
Susceptibility of Four Abalone Species, Haliotis gigantea, Haliotis discus discus, Haliotis discus hannai and Haliotis diversicolor, to Abalone asfa-like Virus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Abalones
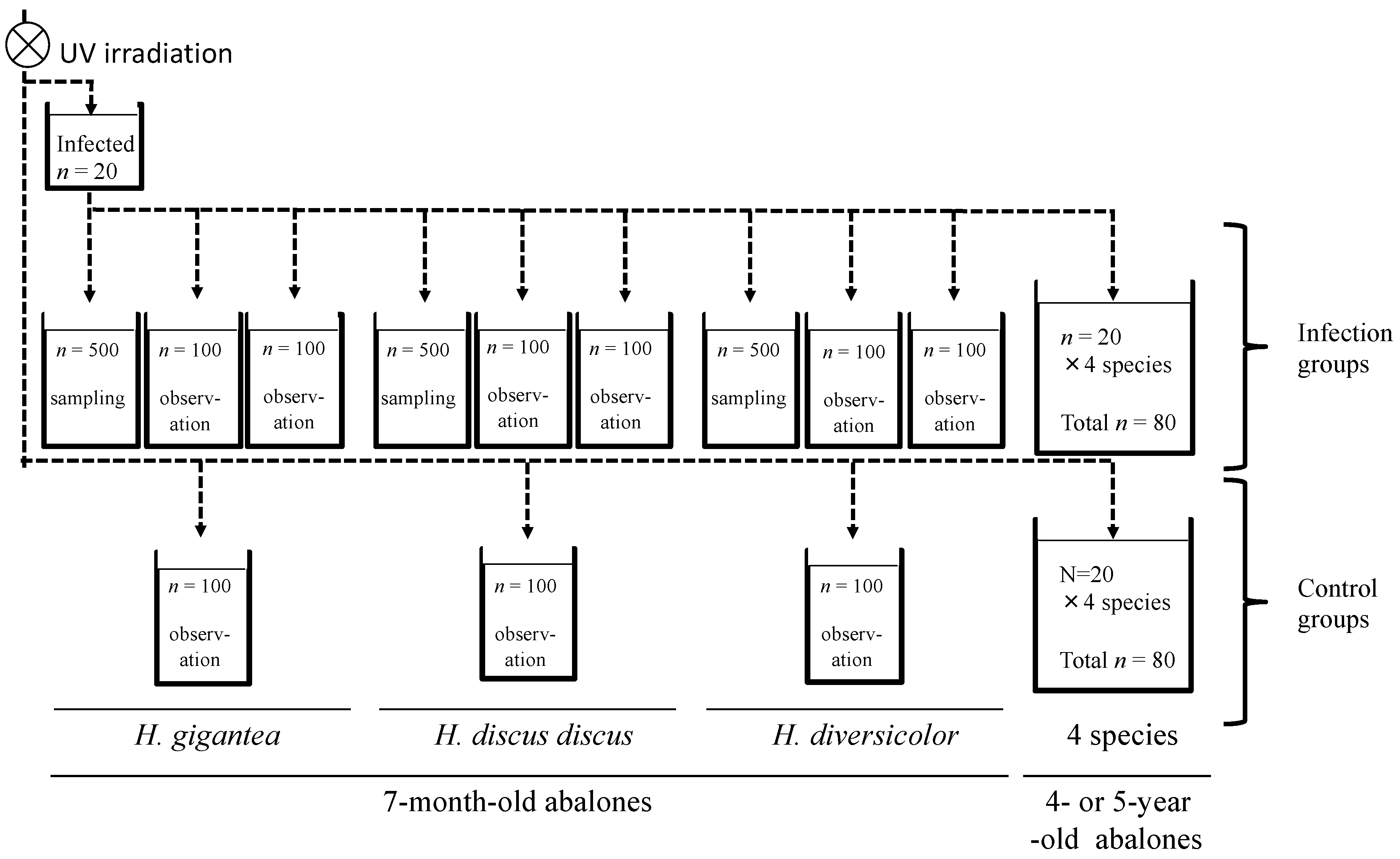
2.3. Infection Experiment 1: Cohabitation Infection with Four Species at Different Ages
2.4. Infection Experiment 2: Injection Infection of H. discus discus and H. diversicolor at 12 Months of Age
2.5. Generation of Mouse Antisera against AbALV MCP and Piscine orthoreovirus 2 (PRV-2) σ-1 Recombinant Proteins
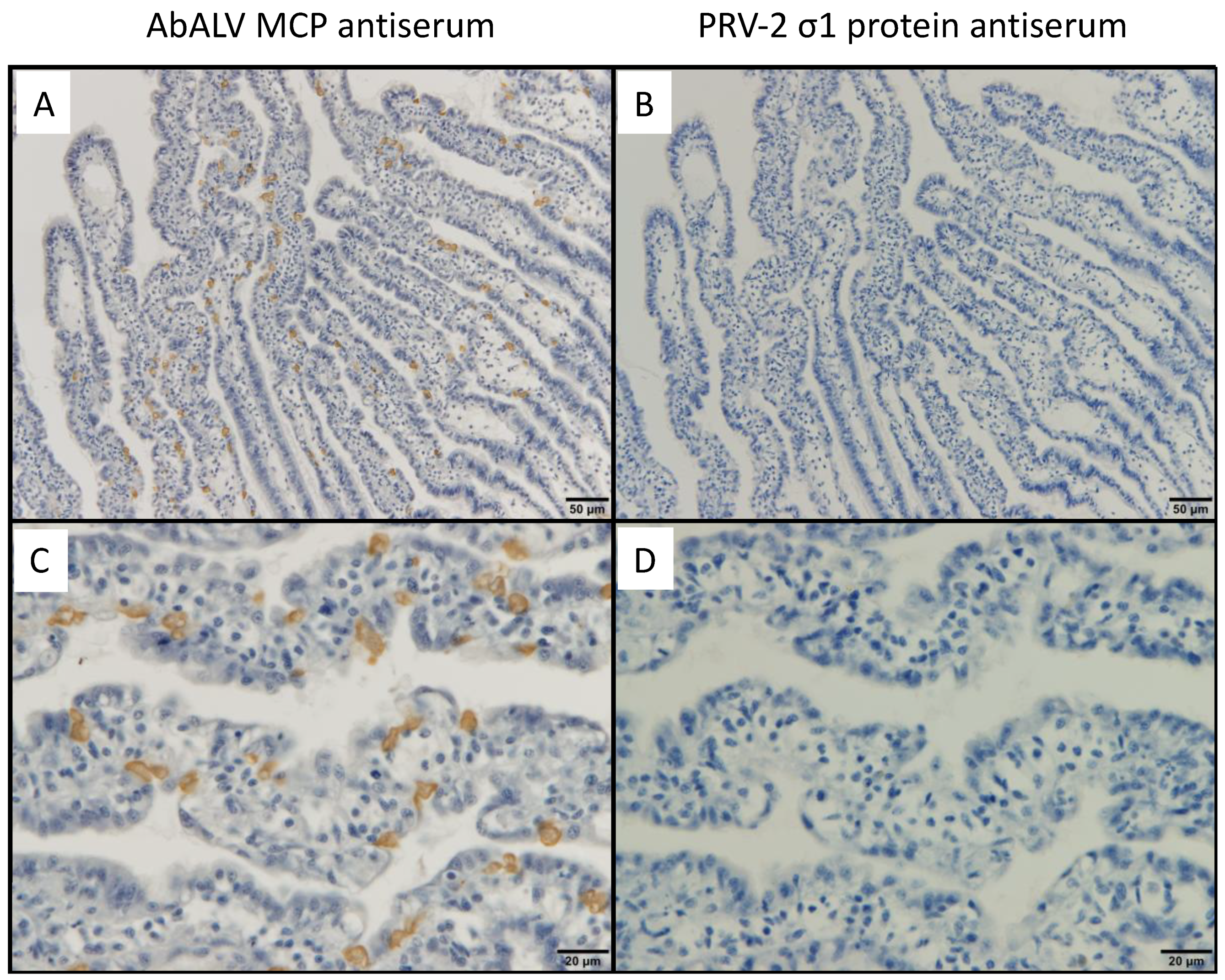
2.6. Immunohistochemistry
2.7. Quantitative PCR
2.8. Statistical Analysis
3. Results
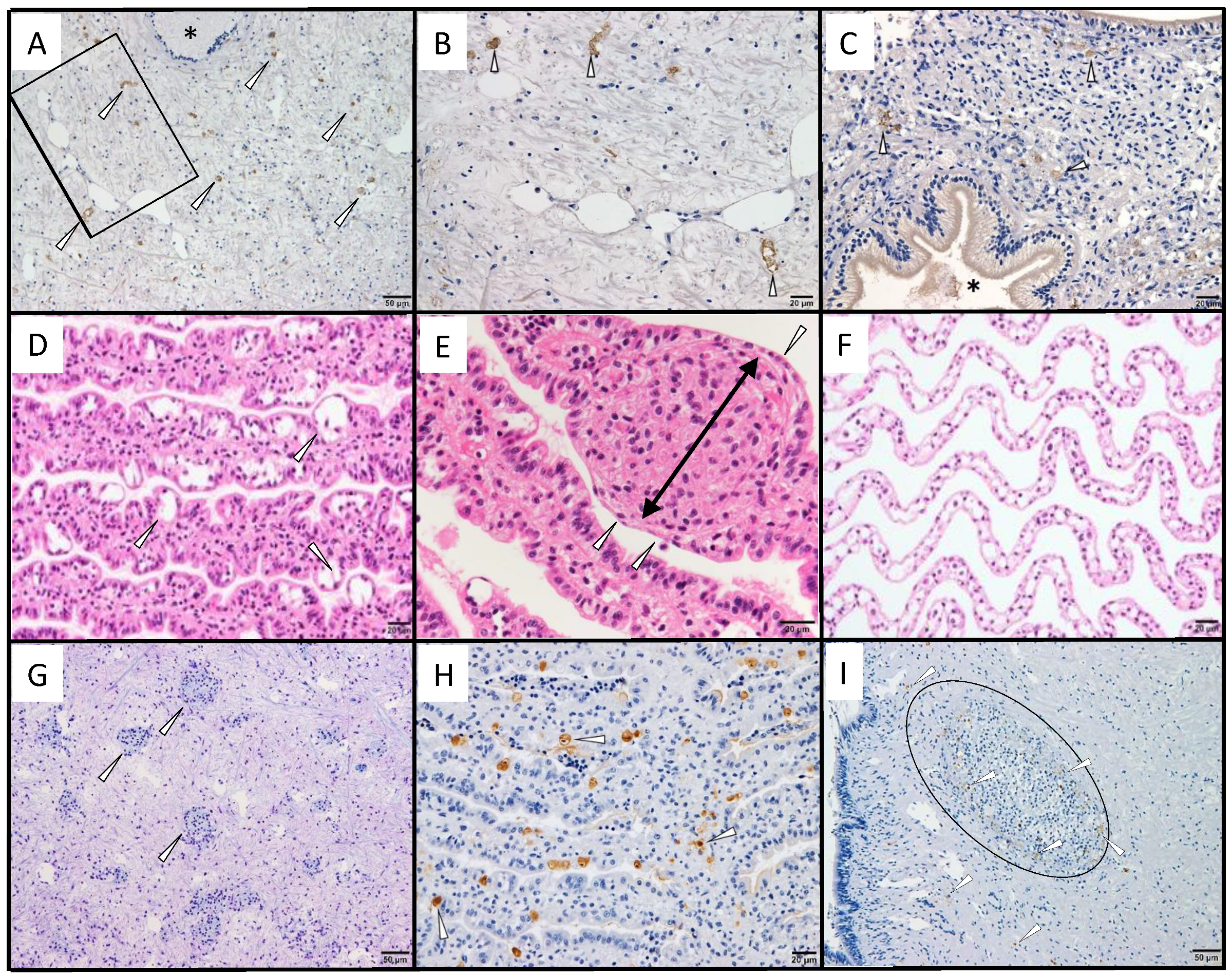
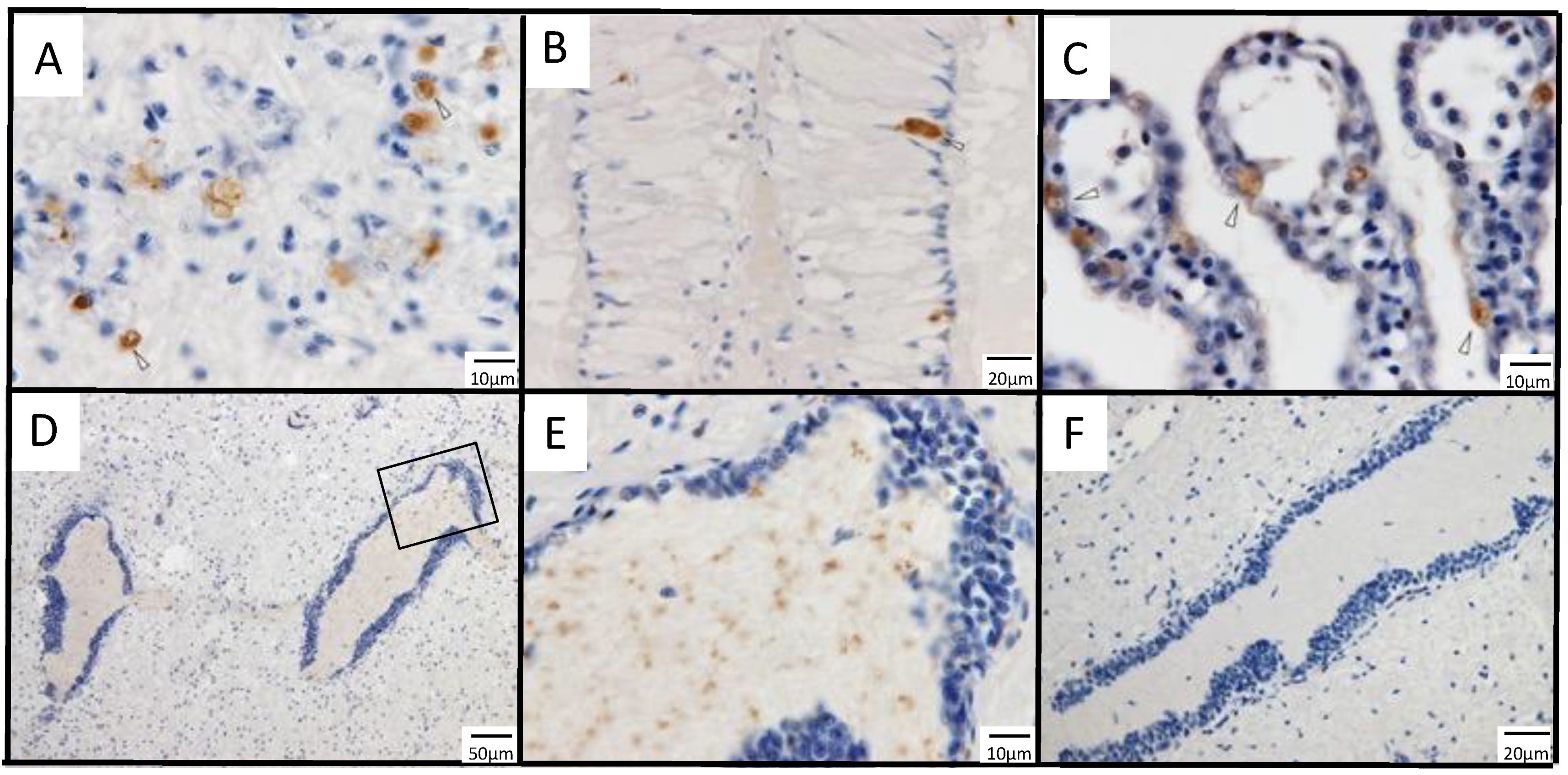
3.1. Immunohistochemistry for a Previously Reported Diseased Abalone and Spontaneously Infected Abalone
3.2. Infection Experiment 1: Cohabitation Infection with Four Species at Different Ages
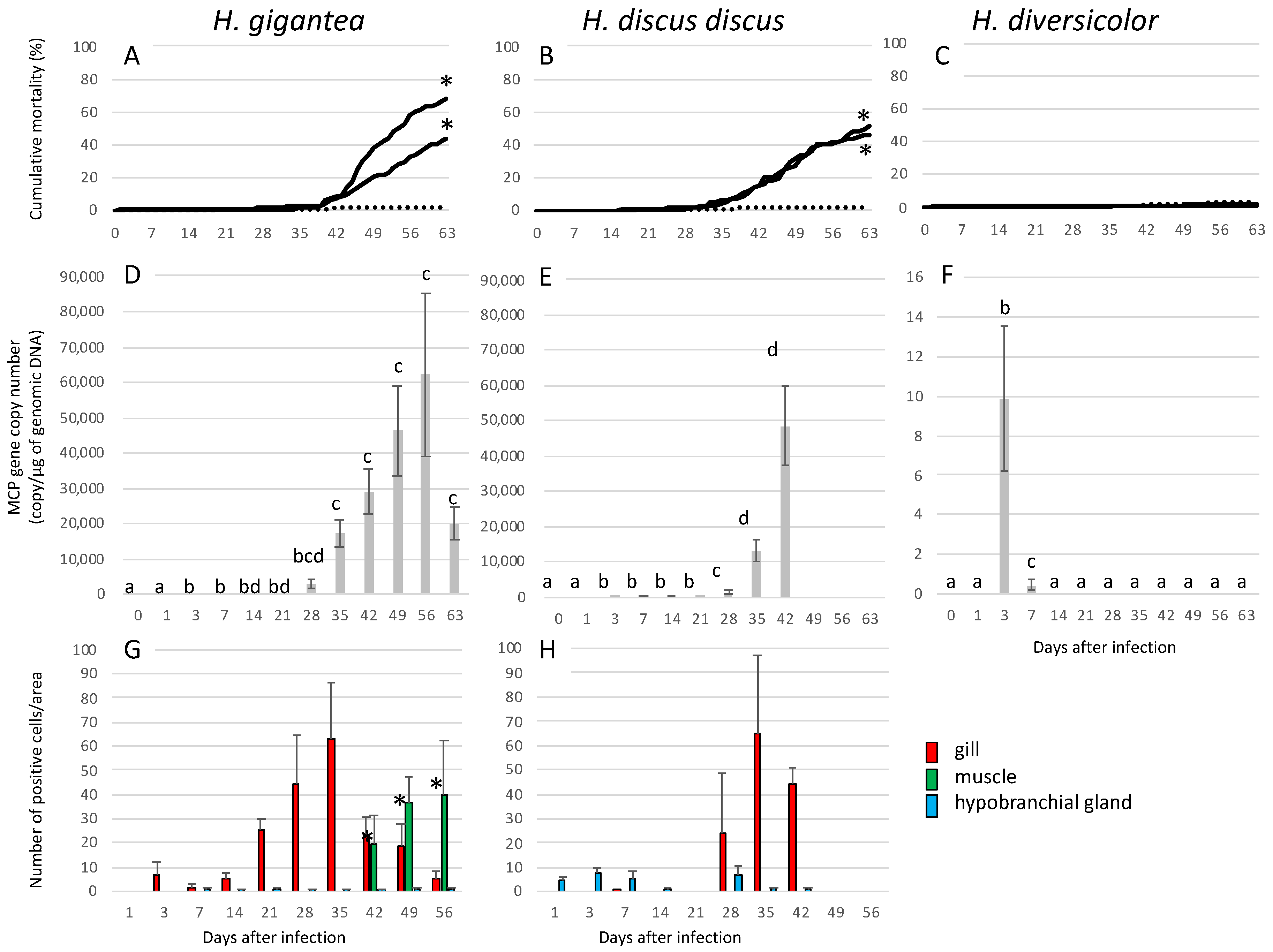
3.2.1. Infection Experiment 1: Mortality and Appearance
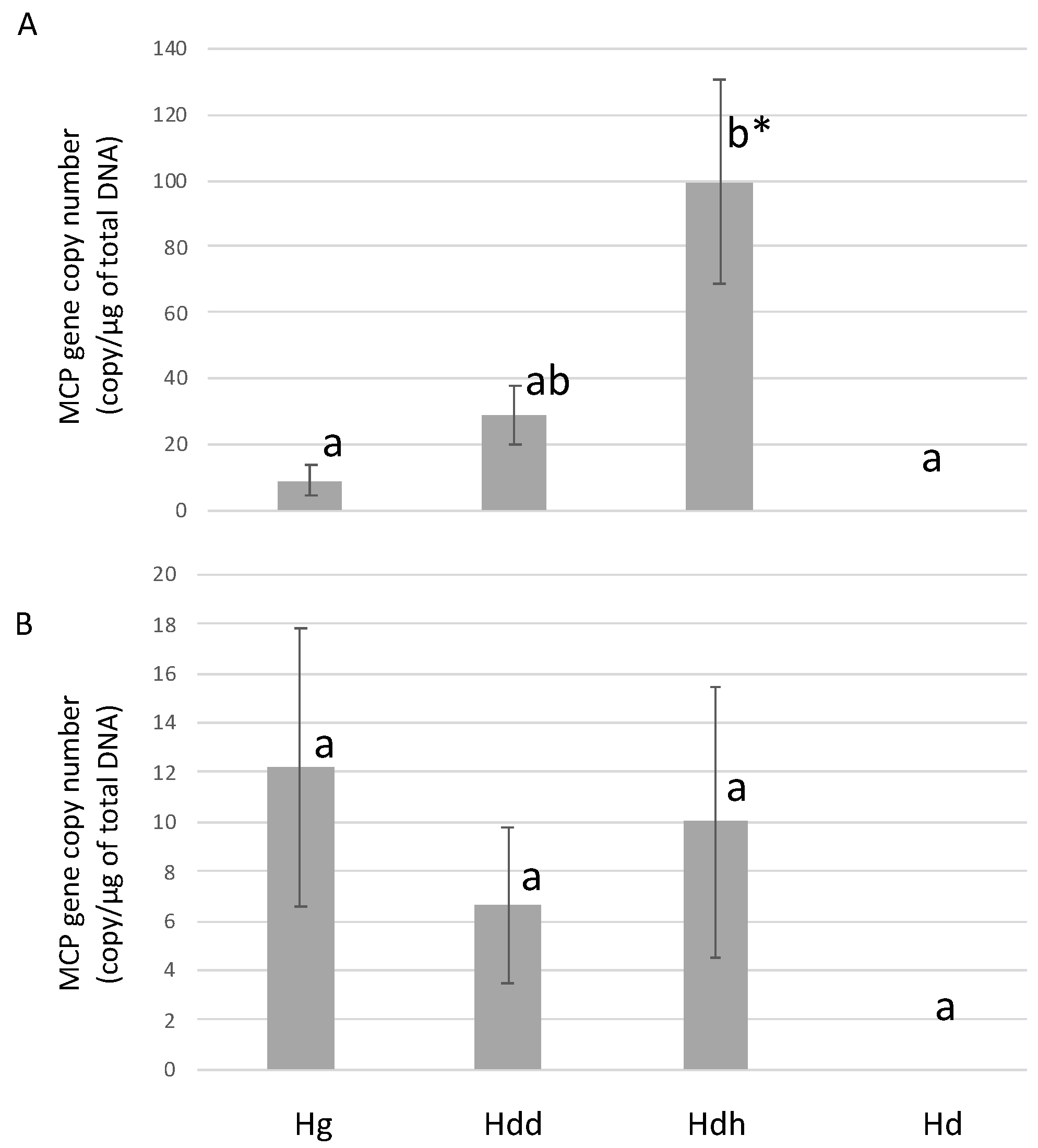
3.2.2. Infection Experiment 1: Viral Load
3.2.3. Infection Experiment 1: Immunohistochemistry
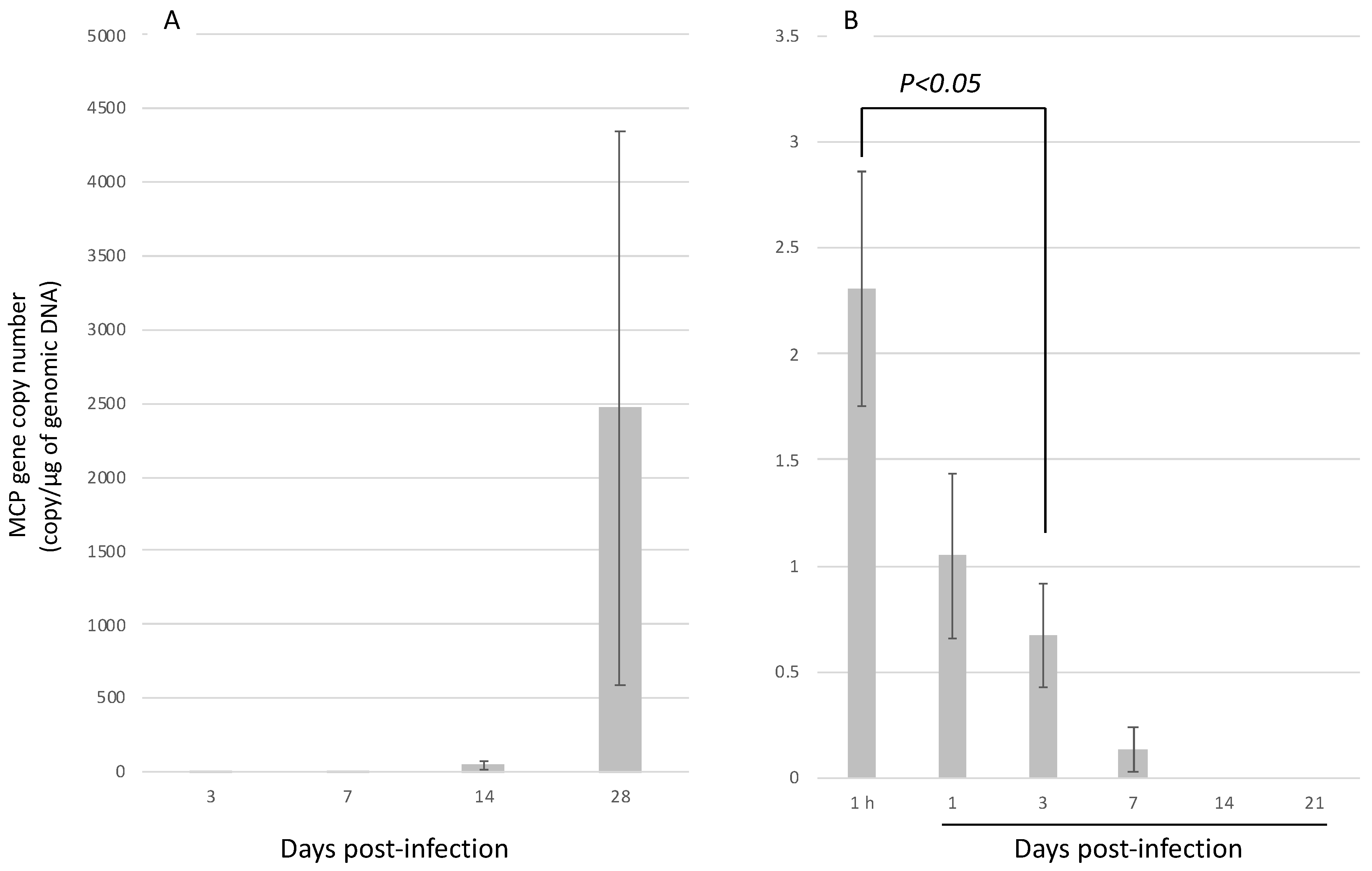
3.3. Infection Experiment 2: Injection Infection with H. discus discus and H. diversicolor at 12 Months of Age
