katoh, I.; Kyushiki, H.; Sakamoto, Y.; Ikawa, Y.; Yoshinaka, Y. Bovine leukemia virus matrix-associated protein MA (p15): Further processing and formation of a specific complex with the dimer of the 5′-terminal genomic RNA fragment. J. Virol. 1991, 65, 6845–6855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamoun, R.; Morisson, M.; Rebeyrotte, N.; Busetta, B.; Couez, D.; Kettmann, R.; Guillemain, B. Sequence variability of bovine leukemia virus env gene and its relevance to the structure and antigenicity of the glycoproteins. J. Virol. 1990, 64, 4180–4188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llames, L.; Goyache, J.; Domenech, A.; Montaña, A.V.; Suarez, G.; Gomez-Lucia, E. Cellular distribution of bovine leukemia virus proteins gp51SU, Pr72env, and Pr66gag-pro in persistently infected cells. Virus Res. 2001, 79, 47–57. [Google Scholar] [CrossRef]
- Camargos, M.F.; Reis, J.K.; Leite, R.C. Bovine leukemia virus. Virus Rev. Res. 2004, 9, 44–59. [Google Scholar] [CrossRef]
- Lairmore, M.D. Animal models of bovine leukemia virus and human T-lymphotrophic virus type-1: Insights in transmission and pathogenesis. Annu. Rev. Anim. Biosci. 2014, 2, 189–208. [Google Scholar] [CrossRef]
- De Brogniez, A.; Bouzar, A.B.; Jacques, J.-R.; Cosse, J.-P.; Gillet, N.; Callebaut, I.; Reichert, M.; Willems, L. Mutation of a single envelope N-linked glycosylation site enhances the pathogenicity of bovine leukemia virus. J. Virol. 2015, 89, 8945–8956. [Google Scholar] [CrossRef] [Green Version]
- Rola-Łuszczak, M.; Sakhawat, A.; Pluta, A.; Ryło, A.; Bomba, A.; Bibi, N.; Kuźmak, J. Molecular Characterization of the env Gene of Bovine Leukemia Virus in Cattle from Pakistan with NGS-Based Evidence of Virus Heterogeneity. Pathogens 2021, 10, 910. [Google Scholar] [CrossRef]
- Licursi, M.; Inoshima, Y.; Wu, D.; Yokoyama, T.; González, E.T.; Sentsui, H. Provirus variants of bovine leukemia virus in naturally infected cattle from Argentina and Japan. Vet. Microbiol. 2003, 96, 17–23. [Google Scholar] [CrossRef]
- Lee, E.; Kim, E.-J.; Joung, H.-K.; Kim, B.-H.; Song, J.-Y.; Cho, I.-S.; Lee, K.-K.; Shin, Y.-K. Sequencing and phylogenetic analysis of the gp51 gene from Korean bovine leukemia virus isolates. Virol. J. 2015, 12, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-H.; Lee, E.-Y.; Oem, J.-K.; Kim, S.-H.; Lee, M.-H.; Lee, K.-K.; Park, S.-C. Genetic Analysis of Env and Gag Gene Fragments of Bovine Leukemia Virus Identified in Cattle from Korea. Korean J.Vet. Res. 2015, 55, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Marawan, M.A.; Mekata, H.; Hayashi, T.; Sekiguchi, S.; Kirino, Y.; Horii, Y.; Moustafa, A.-M.M.; Arnaout, F.K.; Galila, E.S.M.; Norimine, J. Phylogenetic analysis of env gene of bovine leukemia virus strains spread in Miyazaki prefecture, Japan. J. Vet. Med. Sci. 2017, 79, 912–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluta, A.; Rola-Łuszczak, M.; Kubiś, P.; Balov, S.; Moskalik, R.; Choudhury, B.; Kuźmak, J. Molecular characterization of bovine leukemia virus from Moldovan dairy cattle. Arch. Virol. 2017, 162, 1563–1576. [Google Scholar] [CrossRef] [Green Version]
- Polat, M.; Takeshima, S.-N.; Hosomichi, K.; Kim, J.; Miyasaka, T.; Yamada, K.; Arainga, M.; Murakami, T.; Matsumoto, Y.; Diaz, V.B. A new genotype of bovine leukemia virus in South America identified by NGS-based whole genome sequencing and molecular evolutionary genetic analysis. Retrovirology 2016, 13, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, D.T.; Yamashita-Kawanishi, N.; Okamoto, M.; Nguyen, S.V.; Nguyen, N.H.; Sugiura, K.; Miura, T.; Haga, T. Detection and genotyping of bovine leukemia virus (BLV) in Vietnamese cattle. J. Vet. Med. Sci. 2020, 82, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Hamada, R.; Metwally, S.; Polat, M.; Borjigin, L.; Ali, A.O.; Abdel-Hady, A.; Mohamed, A.E.; Wada, S.; Aida, Y. Detection and molecular characterization of bovine leukemia virus in Egyptian dairy cattle. Front. Vet. Sci. 2020, 7, 608. [Google Scholar] [CrossRef]
- Yu, C.; Wang, X.; Zhou, Y.; Wang, Y.; Zhang, X.; Zheng, Y. Genotyping bovine leukemia virus in dairy cattle of Heilongjiang, northeastern China. BMC Vet. Res. 2019, 15, 179. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.M.; Golemba, M.D.; Campos, R.H.; Trono, K.; Jones, L.R. Bovine leukemia virus can be classified into seven genotypes: Evidence for the existence of two novel clades. J. Gen. Virol. 2009, 90, 2788–2797. [Google Scholar] [CrossRef]
- Lee, E.; Kim, E.J.; Ratthanophart, J.; Vitoonpong, R.; Kim, B.H.; Cho, I.S.; Song, J.Y.; Lee, K.K.; Shin, Y.K. Molecular epidemiological and serological studies of bovine leukemia virus (BLV) infection in Thailand cattle. Infect. Genet. Evol. 2016, 41, 245–254. [Google Scholar] [CrossRef]
- Gautam, S.; Mishra, N.; Kalaiyarasu, S.; Jhade, S.K.; Sood, R. Molecular characterization of bovine leukaemia virus (BLV) strains reveals existence of genotype 6 in cattle in India with evidence of a new subgenotype. Transbound Emerg. Dis. 2018, 65, 1968–1978. [Google Scholar] [CrossRef] [PubMed]
- Selim, A.; Manaa, E.; Alanazi, A.; Alyousif, M. Seroprevalence, Risk Factors and Molecular Identification of Bovine Leukemia Virus in Egyptian Cattle. Animals 2021, 11, 319. [Google Scholar] [CrossRef]
- Fenner, F.J.; Bachmann, P.A.; Gibbs, E.P.J. Veterinary Virology; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Inoue, E.; Matsumura, K.; Maekawa, K.; Nagatsuka, K.; Nobuta, M.; Hirata, M.; Minagawa, A.; Osawa, Y.; Okazaki, K. Genetic heterogeneity among bovine leukemia viruses in Japan and their relationship to leukemogenicity. Arch. Virol. 2011, 156, 1137–1141. [Google Scholar] [CrossRef]
- Polat, M.; Takeshima, S.-N.; Aida, Y. Epidemiology and genetic diversity of bovine leukemia virus. Virol. J. 2017, 14, 209. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, L.; Dong, M.; Huang, W.; Hao, X.; Peng, Y.; Gong, Z.; Qin, A.; Shang, S.; Yang, Z. Molecular characterization of bovine leukemia virus reveals existence of genotype 4 in Chinese dairy cattle. Virol. J. 2019, 16, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Phiri, M.M.; Kaimoyo, E.; Changula, K.; Silwamba, I.; Chambaro, H.M.; Kapila, P.; Kajihara, M.; Simuunza, M.; Muma, J.B.; Pandey, G.S. Molecular detection and characterization of genotype 1 bovine leukemia virus from beef cattle in the traditional sector in Zambia. Arch. Virol. 2019, 164, 2531–2536. [Google Scholar] [CrossRef]
- Kazemimanesh, M.; Madadgar, O.; Steinbach, F.; Choudhury, B.; Azadmanesh, K. Detection and molecular characterization of bovine leukemia virus in various regions of Iran. J. Gen. Virol. 2019, 100, 1315–1327. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Chapman, R.; Douglass, N.; Carulei, O.; van Rensburg, J.; Williamson, A.-L. Phylogenetic Analysis of South African Bovine Leukaemia Virus (BLV) Isolates. Viruses 2020, 12, 898. [Google Scholar] [CrossRef]
- Úsuga-Monroy, C.; Díaz, F.J.; Echeverri-Zuluaga, J.J.; González-Herrera, L.G.; López-Herrera, A. Presence of bovine leukemia virus genotypes 1 and 3 in Antioquia, Colombia. Rev. UDCA Actual. Divulg. Científica 2018, 21, 119–126. [Google Scholar]