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Prince, J.D. The decline of global abalone (genus Haliotis) production in the late twentieth century: Is there a future? In Stock Enhancement and Sea Ranching, 2nd ed.; Leber, K.M., Kitada, S., Svasand, T., Blankenship, H.L., Eds.; Blackwell Publishing: Oxford, UK, 2004; pp. 427–443. [Google Scholar]
- Hamasaki, K.; Kitada, S. The enhancement of abalone stocks: Lessons from Japanese case studies. Fish Fish. 2008, 9, 243–260. [Google Scholar] [CrossRef]
- Nishimori, E.; Hasegawa, O.; Numata, T.; Wakabayashi, H. Vibrio carchariae causes mass mortalities in Japanese abalone, Sulculus diversicolor supratexta. Fish Pathol. 1998, 33, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Sawabe, T.; Inoue, S.; Fukui, Y.; Yoshie, K.; Nishihara, Y.; Miura, H. Mass mortality of Japanese abalone Haliotis discus hannai caused by Vibrio harveyi infection. Microbes Environ. 2007, 22, 300–308. [Google Scholar] [CrossRef] [Green Version]
- Kamaishi, T.; Miwa, S.; Goto, E.; Matsuyama, T.; Oseko, N. Mass mortality of giant abalone Haliotis gigantea caused by a Francisella sp. bacterium. Dis. Aquat. Org. 2010, 89, 145–154. [Google Scholar] [CrossRef]
- Brevik, Ø.; Ottem, K.; Kamaishi, T.; Watanabe, K.; Nylund, A. Francisella halioticida sp. nov., a pathogen of farmed giant abalone (Haliotis gigantea) in Japan. J. Appl. Microbiol. 2011, 111, 1044–1056. [Google Scholar] [CrossRef]
- Kiryu, I.; Kurita, J.; Yuasa, K.; Nishioka, T.; Shimahara, Y.; Kamaishi, T.; Ototake, M.; Oseko, N.; Tange, N.; Inoue, M. First Detection of Candidatus Xenohaliotis Californiensis, the Causative Agent of Withering Syndrome, in Japanese Black Abalone Haliotis discus discus in Japan. Fish Pathol. 2013, 48, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Nakatsugawa, T.; Hatai, K.; Kubota, S.S. Histopathological findings on cultured juvenile abalone, Nordotis discus, with muscular atrophy. Fish Pathol. 1988, 23, 203–204. [Google Scholar] [CrossRef] [Green Version]
- Nakatsugawa, T. Infectious nature of a disease in cultured juvenile abalone with muscular atrophy. Fish Pathol. 1990, 25, 207–211. [Google Scholar] [CrossRef]
- Momoyama, K.; Nakatsugawa, T.; Yurano, N. Mass mortalities of juvenile abalones, Haliotis spp., caused by amyotrophia. Fish Pathol. 1999, 34, 7–14. [Google Scholar] [CrossRef]
- Hooper, C.; Hardy-Smith, P.; Handlinger, J. Ganglioneuritis causing high mortalities in farmed Australian abalone (Haliotis laevigata and Haliotis rubra). Aust. Vet. J. 2007, 85, 188–193. [Google Scholar] [CrossRef]
- Zhuang, J.; Cai, G.; Lin, Q.; Wu, Z.; Xie, L. A bacteriophage-related chimeric marine virus infecting abalone. PLoS ONE 2010, 5, e13850. [Google Scholar] [CrossRef] [Green Version]
- Nakatsugawa, T.; Momoyama, K. Susceptibility to the agent of amyotrophia among Japanese black abalone of different ages. Fish Pathol. 1999, 34, 215–216. [Google Scholar] [CrossRef]
- Nakatsugawa, T. Amyotrophia of Japanese black abalone, Nordotis discus discus. Fish Pathol. 1999, 34, 221. [Google Scholar] [CrossRef]