- Moe, K.K.; Polat, M.; Borjigin, L.; Matsuura, R.; Hein, S.T.; Moe, H.H.; Aida, Y. New evidence of bovine leukemia virus circulating in Myanmar cattle through epidemiological and molecular characterization. PLoS ONE 2020, 15, e0229126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arainga, M.; Takeda, E.; Aida, Y. Identification of bovine leukemia virus tax function associated with host cell transcription, signaling, stress response and immune response pathway by microarray-based gene expression analysis. BMC Genom. 2012, 13, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zyrianova, I.M.; Kovalchuk, S.N. Bovine leukemia virus tax gene/Tax protein polymorphism and its relation to Enzootic Bovine Leukosis. Virulence 2020, 11, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Kerkhofs, P.; Heremans, H.; Burny, A.; Kettmann, R.; Willems, L. In vitro and in vivo oncogenic potential of bovine leukemia virus G4 protein. J. Virol. 1998, 72, 2554–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barez, P.-Y.; De Brogniez, A.; Carpentier, A.; Gazon, H.; Gillet, N.; Gutiérrez, G.; Hamaidia, M.; Jacques, J.-R.; Perike, S.; Neelature Sriramareddy, S. Recent advances in BLV research. Viruses 2015, 7, 6080–6088. [Google Scholar] [CrossRef]
- Kincaid, R.P.; Burke, J.M.; Sullivan, C.S. RNA virus microRNA that mimics a B-cell oncomiR. Proc. Natl. Acad. Sci. USA 2012, 109, 3077–3082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosewick, N.; Momont, M.; Durkin, K.; Takeda, H.; Caiment, F.; Cleuter, Y.; Vernin, C.; Mortreux, F.; Wattel, E.; Burny, A. Deep sequencing reveals abundant noncanonical retroviral microRNAs in B-cell leukemia/lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 2306–2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillet, N.A.; Hamaidia, M.; De Brogniez, A.; Gutierrez, G.; Renotte, N.; Reichert, M.; Trono, K.; Willems, L. Bovine leukemia virus small noncoding RNAs are functional elements that regulate replication and contribute to oncogenesis in vivo. PLoS Pathog. 2016, 12, e1005588. [Google Scholar] [CrossRef]
- Durkin, K.; Rosewick, N.; Artesi, M.; Hahaut, V.; Griebel, P.; Arsic, N.; Burny, A.; Georges, M.; Van den Broeke, A. Characterization of novel Bovine Leukemia Virus (BLV) antisense transcripts by deep sequencing reveals constitutive expression in tumors and transcriptional interaction with viral microRNAs. Retrovirology 2016, 13, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Burng, A. Bovine leukemia virus: Molecular biology and epidemiology. Viral Oncol. 1980, 1980, 231–289. [Google Scholar]
- Rodríguez, S.M.; Florins, A.; Gillet, N.; De Brogniez, A.; Sánchez-Alcaraz, M.T.; Boxus, M.; Boulanger, F.; Gutiérrez, G.; Trono, K.; Alvarez, I. Preventive and therapeutic strategies for bovine leukemia virus: Lessons for HTLV. Viruses 2011, 3, 1210–1248. [Google Scholar] [CrossRef]
- MAFF. The National Animal Health Statistics; MAFF: Tokyo, Japan, 2011. (In Japanese) [Google Scholar]
- Nekouei, O.; VanLeeuwen, J.; Sanchez, J.; Kelton, D.; Tiwari, A.; Keefe, G. Herd-level risk factors for infection with bovine leukemia virus in Canadian dairy herds. Prev. Vet. Med. 2015, 119, 105–113. [Google Scholar] [CrossRef]
- Şevik, M.; Avcı, O.; İnce, Ö.B. An 8-year longitudinal sero-epidemiological study of bovine leukaemia virus (BLV) infection in dairy cattle in Turkey and analysis of risk factors associated with BLV seropositivity. Trop. Anim. Health Prod. 2015, 47, 715–720. [Google Scholar] [CrossRef]
- Heinecke, N.; Tórtora, J.; Martínez, H.A.; Gonzalez-Fernandez, V.D.; Ramírez, H. Detection and genotyping of bovine leukemia virus in Mexican cattle. Arch. Virol. 2017, 162, 3191–3196. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.-Y.; Gong, Q.-L.; Sheng, C.-Y.; Liu, Y.; Ge, G.-Y.; Li, D.-L.; Diao, N.-C.; Shi, K.; Li, J.-M.; Sun, Z.-B. Prevalence of bovine leukemia in 1983–2019 in China: A systematic review and meta-analysis. Microb. Pathog. 2020, 2020, 104681. [Google Scholar]
- Zaher, K.S.; Ahmed, W.M. Bovine leukemia virus infection in dairy cows in Egypt. Acad. J. Cancer Res. 2014, 7, 126–130. [Google Scholar]
- Bartlett, P.C.; Sordillo, L.M.; Byrem, T.M.; Norby, B.; Grooms, D.L.; Swenson, C.L.; Zalucha, J.; Erskine, R.J. Options for the control of bovine leukemia virus in dairy cattle. J. Am. Vet. Med. Assoc. 2014, 244, 914–922. [Google Scholar] [CrossRef]
- Whittington, R.; Donat, K.; Weber, M.F.; Kelton, D.; Nielsen, S.S.; Eisenberg, S.; Arrigoni, N.; Juste, R.; Sáez, J.L.; Dhand, N. Control of paratuberculosis: Who, why and how. A review of 48 countries. BMC Vet. Res. 2019, 15, 198. [Google Scholar] [CrossRef] [Green Version]
- Ruggiero, V.J.; Bartlett, P.C. Control of bovine leukemia virus in three US dairy herds by culling ELISA-positive cows. Vet. Med. Int. 2019, 2019, 3202184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- More, S.; Bøtner, A.; Butterworth, A.; Calistri, P.; Depner, K.; Edwards, S.; Garin-Bastuji, B.; Good, M.; Schmidt, C.G.; Michel, V. Assessment of listing and categorisation of animal diseases within the framework of the Animal Health Law (Regulation (EU) No 2016/429): Enzootic bovine leukosis (EBL). EFSA J. 2017, 15, 4956. [Google Scholar]
- OIE. World Animal Health Infromation Database-Version: 1.4; World Organisation for Animal Health: Paris, France, 2009. [Google Scholar]
- OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals: Chapter 2.4.11. Enzootic Bovine Leukosis, 7th ed.; World Organisation for Animal Health: Paris, France, 2012. [Google Scholar]
- Chethanond, U.-S. The Epidemiology of Enzootic Bovine Leukosis in Dairy Cattle in New Zealand: A Thesis Presented in Partial Fulfilment of the Requirements for the Degree of. Master of Veterinary Science at Massey University. Doctoral Dissertation, Massey University, Palmerston North, New Zealand, 1999. [Google Scholar]
- Balic, D.; Lojkic, I.; Periskic, M.; Bedekovic, T.; Jungic, A.; Lemo, N.; Roic, B.; Cac, Z.; Barbic, L.; Madic, J. Identification of a new genotype of bovine leukemia virus. Arch. Virol. 2012, 157, 1281–1290. [Google Scholar] [CrossRef]
- Rola-Luszczak, M.; Pluta, A.; Olech, M.; Donnik, I.; Petropavlovskiy, M.; Gerilovych, A.; Vinogradova, I.; Choudhury, B.; Kuzmak, J. The molecular characterization of bovine leukaemia virus isolates from eastern Europe and Siberia and its impact on phylogeny. PLoS ONE 2013, 8, e58705. [Google Scholar] [CrossRef]
- Moratorio, G.; Obal, G.; Dubra, A.; Correa, A.; Bianchi, S.; Buschiazzo, A.; Cristina, J.; Pritsch, O. Phylogenetic analysis of bovine leukemia viruses isolated in South America reveals diversification in seven distinct genotypes. Arch. Virol. 2010, 155, 481–489. [Google Scholar] [CrossRef] [PubMed]
- APHIS. Bovine Leukosis Virus (BLV) on U.S. Dairy Operations, 2007; United States Department of Agriculture: Washington, DC, USA, 2008; Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.739.3674&rep=rep1&type=pdf (accessed on 23 October 2021).