- Momoyama, K. Experiments for characterizing the causative agent of amyotrophia in juvenile abalones Haliotis spp. Fish Pathol. 2000, 35, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Otsu, R.; Sasaki, K. Virus-like particles detected from juvenile abalones (Nordotis discus discus) reared with an epizootic fatal wasting disease. J. Invertebr. Pathol. 1997, 70, 167–168. [Google Scholar] [CrossRef] [PubMed]
- Nakatsugawa, T.; Nagai, T.; Hiya, K.; Nishizawa, T.; Muroga, K. A virus isolated from juvenile Japanese black abalone Nordotis discus discus affected with amyotrophia. Dis. Aquat. Org. 1999, 36, 159–161. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, T.; Takano, T.; Nishiki, I.; Fujiwara, A.; Kiryu, I.; Inada, M.; Sakai, T.; Terashima, S.; Matsuura, Y.; Isowa, K. A novel Asfarvirus-like virus identified as a potential cause of mass mortality of abalone. Sci. Rep. 2020, 10, 4620. [Google Scholar] [CrossRef]
- Nakatsugawa, T.; Okabe, M.; Muroga, K. Horizontal transmission of amyotrophia in Japanese black abalone. Fish Pathol. 2000, 35, 11–14. [Google Scholar] [CrossRef]
- Shaw, B.L.; Battle, H.I. The gross and microscopic anatomy of the digestive tract of the oyster Crassostrea virginica (Gmelin). Can. J. Zool. 1957, 35, 325–347. [Google Scholar] [CrossRef]
- Takano, T.; Nawata, A.; Sakai, T.; Matsuyama, T.; Ito, T.; Kurita, J.; Terashima, S.; Yasuike, M.; Nakamura, Y.; Fujiwara, A. Full-genome sequencing and confirmation of the causative agent of erythrocytic inclusion body syndrome in coho salmon identifies a new type of piscine orthoreovirus. PLoS ONE 2016, 11, e0165424. [Google Scholar] [CrossRef] [Green Version]