- SUZAN, V.M.; ONUMA, M.; AGUILAR, R.E.; MURAKAMI, Y. Prevalence of bovine herpesvirus-1, parainfluenza-3, bovine rotavirus, bovine viral diarrhea, bovine adenovirus-7, bovine leukemia virus and bluetongue virus antibodies in cattle in Mexico. Jpn. J. Vet. Res. 1983, 31, 125–132. [Google Scholar]
- Samara, S.I.; Lima, E.G.; do Nascimento, A.A. Monitoring of enzootic bovine leukosis in dairy cattle from the Pitangueiras region in São Paulo, Brazil. Braz. J. Vet. Res. Anim. Sci. 1997, 34, 349–351. [Google Scholar] [CrossRef] [Green Version]
- D’angelino, J.; Garcia, M.; Birgel, E. Epidemiological study of enzootic bovine leukosis in Brazil. Trop. Anim. Health Prod. 1998, 30, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Trono, K.G.; Pérez-Filgueira, D.M.; Duffy, S.; Borca, M.V.; Carrillo, C. Seroprevalence of bovine leukemia virus in dairy cattle in Argentina: Comparison of sensitivity and specificity of different detection methods. Vet. Microbiol. 2001, 83, 235–248. [Google Scholar] [CrossRef]
- Hung Ch, A. Bovine leukaemia virus infection in Peru. Trop. Anim. Health Prod. 1983, 15, 61. [Google Scholar] [CrossRef] [PubMed]
- Marin, C.; de López, N.M.; Alvarez, L.; Lozano, O.; España, W.; Castaños, H.; León, A. Epidemiology of bovine leukemia in Venezuela. Ann. De Rech. Vét. 1978, 9, 743–746. [Google Scholar]
- Corredor-Figueroa, A.P.; Salas, S.; Olaya-Galán, N.N.; Quintero, J.S.; Fajardo, Á.; Soñora, M.; Moreno, P.; Cristina, J.; Sánchez, A.; Tobón, J. Prevalence and molecular epidemiology of bovine leukemia virus in Colombian cattle. Infect. Genet. Evol. 2020, 80, 104171. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fan, W.; Mao, Y.; Yang, Z.; Lu, G.; Zhang, R.; Zhang, H.; Szeto, C.; Wang, C. Bovine leukemia virus infection in cattle of China: Association with reduced milk production and increased somatic cell score. J. Dairy Sci. 2016, 99, 3688–3697. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, J.-C.; Li, C.-Y.; Hsu, W.-L.; Chuang, S.-T. Molecular epidemiological and serological studies of bovine leukemia virus in Taiwan dairy cattle. Front. Vet. Sci. 2019, 6, 427. [Google Scholar] [CrossRef] [Green Version]
- Meas, S.; Ohashi, K.; Tum, S.; Chhin, M.; Te, K.; Miura, K.; Sugimoto, C.; Onuma, M. Seroprevalence of bovine immunodeficiency virus and bovine leukemia virus in draught animals in Cambodia. J. Vet. Med. Sci. 2000, 62, 779–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, K.; Kobayashi, S.; Konishi, M.; Kameyama, K.-I.; Tsutsui, T. Nationwide survey of bovine leukemia virus infection among dairy and beef breeding cattle in Japan from 2009–2011. J. Vet. Med. Sci. 2013, 75, 1123–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, S.; Tsutsui, T.; Yamamoto, T.; Hayama, Y.; Kameyama, K.; Konishi, M.; Murakami, K. Risk factors associated with within-herd transmission of bovine leukemia virus on dairy farms in Japan. BMC Vet. Res. 2010, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, A.; Takeshima, S.N.; Matsumoto, Y.; Aida, Y. Risk factors associated with increased bovine leukemia virus proviral load in infected cattle in Japan from 2012 to 2014. Virus Res. 2015, 210, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Ochirkhuu, N.; Konnai, S.; Odbileg, R.; Nishimori, A.; Okagawa, T.; Murata, S.; Ohashi, K. Detection of bovine leukemia virus and identification of its genotype in Mongolian cattle. Arch. Virol. 2016, 161, 985–991. [Google Scholar] [CrossRef]
- Polat, M.; Ohno, A.; Takeshima, S.N.; Kim, J.; Kikuya, M.; Matsumoto, Y.; Mingala, C.N.; Onuma, M.; Aida, Y. Detection and molecular characterization of bovine leukemia virus in Philippine cattle. Arch. Virol. 2015, 160, 285–296. [Google Scholar] [CrossRef]
- Khan, M.F.; Siddique, U.; Shah, A.A.; Khan, I.; Anwar, F.; Ahmad, I.; Zeb, M.T.; Hassan, M.F.; Ali, T. Seroprevalence of Bovine Leukemia Virus (BLV) in Cattle from the North West of Pakistan. Pak. Vet. J. 2019, 40, 127–129. [Google Scholar]
- Hafez, S.; Sharif, M.; Al-Sukayran, A.; Dela-Cruz, D. Preliminary studies on enzootic bovine leukosis in Saudi dairy farms. DTW Dtsch. Tierarztl. Wochenschr. 1990, 97, 61–63. [Google Scholar]
- Burgu, I.; Alkan, F.; Karaoglu, T.; Bilge-Dagalp, S.; Can-Sahna, K.; Güngör, B.; Demir, B. Control and eradication programme of enzootic bovine leucosis (EBL) from selected dairy herds in Turkey. DTW Dtsch. Tierarztl. Wochenschr. 2005, 112, 271–274. [Google Scholar] [PubMed]
- Trainin, Z.; Brenner, J. The direct and indirect economic impacts of bovine leukemia virus infection on dairy cattle. Isr. J. Vet. Med. 2005, 60, 94. [Google Scholar]
- Khudhair, Y.I.; Hasso, S.A.; Yaseen, N.Y.; Al-Shammari, A.M. Serological and molecular detection of bovine leukemia virus in cattle in Iraq. Emerg. Microbes Infect. 2016, 5, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selim, A.; Megahed, A.A.; Kandeel, S.; Abdelhady, A. Risk factor analysis of bovine leukemia virus infection in dairy cattle in Egypt. Comp. Immunol. Microbiol. Infect. Dis. 2020, 72, 101517. [Google Scholar] [CrossRef] [PubMed]
- Lewin, H.A.; Bernoco, D. Evidence for BoLA-linked resistance and susceptibility to subclinical progression of bovine leukaemia virus infection. Anim. Genet. 1986, 17, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Etxebarria, K.; Sistiaga-Poveda, M.; Jugo, B.M. Endogenous retroviruses in domestic animals. Curr. Genom. 2014, 15, 256–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Angelino, R.H.R.; Pituco, E.M.; Villalobos, E.M.C.; Harakava, R.; Gregori, F.; Del Fava, C. Detection of bovine leukemia virus in brains of cattle with a neurological syndrome: Pathological and molecular studies. BioMed Res. Int. 2013, 2013, 425646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kettmann, R.; Burny, A.; Callebaut, I.; Droogmans, L.; Mammerickx, M.; Willems, L.; Portetelle, D. Bovine leukemia virus. In The Retroviridae; Springer: Berlin/Heidelberg, Germany, 1994; pp. 39–81. [Google Scholar]
- Porta, N.G.; Alvarez, I.; Archilla, G.S.; Ruiz, V.; Abdala, A.; Trono, K. Experimental infection of sheep with Bovine leukemia virus (BLV): Minimum dose of BLV-FLK cells and cell-free BLV and neutralization activity of natural antibodies. Rev. Argent. Microbiol. 2019, 51, 316–323. [Google Scholar] [CrossRef]
- Dimitrov, P.; Simeonov, K.; Todorova, K.; Ivanova, Z.; Toshkova, R.; Shikova, E.; Russev, R. Pathological features of experimental bovine leukaemia viral (BLV) infection in rats and rabbits. Bull. Vet. Inst. Pulawy 2012, 56, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, E.; Weigel, K.; Byrem, T.; Rosa, G. Genetic correlation of bovine leukosis incidence with somatic cell score and milk yield in a US Holstein population. J. Dairy Sci. 2016, 99, 2005–2009. [Google Scholar] [CrossRef] [Green Version]
- Burmeister, T. Oncogenic retroviruses in animals and humans. Rev. Med. Virol. 2001, 11, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Bauermann, F.V.; Ridpath, J.F.; Dargatz, D.A. Bovine leukemia virus seroprevalence among cattle presented for slaughter in the United States. J. Vet. Diagn. Investig. 2017, 29, 704–706. [Google Scholar] [CrossRef] [Green Version]
- Murakami, K.; Kobayashi, S.; Konishi, M.; Kameyama, K.-i.; Yamamoto, T.; Tsutsui, T. The recent prevalence of bovine leukemia virus (BLV) infection among Japanese cattle. Vet. Microbiol. 2011, 148, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Maezawa, M.; Watanabe, K.-i.; Horiuchi, N.; Matsumoto, K.; Kobayashi, Y.; Inokuma, H. A clinical case of enzootic bovine leukosis in a 13-month-old Holstein heifer. Jpn. J. Vet. Res. 2018, 66, 209–213. [Google Scholar]
- Oguma, K.; Suzuki, M.; Sentsui, H. Enzootic bovine leukosis in a two-month-old calf. Virus Res. 2017, 233, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Shaghayegh, A. Detection and identification of Enzootic Bovine Leukosis (EBL) in Calves in Iran. Arch. Razi Inst. 2019, 74, 321–325. [Google Scholar] [PubMed]
- Hutchinson, H.C.; Ruggiero, V.J.; Norby, B.; Sporer, K.R.B.; Bartlett, P.C. Diagnostic Measures of Disease Progression in Cattle Following Natural Infection with Bovine Leukemia Virus. Pathogens 2021, 10, 987. [Google Scholar] [CrossRef]
- Watanuki, S.; Takeshima, S.N.; Borjigin, L.; Sato, H.; Bai, L.; Murakami, H.; Sato, R.; Ishizaki, H.; Matsumoto, Y.; Aida, Y. Visualizing bovine leukemia virus (BLV)-infected cells and measuring BLV proviral loads in the milk of BLV seropositive dams. Vet. Res. 2019, 50, 102. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Tsutsui, T.; Yamamoto, T.; Hayama, Y.; Muroga, N.; Konishi, M.; Kameyama, K.-i.; Murakami, K. The role of neighboring infected cattle in bovine leukemia virus transmission risk. J. Vet. Med. Sci. 2015, 77, 861–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Kitamura-Muramatsu, Y.; Saito, S.; Ishizaki, H.; Nakano, M.; Haga, S.; Matoba, K.; Ohno, A.; Murakami, H.; Takeshima, S.N.; et al. Detection of the BLV provirus from nasal secretion and saliva samples using BLV-CoCoMo-qPCR-2: Comparison with blood samples from the same cattle. Virus Res. 2015, 210, 248–254. [Google Scholar] [CrossRef]
- Foley, C.; O’Farrelly, C.; Meade, K.G. Technical note: Comparative analyses of the quality and yield of genomic DNA from invasive and noninvasive, automated and manual extraction methods. J. Dairy Sci. 2011, 94, 3159–3165. [Google Scholar] [CrossRef]
- Mammerickx, M.; Portetelle, D.; de Clercq, K.; Burny, A. Experimental transmission of enzootic bovine leukosis to cattle, sheep and goats: Infectious doses of blood and incubation period of the disease. Leuk. Res. 1987, 11, 353–358. [Google Scholar] [CrossRef]
- Mekata, H.; Sekiguchi, S.; Konnai, S.; Kirino, Y.; Honkawa, K.; Nonaka, N.; Horii, Y.; Norimine, J. Evaluation of the natural perinatal transmission of bovine leukaemia virus. Vet. Rec. 2015, 176, 254. [Google Scholar] [CrossRef] [PubMed]
- Haga, T. Enzootic Bovine Leukosis: How to prevent the disease and control the spread of BLV infection. In Proceedings of the 1st International Conference Postgraduate School Universitas Airlangga: “Implementation of Climate Change Agreement to Meet Sustainable Development Goals” (ICPSUAS 2017), Surabaya, Indonesia, 1–2 August 2017. [Google Scholar]
- Kohara, J.; Takeuchi, M.; Hirano, Y.; Sakurai, Y.; Takahashi, T. Vector control efficacy of fly nets on preventing bovine leukemia virus transmission. J. Vet. Med. Sci. 2018, 80, 1524–1527. [Google Scholar] [CrossRef] [Green Version]
- Ooshiro, M.; Konnai, S.; Katagiri, Y.; Afuso, M.; Arakaki, N.; Tsuha, O.; Murata, S.; Ohashi, K. Horizontal transmission of bovine leukemia virus from lymphocytotic cattle, and beneficial effects of insect vector control. Vet. Rec. 2013, 173, 527. [Google Scholar] [CrossRef] [PubMed]
- Selim, A.; Radwan, A. Seroprevalence and molecular characterization of West Nile Virus in Egypt. Comp. Immunol. Microbiol. Infect. Dis. 2020, 71, 101473. [Google Scholar] [CrossRef] [PubMed]
- Selim, A.; Ali, A.-F. Seroprevalence and risk factors for C. burentii infection in camels in Egypt. Comp. Immunol. Microbiol. Infect. Dis. 2020, 68, 101402. [Google Scholar] [CrossRef] [PubMed]
- Mekata, H.; Yamamoto, M.; Hayashi, T.; Kirino, Y.; Sekiguchi, S.; Konnai, S.; Horii, Y.; Norimine, J. Cattle with a low bovine leukemia virus proviral load are rarely an infectious source. Jpn. J. Vet. Res. 2018, 66, 157–163. [Google Scholar]
- Konishi, M.; Ishizaki, H.; Kameyama, K.-I.; Murakami, K.; Yamamoto, T. The effectiveness of colostral antibodies for preventing bovine leukemia virus (BLV) infection in vitro. BMC Vet. Res. 2018, 14, 419. [Google Scholar] [CrossRef] [PubMed]
- Andoh, K.; Nishimori, A.; Sakumoto, R.; Hayashi, K.-G.; Hatama, S. The chemokines CCL2 and CXCL10 produced by bovine endometrial epithelial cells induce migration of bovine B lymphocytes, contributing to transuterine transmission of BLV infection. Vet. Microbiol. 2020, 242, 108598. [Google Scholar] [CrossRef]
- Erskine, R.J.; Bartlett, P.C.; Byrem, T.M.; Render, C.L.; Febvay, C.; Houseman, J.T. Herd-level determinants of bovine leukaemia virus prevalence in dairy farms. J. Dairy Res. 2012, 79, 445–450. [Google Scholar] [CrossRef]
- Benitez, O.J.; Roberts, J.N.; Norby, B.; Bartlett, P.C.; Takeshima, S.-N.; Watanuki, S.; Aida, Y.; Grooms, D.L. Breeding bulls as a potential source of bovine leukemia virus transmission in beef herds. J. Am. Vet. Med. Assoc. 2019, 254, 1335–1340. [Google Scholar] [CrossRef]
- Stott, M.L.; Thurmond, M.C.; Dunn, S.J.; Osburn, B.I.; Stott, J.L. Integrated bovine leukosis proviral DNA in T helper and T cytotoxic/suppressor lymphocytes. J. Gen. Virol. 1991, 72 Pt 2, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Dube, S.; Bachman, S.; Spicer, T.; Love, J.; Choi, D.; Esteban, E.; Ferrer, J.F.; Poiesz, B.J. Degenerate and specific PCR assays for the detection of bovine leukaemia virus and primate T cell leukaemia/lymphoma virus pol DNA and RNA: Phylogenetic comparisons of amplified sequences from cattle and primates from around the world. J. Gen. Virol. 1997, 78, 1389–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantor, G.H.; Pritchard, S.M.; Dequiedt, F.; Willems, L.; Kettmann, R.; Davis, W.C. CD5 is dissociated from the B-cell receptor in B cells from bovine leukemia virus-infected, persistently lymphocytotic cattle: Consequences to B-cell receptor-mediated apoptosis. J. Virol. 2001, 75, 1689–1696. [Google Scholar] [CrossRef] [Green Version]