- Estes, J.A.; Lindberg, D.R.; Wray, C. Evolution of large body size in abalones (Haliotis): Patterns and implications. Paleobiology 2005, 31, 591–606. [Google Scholar] [CrossRef] [Green Version]
- Hirase, S.; Yamasaki, Y.Y.; Sekino, M.; Nishisako, M.; Ikeda, M.; Hara, M.; Merilä, J.; Kikuchi, K. Genomic evidence for speciation with gene flow in broadcast spawning marine invertebrates. Mol. Biol. Evol. 2021, 38, 4683–4699. [Google Scholar] [CrossRef]
- Fujii, A. Difference of survival and growth between juveniles of disk abalone, Nordotis discus, and giant ablone, N. gigantea, under rearing conditions. Bull. Nagasaki Prefect. Inst. Fish. 1996, 22, 27–30. [Google Scholar]
- Post, J.; Weesendorp, E.; Montoya, M.; Loeffen, W.L. Influence of age and dose of African swine fever virus infections on clinical outcome and blood parameters in pigs. Viral Immunol. 2017, 30, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Blome, S.; Gabriel, C.; Dietze, K.; Breithaupt, A.; Beer, M. High virulence of African swine fever virus caucasus isolate in European wild boars of all ages. Emerg. Infect. Dis. 2012, 18, 708. [Google Scholar] [CrossRef]
- Thomson, G.; Gainaru, M.; Van Dellen, A. Experimental infection of warthog (Phacochoerus aethiopicus) with African swine fever virus. Onderstepoort J. Vet. Res. 1980, 47, 19–22. [Google Scholar]