- Park, B.H.; Matuschke, B.; Lavi, E.; Gaulton, G.N. A point mutation in the env gene of a murine leukemia virus induces syncytium formation and neurologic disease. J. Virol. 1994, 68, 7516–7524. [Google Scholar] [CrossRef] [Green Version]
- Willems, L.; Burny, A.; Collete, D.; Dangoisse, O.; Dequiedt, F.; Gatot, J.S.; Kerkhofs, P.; Lefebvre, L.; Merezak, C.; Peremans, T.; et al. Genetic determinants of bovine leukemia virus pathogenesis. AIDS Res. Hum. Retrovir. 2000, 16, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Gatot, J.S.; Callebaut, I.; Van Lint, C.; Demonte, D.; Kerkhofs, P.; Portetelle, D.; Burny, A.; Willems, L.; Kettmann, R. Bovine leukemia virus SU protein interacts with zinc, and mutations within two interacting regions differently affect viral fusion and infectivity in vivo. J. Virol. 2002, 76, 7956–7967. [Google Scholar] [CrossRef] [Green Version]
- Camargos, M.F.; Pereda, A.; Stancek, D.; Rocha, M.A.; dos Reis, J.K.; Greiser-Wilke, I.; Leite, R.C. Molecular characterization of the env gene from Brazilian field isolates of Bovine leukemia virus. Virus Genes 2007, 34, 343–350. [Google Scholar] [CrossRef]
- Tanaka, A.S.; Komuro, K. Targeted rearrangement of a chromosomal repeat sequence by transfection of a homologous DNA sequence using purified integrase. Gene Ther. 2005, 12, 783–794. [Google Scholar] [CrossRef]
- EFSA Panel on Animal Health and Welfare (AHAW). Scientific Opinion on Enzootic Bovine Leukosis. Eur. Food Saf. Auth. (EFSA) 2015, 13, 4188. [Google Scholar]
- De Brogniez, A.; Mast, J.; Willems, L. Determinants of the Bovine Leukemia Virus Envelope Glycoproteins Involved in Infectivity, Replication and Pathogenesis. Viruses 2016, 8, 88. [Google Scholar] [CrossRef] [Green Version]
- Gillet, N.; Florins, A.; Boxus, M.; Burteau, C.; Nigro, A.; Vandermeers, F.; Balon, H.; Bouzar, A.-B.; Defoiche, J.; Burny, A. Mechanisms of leukemogenesis induced by bovine leukemia virus: Prospects for novel anti-retroviral therapies in human. Retrovirology 2007, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, I.; Levy, D. Pathobiology of bovine leukemia virus. Vet. Res. 1994, 25, 521–536. [Google Scholar] [PubMed]
- Alvarez, I.; Gutiérrez, G.; Gammella, M.; Martínez, C.; Politzki, R.; González, C.; Caviglia, L.; Carignano, H.; Fondevila, N.; Poli, M.; et al. Evaluation of total white blood cell count as a marker for proviral load of bovine leukemia virus in dairy cattle from herds with a high seroprevalence of antibodies against bovine leukemia virus. Am. J. Vet. Res. 2013, 74, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Kahn, C.; Line, S. The Merck Veterinary Manual. Bovine Leukosis; Merck and Company: White House Station, NJ, USA, 2010. [Google Scholar]
- Zaghawa, A.; Beier, D.; Abd El-Rahim, I.; El-Ballal, S.; Karim, I.; Conraths, F.; Marquardt, O. An outbreak of enzootic bovine leukosis in upper egypt: Clinical, laboratory and molecular–epidemiological studies. J. Vet. Med. Ser. B 2002, 49, 123–129. [Google Scholar] [CrossRef]
- Sparling, A.M. An unusual presentation of enzootic bovine leukosis. Can. Vet. J. 2000, 41, 315. [Google Scholar] [PubMed]
- Uchiyama, J.; Murakami, H.; Sato, R.; Mizukami, K.; Suzuki, T.; Shima, A.; Ishihara, G.; Sogawa, K.; Sakaguchi, M. Examination of the fecal microbiota in dairy cows infected with bovine leukemia virus. Vet. Microbiol. 2020, 240, 108547. [Google Scholar] [CrossRef]
- Blagitz, M.G.; Souza, F.N.; Batista, C.F.; Azevedo, L.F.F.; Sanchez, E.M.R.; Diniz, S.A.; Silva, M.X.; Haddad, J.P.; Della Libera, A.M.M.P. Immunological implications of bovine leukemia virus infection. Res. Vet. Sci. 2017, 114, 109–116. [Google Scholar] [CrossRef]
- Patel, J.R.; Heldens, J.G.M.; Bakonyi, T.; Rusvai, M. Important mammalian veterinary viral immunodiseases and their control. Vaccine 2012, 30, 1767–1781. [Google Scholar] [CrossRef]
- Ferrer, J.F.; Kenyon, S.J.; Gupta, P. Milk of dairy cows frequently contains a leukemogenic virus. Science 1981, 213, 1014–1016. [Google Scholar] [CrossRef]
- McClure, H.M.; Keeling, M.E.; Custer, R.P.; Marshak, R.R.; Abt, D.A.; Ferrer, J.F. Erythroleukemia in two infant chimpanzees fed milk from cows naturally infected with the bovine C-type virus. Cancer Res. 1974, 34, 2745–2757. [Google Scholar]
- Johnson, R.; Kaneene, J.B. Bovine leukemia virus and enzootic bovine leukosis. Vet. Bull. 1992, 62, 287–312. [Google Scholar]
- Bender, A.P.; Robison, L.L.; Kashmiri, S.; McClain, K.L.; Woods, W.G.; Smithson, W.A.; Heyn, R.; Finlay, J.; Schuman, L.M.; Renier, C. No involvement of bovine leukemia virus in childhood acute lymphoblastic leukemia and non-Hodgkin’s lymphoma. Cancer Res. 1988, 48, 2919–2922. [Google Scholar] [PubMed]
- Lee, J.; Kim, Y.; Kang, C.S.; Cho, D.H.; Shin, D.H.; Yum, Y.N.; Oh, J.H.; Kim, S.H.; Hwang, M.S.; Lim, C.J. Investigation of the bovine leukemia virus proviral DNA in human leukemias and lung cancers in Korea. J. Korean Med. Sci. 2005, 20, 603–606. [Google Scholar] [CrossRef]
- Zhang, R.; Jiang, J.; Sun, W.; Zhang, J.; Huang, K.; Gu, X.; Yang, Y.; Xu, X.; Shi, Y.; Wang, C. Lack of association between bovine leukemia virus and breast cancer in Chinese patients. Breast Cancer Res. 2016, 18, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillet, N.A.; Willems, L. Whole genome sequencing of 51 breast cancers reveals that tumors are devoid of bovine leukemia virus DNA. Retrovirology 2016, 13, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohara, J.; Konnai, S.; Onuma, M. Experimental transmission of Bovine leukemia virus in cattle via rectal palpation. Jpn. J. Vet. Res. 2006, 54, 25–30. [Google Scholar]
- Jimba, M.; Takeshima, S.N.; Murakami, H.; Kohara, J.; Kobayashi, N.; Matsuhashi, T.; Ohmori, T.; Nunoya, T.; Aida, Y. BLV-CoCoMo-qPCR: A useful tool for evaluating bovine leukemia virus infection status. BMC Vet. Res. 2012, 8, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monti, G.E.; Frankena, K.; Engel, B.; Buist, W.; Tarabla, H.D.; de Jong, M.C. Evaluation of a new antibody-based enzyme-linked immunosorbent assay for the detection of bovine leukemia virus infection in dairy cattle. J. Vet. Diagn. Investig. 2005, 17, 451–457. [Google Scholar] [CrossRef] [Green Version]
- Naif, H.M.; Daniel, R.C.; Cougle, W.G.; Lavin, M.F. Early detection of bovine leukemia virus by using an enzyme-linked assay for polymerase chain reaction-amplified proviral DNA in experimentally infected cattle. J. Clin. Microbiol. 1992, 30, 675–679. [Google Scholar] [CrossRef] [Green Version]