- Oura, C.; Powell, P.; Parkhouse, R. Detection of African swine fever virus in infected pig tissues by immunocytochemistry and in situ hybridisation. J. Virol. Methods 1998, 72, 205–217. [Google Scholar] [CrossRef]
- Kleiboeker, S.B.; Scoles, G.A. Pathogenesis of African swine fever virus in Ornithodoros ticks. Anim. Health Res. Rev. 2001, 2, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Nishimura, M.; Kawamura, T. Prevention of amyotrophia in mass production of 0-year-old abalone, Haliotis discus, by quarantine. Aquac. Sci. 2000, 48, 657–663. [Google Scholar]
- Matsuyama, T.; Kiryu, I.; Inada, M.; Nakayasu, C. Verification of sample collection sites for PCR-based detection assay for abalone asfa-like virus. Fish Pathol. 2021, 56, 18–21. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Length (mm) | Body Weight (g) | |||||||
|---|---|---|---|---|---|---|---|---|
| Experiments | Species Scientific Name | Starting Disease Status | Use in Experiments | Age at the Start of the Experiment | Range | Mean | Range | Mean |
| Infection experiment 1 | ||||||||
| H. gigantea | Spontaneously infected * | Infectious source | 8 months | 8–11 | 9.9 ± 0.3 | 0.09–0.18 | 0.15 ± 0.02 | |
| Healthy | Recipient | 7 months | 7–11 | 9.7 ± 0.2 | 0.07–0.17 | 0.11 ± 0.01 | ||
| Healthy | Recipient | 4 years | 50–61 | 54.8 ± 1.1 | 11.0–21.5 | 15.2 ± 1.0 | ||
| H. discus discus | Healthy | Recipient | 7 months | 8–12 | 10.4 ± 0.3 | 0.09–0.27 | 0.16 ± 0.01 | |