- Kurdi, A.; Blankenstein, P.; Marquardt, O.; Ebner, D. Serologic and virologic investigations on the presence of BLV infection in a dairy herd in Syria. Berl. Munch Tierarztl. Wochenschr. 1999, 112, 18–23. [Google Scholar]
- Levy, D.; Deshayes, L.; Parodi, A.L.; Levy, J.P.; Stephenson, J.R.; Devare, S.G.; Gilden, R.V. Bovine leukemia virus specific antibodies among French cattle. II. Radioimmunoassay with the major structural protein (BLV p24). Int. J. Cancer 1977, 20, 543–550. [Google Scholar] [CrossRef]
- Nguyen, V.K.; Maes, R.F. Evaluation of an enzyme-linked immunosorbent assay for detection of antibodies to bovine leukemia virus in serum and milk. J. Clin. Microbiol. 1993, 31, 979–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukai, K.; Sato, M.; Kawara, M.; Hoshi, Z.; Ueno, S.; Chyou, N.; Akashi, H. A case of an embryo transfer calf infected with bovine leukemia virus from the recipient cow. Zent. Vet. B 1999, 46, 511–515. [Google Scholar] [CrossRef]
- Jimba, M.; Takeshima, S.; Matoba, K.; Endoh, D.; Aida, Y. BLV-CoCoMo-qPCR: Quantitation of bovine leukemia virus proviral load using the CoCoMo algorithm. Retrovirology 2010, 7, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somura, Y.; Sugiyama, E.; Fujikawa, H.; Murakami, K. Comparison of the copy numbers of bovine leukemia virus in the lymph nodes of cattle with enzootic bovine leukosis and cattle with latent infection. Arch. Virol. 2014, 159, 2693–2697. [Google Scholar] [CrossRef] [PubMed]
- Lew, A.E.; Bock, R.E.; Miles, J.; Cuttell, L.B.; Steer, P.; Nadin-Davis, S.A. Sensitive and specific detection of bovine immunodeficiency virus and bovine syncytial virus by 5′ Taq nuclease assays with fluorescent 3′ minor groove binder-DNA probes. J. Virol. Methods 2004, 116, 1–9. [Google Scholar] [CrossRef]
- Tajima, S.; Ikawa, Y.; Aida, Y. Complete bovine leukemia virus (BLV) provirus is conserved in BLV-infected cattle throughout the course of B-cell lymphosarcoma development. J. Virol. 1998, 72, 7569–7576. [Google Scholar] [CrossRef] [Green Version]
- Tajima, S.; Tsukamoto, M.; Aida, Y. Latency of viral expression in vivo is not related to CpG methylation in the U3 region and part of the R region of the long terminal repeat of bovine leukemia virus. J. Virol. 2003, 77, 4423–4430. [Google Scholar] [CrossRef] [Green Version]
- Takeshima, S.-N.; Watanuki, S.; Ishizaki, H.; Matoba, K.; Aida, Y. Development of a direct blood-based PCR system to detect BLV provirus using CoCoMo primers. Arch. Virol. 2016, 161, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Tawfeeq, M.M.; Horiuchi, N.; Kobayashi, Y.; Furuoka, H.; Inokuma, H. Evaluation of gene expression in peripheral blood cells as a potential biomarker for enzootic bovine leukosis. J. Vet. Med. Sci. 2013, 75, 1213–1217. [Google Scholar] [CrossRef] [Green Version]
- El Daous, H.; Mitoma, S.; Elhanafy, E.; Thi Nguyen, H.; Thi Mai, N.; Hara, A.; Duangtathip, K.; Takezaki, Y.; Kaneko, C.; Norimine, J. Establishment of a novel diagnostic test for Bovine leukaemia virus infection using direct filter PCR. Transbound. Emerg. Dis. 2020, 67, 1671–1676. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, G.; Alvarez, I.; Fondevila, N.; Politzki, R.; Lomónaco, M.; Rodriguez, S.; Santos, M.D.; Trono, K. Detection of bovine leukemia virus specific antibodies using recombinant p24-ELISA. Vet. Microbiol. 2009, 137, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Kuczewski, A.; Orsel, K.; Barkema, H.W.; Kelton, D.F.; Hutchins, W.A.; van der Meer, F.J. Evaluation of 5 different ELISA for the detection of bovine leukemia virus antibodies. J. Dairy Sci. 2018, 101, 2433–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.Y.; Liu, R.B.; Buehring, G.C. Relative sensitivity and specificity of agar gel immunodiffusion, enzyme immunosorbent assay, and immunoblotting for detection of anti-bovine leukemia virus antibodies in cattle. J. Virol. Methods 2002, 104, 33–39. [Google Scholar] [CrossRef]
- De Giuseppe, A.; Feliziani, F.; Rutili, D.; De Mia, G.M. Expression of the bovine leukemia virus envelope glycoprotein (gp51) by recombinant baculovirus and its use in an enzyme-linked immunosorbent assay. Clin. Diagn. Lab. Immunol. 2004, 11, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.; Yokoyama, K.; Watanuki, S.; Ishizaki, H.; Takeshima, S.-N.; Aida, Y. Development of a new recombinant p24 ELISA system for diagnosis of bovine leukemia virus in serum and milk. Arch. Virol. 2019, 164, 201–211. [Google Scholar] [CrossRef]
- Kobayashi, S.; Hidano, A.; Tsutsui, T.; Yamamoto, T.; Hayama, Y.; Nishida, T.; Muroga, N.; Konishi, M.; Kameyama, K.; Murakami, K. Analysis of risk factors associated with bovine leukemia virus seropositivity within dairy and beef breeding farms in Japan: A nationwide survey. Res. Vet. Sci. 2014, 96, 47–53. [Google Scholar] [CrossRef]
- Endoh, D.; Mizutani, T.; Kirisawa, R.; Maki, Y.; Saito, H.; Kon, Y.; Morikawa, S.; Hayashi, M. Species-independent detection of RNA virus by representational difference analysis using non-ribosomal hexanucleotides for reverse transcription. Nucleic Acids Res. 2005, 33, e65. [Google Scholar] [CrossRef]
- Petersen, M.; Alvarez, I.; Trono, K.; Jaworski, J. Quantification of bovine leukemia virus proviral DNA using a low-cost real-time polymerase chain reaction. J. Dairy Sci. 2018, 101, 6366–6374. [Google Scholar] [CrossRef] [Green Version]
- Lojkić, I.; Balić, D.; Rudan, N.; Kovačić, M.; Čač, Ž.; Periškić, M.; Bedeković, T.; Roić, B.; Ciglar Grozdanić, I. Eradication of bovine leukosis virus on a diary farm through improved virus detection. Vet. Arh. 2013, 83, 581–591. [Google Scholar]
- Klintevall, K.; Ballagi-Pordany, A.; Näslund, K.; Belak, S. Bovine leukaemia virus: Rapid detection of proviral DNA by nested PCR in blood and organs of experimentally infected calves. Vet. Microbiol. 1994, 42, 191–204. [Google Scholar] [CrossRef]
- Molloy, J.B.; Dimmock, C.K.; Eaves, F.W.; Bruyeres, A.G.; Cowley, J.A.; Ward, W.H. Control of bovine leukaemia virus transmission by selective culling of infected cattle on the basis of viral antigen expression in lymphocyte cultures. Vet. Microbiol. 1994, 39, 323–333. [Google Scholar] [CrossRef]
- Marawan, M.A. Molecular Studies on Bovine Leukemia Virus. Ph.D. Thesis, Faculty of Veterinary Medicine, Benha University, Banha, Egypt, 2017. [Google Scholar]
- Selim, A.; El-Haig, M.; Galila, E.S.; Gaede, W. Direct detection of Mycobacterium avium subsp. Paratuberculosis in bovine milk by multiplex Real-time PCR. Anim. Sci. Pap. Rep. 2013, 31, 291–302. [Google Scholar] [CrossRef]
- Elhaig, M.M.; Selim, A.; Mandour, A.S.; Schulz, C.; Hoffmann, B. Prevalence and molecular characterization of peste des petits ruminants virus from Ismailia and Suez, Northeastern Egypt, 2014–2016. Small Rumin. Res. 2018, 169, 94–98. [Google Scholar] [CrossRef]
- Selim, A.; Attia, K.; Ramadan, E.; Hafez, Y.M.; Salman, A. Seroprevalence and molecular characterization of Brucella species in naturally infected cattle and sheep. Prev. Vet. Med. 2019, 171, 104756. [Google Scholar] [CrossRef]
- Selim, A.; Almohammed, H.; Abdelhady, A.; Alouffi, A.; Alshammari, F.A. Molecular detection and risk factors for Anaplasma platys infection in dogs from Egypt. Parasites Vectors 2021, 14, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Takeshima, S.N.; Kitamura-Muramatsu, Y.; Yuan, Y.; Polat, M.; Saito, S.; Aida, Y. BLV-CoCoMo-qPCR-2: Improvements to the BLV-CoCoMo-qPCR assay for bovine leukemia virus by reducing primer degeneracy and constructing an optimal standard curve. Arch. Virol. 2015, 160, 1325–1332. [Google Scholar] [CrossRef]
- Panei, C.J.; Takeshima, S.-N.; Omori, T.; Nunoya, T.; Davis, W.C.; Ishizaki, H.; Matoba, K.; Aida, Y. Estimation of bovine leukemia virus (BLV) proviral load harbored by lymphocyte subpopulations in BLV-infected cattle at the subclinical stage of enzootic bovine leucosis using BLV-CoCoMo-qPCR. BMC Vet. Res. 2013, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Watanuki, S.; Murakami, H.; Sato, R.; Ishizaki, H.; Aida, Y. Development of a luminescence syncytium induction assay (LuSIA) for easily detecting and quantitatively measuring bovine leukemia virus infection. Arch. Virol. 2018, 163, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Watanuki, S.; Bai, L.; Borjigin, L.; Ishizaki, H.; Matsumoto, Y.; Hachiya, Y.; Sentsui, H.; Aida, Y. A sensitive luminescence syncytium induction assay (LuSIA) based on a reporter plasmid containing a mutation in the glucocorticoid response element in the long terminal repeat U3 region of bovine leukemia virus. Virol. J. 2019, 16, 66. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Bai, L.; Borjigin, L.; Aida, Y. Overexpression of bovine leukemia virus receptor SLC7A1/CAT1 enhances cellular susceptibility to BLV infection on luminescence syncytium induction assay (LuSIA). Virol. J. 2020, 17, 1–6. [Google Scholar] [CrossRef]
- Suzuki, T.; Ikeda, H.; Mase, M. Restricted viral cDNA synthesis in cell lines that fail to support productive infection by bovine leukemia virus. Arch. Virol. 2018, 163, 2415–2422. [Google Scholar] [CrossRef] [PubMed]
- OIE. Enzootic Bovine Leucosis, OIE Manual of Standers for Diagnostic Tests and Vaccines; OIE: Paris, France, 2000; pp. 371–380. Available online: https://pubag.nal.usda.gov/catalog/36672 (accessed on 23 October 2021).
- Bai, L.; Hirose, T.; Assi, W.; Wada, S.; Takeshima, S.-N.; Aida, Y. Bovine Leukemia Virus Infection Affects Host Gene Expression Associated with DNA Mismatch Repair. Pathogens 2020, 9, 909. [Google Scholar] [CrossRef] [PubMed]
- Souza, F.N.; Monteiro, A.M.; dos Santos, P.R.; Sanchez, E.M.R.; Blagitz, M.G.; Latorre, A.O.; Neto, A.M.F.; Gidlund, M.; Della Libera, A.M. Antioxidant status and biomarkers of oxidative stress in bovine leukemia virus-infected dairy cows. Vet. Immunol. Immunopathol. 2011, 143, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Krasnikova, E.S.; Krasnikov, A.V.; Annikov, V.V.; Rykhlov, A.S.; Galiullin, A.K.; Alimov, A.M. Analysis of hemo-biochemical status of cows infected with retroviruse. Res. J. Pharm. Biol. Chem. Sci. 2018, 9, 1122–1128. [Google Scholar]
- Sandev, N.; Zapryanova, D.; Stoycheva, I.; Rusenova, N.; Mircheva, T. Investigation of some hematological and blood biochemical parameters in cattle spontaneously infected with bovine leukosis virus. Maced. Vet. Rev. 2013, 36, 107–110. [Google Scholar]
- Ali, A.-F.; Selim, A.; Manaa, E.A.; Abdelrahman, A.; Sakr, A. Oxidative state markers and clinicopathological findings associated with bovine leukemia virus infection in cattle. Microb. Pathog. 2019, 136, 103662. [Google Scholar] [CrossRef]
- Notsu, K.; Hashida, S.; Mitoma, S.; Kubo, M.; Arikawa, G.; Agah, M.A.; El-Khaiat, H.M.; Mai, T.N.; Nguyen, T.H.; Elhanafy, E. Assessment of Hematological Parameters and Carcass Weight in Bovine Leukemia Virus Infection in Slaughtered Beef Cattle. J. Vet. Epidemiol. 2018, 22, 43–48. [Google Scholar] [CrossRef]
- Ruiz, V.; Porta, N.G.; Lomónaco, M.; Trono, K.; Alvarez, I. Bovine leukemia virus infection in neonatal calves. Risk factors and control measures. Front. Vet. Sci. 2018, 5, 267. [Google Scholar] [CrossRef] [Green Version]
- Nishiike, M.; Haoka, M.; Doi, T.; Kohda, T.; Mukamoto, M. Development of a preliminary diagnostic measure for bovine leukosis in dairy cows using peripheral white blood cell and lymphocyte counts. J. Vet. Med. Sci. 2016, 78, 1145–1151. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, H.C.; Norby, B.; Droscha, C.J.; Sordillo, L.M.; Coussens, P.M.; Bartlett, P.C. Bovine leukemia virus detection and dynamics following experimental inoculation. Res. Vet. Sci. 2020, 133, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Mekata, H.; Sekiguchi, S.; Kirino, Y.; Mitoma, S.; Honkawa, K.; Horii, Y.; Norimine, J. Cattle with the BoLA class II DRB3* 0902 allele have significantly lower bovine leukemia proviral loads. J. Vet. Med. Sci. 2017, 79, 1552–1555. [Google Scholar] [CrossRef] [Green Version]
- Notsu, K.; Wiratsudakul, A.; Mitoma, S.; El Daous, H.; Kaneko, C.; El-Khaiat, H.M.; Norimine, J.; Sekiguchi, S. Quantitative Risk Assessment for the Introduction of Bovine Leukemia Virus-Infected Cattle Using a Cattle Movement Network Analysis. Pathogens 2020, 9, 903. [Google Scholar] [CrossRef]
- Bartlett, P.C.; Ruggiero, V.J.; Hutchinson, H.C.; Droscha, C.J.; Norby, B.; Sporer, K.R.; Taxis, T.M. Current Developments in the Epidemiology and Control of Enzootic Bovine Leukosis as Caused by Bovine Leukemia Virus. Pathogens 2020, 9, 1058. [Google Scholar] [CrossRef] [PubMed]
- Kanno, T.; Ishihara, R.; Hatama, S.; Oue, Y.; Edamatsu, H.; Konno, Y.; Tachibana, S.; Murakami, K. Effect of freezing treatment on colostrum to prevent the transmission of bovine leukemia virus. J. Vet. Med. Sci. 2014, 76, 255–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsuki, H.; Takeshima, S.-n.; Aida, Y. Generation of virus-like particle as a vaccine strategy against bovine leukemia virus. Retrovirology 2015, 12, P47. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.; Sato, H.; Kubo, Y.; Wada, S.; Aida, Y. CAT1/SLC7A1 acts as a cellular receptor for bovine leukemia virus infection. FASEB J. 2019, 33, 14516–14527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, L.; Takeshima, S.-N.; Isogai, E.; Kohara, J.; Aida, Y. Novel CD8+ cytotoxic T cell epitopes in bovine leukemia virus with cattle. Vaccine 2015, 33, 7194–7202. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, A. Development and Characterisation of Recombinant LSDV-Vectored Dual Vaccines against Bovine Leukaemia Virus and Lumpy Skin Disease Virus. Master’s Thesis, University of Cape Town, Cape Town, South Africa, 2019. [Google Scholar]