| Healthy | Recipient | 4 years | 46–53 | 49.0 ± 0.7 | 10.0–13.9 | 12.8 ± 0.5 | ||
| H. discus hanai | Healthy | Recipient | 4 years | 61–71 | 64.6 ± 0.8 | 21.1–32.8 | 25.6 ± 0.9 | |
| H. diversicolor | Healthy | Recipient | 7 months | 10–17 | 13.9 ± 0.4 | 0.15–0.63 | 0.40 ± 0.03 | |
| Healthy | Recipient | 5 years | 52–58 | 52.8 ± 0.8 | 14.7–30.2 | 20.1 ± 1.2 | ||
| Infection experiment 2 | ||||||||
| H. discus discus | Spontaneously infected † | Infectious source | 13 months | 23–25 | 24.0 ± 0.6 | 1.3–2.0 | 1.7 ± 0.2 | |
| H. discus discus | Healthy | Recipient | 12 months | 22–27 | 23.9 ± 0.5 | 1.1–2.5 | 1.5 ± 0.03 | |
| H. diversicolor | Healthy | Recipient | 12 months | 20–27 | 23.1 ± 0.9 | 1.0–2.4 | 1.5 ± 0.03 | |
| Immunohistochemistry | ||||||||
| H. discus discus | Experimentally infected ‡ | 12 months | No record | No record | No record | No record | ||
| Healthy ‡ | 12 months | No record | No record | No record | No record | |||
| Spontaneously infected § | 8 months | 16–28 | No record | No record | No record | |||
| Healthy | 7 months | 8–12 | 10.4 ± 0.3 | 0.09–0.27 | 0.16 ± 0.01 | |||
| Number of Abalones with Positive Nerve Trunks (Out of 5 Animals) | Number of Abalones with Abnormal Cell Masses (Out of 5 Animals) | |||||
|---|---|---|---|---|---|---|
| Days after Infection | H. gigantea | H. discus discus | H. diversicolor | H. gigantea | H. discus discus | H. diversicolor |
| 1 | 0 | 0 | 0 | 0 | 1 | 0 |
| 3 | 2 | 0 | 0 | 0 | 4 | 0 |
| 7 | 3 | 0 | 0 | 0 | 4 | 0 |
| 14 | 1 | 0 | 0 | 0 | 3 | 0 |
| 21 | 2 | 2 | 0 | 0 | 1 | 0 |
| 28 | 1 | 0 | 0 | 0 | 0 | 0 |
| 35 | 1 | 0 | 0 | 0 | 0 | 0 |
| 42 | 0 | 0 | 0 | 0 | 1 | 0 |
| 49 | 0 | - | 0 | 0 | - | 0 |
| 56 | 0 | - | 0 | 0 | - | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsuyama, T.; Kiryu, I.; Inada, M.; Takano, T.; Matsuura, Y.; Kamaishi, T. Susceptibility of Four Abalone Species, Haliotis gigantea, Haliotis discus discus, Haliotis discus hannai and Haliotis diversicolor, to Abalone asfa-like Virus. Viruses 2021, 13, 2315. https://doi.org/10.3390/v13112315
Matsuyama T, Kiryu I, Inada M, Takano T, Matsuura Y, Kamaishi T. Susceptibility of Four Abalone Species, Haliotis gigantea, Haliotis discus discus, Haliotis discus hannai and Haliotis diversicolor, to Abalone asfa-like Virus. Viruses. 2021; 13(11):2315. https://doi.org/10.3390/v13112315
Chicago/Turabian StyleMatsuyama, Tomomasa, Ikunari Kiryu, Mari Inada, Tomokazu Takano, Yuta Matsuura, and Takashi Kamaishi. 2021. "Susceptibility of Four Abalone Species, Haliotis gigantea, Haliotis discus discus, Haliotis discus hannai and Haliotis diversicolor, to Abalone asfa-like Virus" Viruses 13, no. 11: 2315. https://doi.org/10.3390/v13112315
APA StyleMatsuyama, T., Kiryu, I., Inada, M., Takano, T., Matsuura, Y., & Kamaishi, T. (2021). Susceptibility of Four Abalone Species, Haliotis gigantea, Haliotis discus discus, Haliotis discus hannai and Haliotis diversicolor, to Abalone asfa-like Virus. Viruses, 13(11), 2315. https://doi.org/10.3390/v13112315