- Abdala, A.; Alvarez, I.; Brossel, H.; Calvinho, L.; Carignano, H.; Franco, L.; Gazon, H.; Gillissen, C.; Hamaidia, M.; Hoyos, C. BLV: Lessons on vaccine development. Retrovirology 2019, 16, 26. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Continent | Countries | Genotype | References |
|---|---|---|---|
| Europe | France | 4 | [44,50,53,58,59,60,61,62,63,64] |
| Belgium | 4 | ||
| Moldova | 7 | ||
| Asia | Korea | 1&3 | |
| Japan | 1&2&3 | ||
| Russia | 4&7&8 | ||
| Thailand | 1&6&10 | ||
| Myanmar | 1&6&10 | ||
| China | 4&6&11 | ||
| Philippines | 1&6 | ||
| Iran | 4 | ||
| Australia | Australia | 1 | |
| North America | USA | 1&3 | |
| Central America | Costa Rica | 1&5 | |
| South America | Brazil | 1&2&6 | |
| Uruguay | 1 | ||
| Paraguay | 1&6 | ||
| Bolivia | 9 | ||
| Argentine | 1&2&4&6 | ||
| Peru | 2 | ||
| Colombia | 1&3&6 | ||
| Africa | Egypt | 1&4 | |
| South Africa | 1&4 | ||
| Zambia | 1 |
| Status | Continent | Countries | Year | References |
|---|---|---|---|---|
| BLV free countries | Europe | Andorra | 1994 | [85] |
| Cyprus | 1995 | [85] | ||
| Czech Republic | 2010 | [85] | ||
| Finland | 2008 | [85] | ||
| Ireland | 1999 | [85] | ||
| Norway | 2002 | [85] | ||
| Spain | 1994 | [85] | ||
| UK | 1996 | [85] | ||
| The Netherlands | 2009 | [86] | ||
| Sweden | 2007 | [85] | ||
| Denmark | 1990 | [85] | ||
| Estonia | 2013 | [85] | ||
| Switzerland | 2005 | [85] | ||
| Slovenia | 2006 | [85] | ||
| Oceania | Australia | 2013 | [12] | |
| New Zealand | 2008 | [87] | ||
| Tunisia | 2005 | [85] | ||
| Asia | Kyrgyzstan | 2008 | [85] | |
| Kazakhstan | 2007 | [85] | ||
| BLV existing countries with unknown prevalence | Europe | Croatia | Present | [86,88] |
| Ukraine | [89] | |||
| Italy | [85] | |||
| Portugal | [85] | |||
| Belarus | [86,89] | |||
| Greece | [85] | |||
| Bulgaria | [85] | |||
| Latvia | [85] | |||
| South America | Uruguay | [90] | ||
| BLV existing countries with variable prevalence | North America | USA (Dairy 83.9%, Beef 39%) | 2007 | [91] |
| Mexico (Dairy 36.1%, Beef 4%) | 1983 | [92] | ||
| Canada (78% at herd level) | 1998–2003 | [5] | ||
| South America | Chile (southern regions) (27.9% individual level) | 2009 | [48] | |
| Brazil 17.1% 60.8% | 1980–1989 1992–1995 | [93,94] | ||
| Argentina (Buenos Aires) (77.4% at an individual, 90.9% at herd level) (Multiple regions) (32.85% at individual, 84% at herd level) | 2007 1998–1999 | [48] [95] | ||
| Peru (Multiple regions) (31% at the individual level, 42.3% individual level) | 1983 2008 | [96] [48] | ||
| Bolivia (Multiple regions) 30.7% individual level | 2008 | [48] | ||
| Venezuela 33.3% individual level | 1978 | [97] | ||
| Paragua (Asuncion) 54.7% individual level | 2008 | [48] | ||
| Colombia 62% individual level | 2020 | [98] | ||
| Asia | China (Dairy 49.1%, Beef 1.6%) | 2013–2014 | [99] | |
| Taiwan (81.8% at animal level and 99.1% at herd level) | 2019 | [100] | ||
| Cambodia Draught cattle 5.3% | 2000 | [101] | ||
| Japan (Nationwide) Dairy 49.1% Beef 1.6% 79.1% of the dairy herd 73.3% at individual | 2009–2011 2007 2012–2014 | [102] [103] [104] | ||
| Mongolia (Dairy 3.9%) | 2014 | [105] | ||
| Iran (nationwide) (22.1% to 25.4%) | 2012–2014 | [5] | ||
| Philippines (4.8% to 9.7%) | 2010–2012 | [106] | ||
| Myanmar (9.1% at individual) | 2016 | [48] | ||
| Thailand (58.7% at individual) | 2013–2014 | [53] | ||
| Pakistan (20% of dairy) | 2019 | [107] | ||
| Middle East | Saudi Arabia (20.2% of dairy) | 1990 | [108] | |
| Turkey (48.3% dairy) | [109] | |||
| Israel (5% at individual) | [110] | |||
| Iraq (7% of dairy) | 2015 | [111] | ||
| Egypt (17.7% of dairy) | 2020 | [112] |
| Rewarded Samples | Test | Advantages | Disadvantages | References |
|---|---|---|---|---|
| Serum sample | 1. ELISA (Antibodies p24, gp51) | Sensitive, specific, large scale screening and rapid | False negatives (cattle in the early infection phase) False-positive (maternally derived antibodies) Cannot evaluate disease states of infected cattle. | [157,170,171,172,173] |
| 2. RIA (Antibodies p24) | Sensitive Able to detect BLV during the early period of infection | Cannot be used for mass screening | [174,175] | |
| 3. AGIDT (Antibodies p24, gp51) | Specific, simple, rapid, screening and Less expensive | Less sensitive Inconclusive Fail to evaluate disease states | [9,171,172,173] | |
| Milk and Bulk milk sample | ELISA [157,170,171,172,173] | |||
| Virus particle | PHA (BLV glycoprotein) |
|
| [176] |
| Rewarded Samples | Test | Advantages | Disadvantages | References |
|---|---|---|---|---|
| (Blood, PBMC, Tumour sample, Buffy coat, Milk, somatic cells, Semen, Saliva and Nasal secretions). | Realtime PCR |
|
| [170,177,178,179] |
| Conventional PCR (Single, Semi-Nested, and Nested PCR) |
|
| [157,171,173,180,181] | |
| Blood only | Direct blood-based PCR |
|
| [11,182,183] |
| Blood only | Direct filter PCR |
|
| [184] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marawan, M.A.; Alouffi, A.; El Tokhy, S.; Badawy, S.; Shirani, I.; Dawood, A.; Guo, A.; Almutairi, M.M.; Alshammari, F.A.; Selim, A. Bovine Leukaemia Virus: Current Epidemiological Circumstance and Future Prospective. Viruses 2021, 13, 2167. https://doi.org/10.3390/v13112167
Marawan MA, Alouffi A, El Tokhy S, Badawy S, Shirani I, Dawood A, Guo A, Almutairi MM, Alshammari FA, Selim A. Bovine Leukaemia Virus: Current Epidemiological Circumstance and Future Prospective. Viruses. 2021; 13(11):2167. https://doi.org/10.3390/v13112167
Chicago/Turabian StyleMarawan, Marawan A., Abdulaziz Alouffi, Suleiman El Tokhy, Sara Badawy, Ihsanullah Shirani, Ali Dawood, Aizhen Guo, Mashal M. Almutairi, Fahdah Ayed Alshammari, and Abdelfattah Selim. 2021. "Bovine Leukaemia Virus: Current Epidemiological Circumstance and Future Prospective" Viruses 13, no. 11: 2167. https://doi.org/10.3390/v13112167
APA StyleMarawan, M. A., Alouffi, A., El Tokhy, S., Badawy, S., Shirani, I., Dawood, A., Guo, A., Almutairi, M. M., Alshammari, F. A., & Selim, A. (2021). Bovine Leukaemia Virus: Current Epidemiological Circumstance and Future Prospective. Viruses, 13(11), 2167. https://doi.org/10.3390/v13112167