
Drug-Screening Strategies for Inhibition of Virus-Induced Neuronal Cell Death


Abstract
:1. Introduction
2. Primary Neurons and Neuronal Cell Lines for Use in Testing Candidate Therapeutics
2.1. Primary Cortical Neuron Isolation
2.1.1. Instruments/Materials for Isolation of Neurons
- Magnifying glass with light source/dissecting scope
- Small scissors
- Two pairs of sharp #5 forceps
- Absorbent pads
- Absorbent wipes
- 15 and 50 mL polypropylene centrifuge tubes
- 10 mL pipets
- 1 mL syringes
- 20 g needles
- Sterile transfer pipettes
- Sterile 100 mL beaker for keeping sterile instruments
- Ice buckets and tray
- Biohazard bags
- 10× PBS pH 7.4
- 12% D-glucose (20×) in PBS (filter sterile)
- 6-, 12- and 24-well amine-coated tissue culture plates
- 0.4% Trypan blue solution
2.1.2. Essential Media and Reagents
- Neurobasal medium 1×, 500 mL
- Aliquot in 50 mL and store at 4 °C in dark
- Neurobasal media with B27 supplement and 0.5 mM L-glutamine
- B-27 serum-free supplement 50×, 10 mL
- Glutamax I (200 mM, in 0.85% NaCl) 100 mL
- Calcium, Magnesium Free (CMF)-Hanks Balanced Salt Solution (HBSS) 10× liquid 500 mL
- KCl (4000 mg/L), KH2PO4 (600 mg/L), NaCl (80,000 mg/L), Na2HPO4.7H2O (900 mg/L), D-glucose (10,000 mg/L)
- HBSS
- HEPES buffer solution 100× (1 M)
- 238.3 g/L of HEPES prepared in distilled water. pH: 7.2–7.5
- Trypsin, 2.5% (10×) 100 mL
- 25 g of trypsin and 8.5 g of NaCl in 100 mL
2.1.3. Solutions or Media to Prepare Just before Use
- Dissection medium (100 mL): CMF-HBSS (Ca2+, Mg2+ free Hank’s Balanced Salt Solution) buffered with 10 mM HEPES, pH 7.310 mL 10× CMF-HBSS1 mL 1 M HEPES (100×)89 mL H2O
- Digestion solution (5 mL)4.5 mL CMF-HBSS with HEPES0.5 mL 2.5% Trypsin
- Add 4.5 mL of CMF-HBSS with HEPES in 15 mL tubes and keep at room temp. Do not add trypsin before use.
- Keep 2.5% trypsin tube at 37 °C water bath.
- Neuronal plating medium (100 mL)90 mL Opti-MEM with L-glutamine5 mL D-glucose 12% (20×) filter sterilized5 mL FBS (5% final conc.)
- Neuronal maintenance medium (100 mL)98 mLNeurobasal medium2 mL B-27 supplement 50× (2% final conc.)250 µL Glutamine 200 mM (0.5 mM final conc.)
2.1.4. Bench Preparation
- Clean the workbench, dissecting microscope and mouse pad thoroughly with 70% ethanol.
- Place instruments in 70% ethanol for several minutes to sterilize. Lightly shake off ethanol from instruments and move them to an empty sterile 100 mL beaker. Let them air dry completely before use.
- Fill ice bucket and kidney tray (for keeping petri dishes ice cold) with ice.
- Prepare 15 mL polypropylene tubes with 4.5 mL of dissection medium and let them sit at room temp.
- Add 30–35 mL of ice-cold sterile dissection medium in petri dishes (100 mm) and let them sit on ice in kidney tray (at least 4 dishes).
- Warm 2.5% trypsin to 37 °C and neuron plating media to room temperature.
2.1.5. Protocol for Isolation and Culturing
- Euthanize a pregnant mouse (14–17-day gestation) by Isoflurane anesthesia and exsanguination (as defined by an accepted Animal Care and Use protocol). Dissect out the entire uterus along with embryos and place in a sterile petri dish containing ice-cold dissection medium. Keep petri dish on ice in a kidney tray.
- Remove the fetuses from the uterus, and place in a fresh petri dish with ice-cold dissection medium. Separate the head from the body by pulling. The tissue must always be kept submerged in the dissection medium.
- Remove the skin from the head and open the skull cap with forceps. Remove the brain and separate from the rest of the tissue. Separate the cortices from the brain and remove the meninges from the cerebral cortices.
- Collect all the cortices in a 15 mL conical centrifuge tube containing 4.5 mL dissection medium (at room temp).
- Add 0.5 mL of 2.5% trypsin and incubate for 15 min in a water bath at 37 °C (invert tube halfway through).
- Gently remove supernatant with a sterile transfer pipet, leaving cortices at the bottom of the tube.
- Bring the volume to 15 mL with fresh dissection medium, invert the tube 3–4 times gently and let it stand for 5 min at room temperature. Repeat this to allow residual trypsin to diffuse from the tissue. Bring the final volume to 4 mL with neuron plating medium.
- Dissociate the cortices by gently pipetting up and down 10 times with a 10 mL pipet. Split into two 15 mL tubes, each with 2 mL of rough homogenate in them. Dissociate rough homogenate by putting through a 27-gauge needle on a 1 mL syringe approximately 10 times.
- Bring the volume to 10 mL with neuron plating medium.
- Count the total number of cells: Add 10 µL of cell suspension and 10 µL of 0.4% filtered trypan blue in a 96-well plate, mix them well and load on a Neubauer chamber slide and count number of live cells in each corner square and take the average number per corner square and calculate total number of cells per mL using the formula.Cells/mL = Average cells counted in corner square × dilution factor × volume conversion factor (104)
- Plate cells on amine-coated plate using neuronal plating medium.
- After 3–4 h, examine the dishes to ensure that most of the cells are attached, then aspirate the entire medium from all the wells and replace with neuronal maintenance medium.
- Thereafter, replace 1/2 of the media with fresh neuronal maintenance media once per week.
2.2. Culture of Neuronal Cell Lines
2.2.1. Mouse Neuroblastoma Cell Lines
- i
- Neuro-2a
- ii
- C17.2
Essential Media and Reagents
- Dulbecco’s Modified Eagle Medium (D-MEM) containing 4500 mg/L D-glucose, L-glutamine and 110 mg/L sodium pyruvate including 10% fetal bovine serum (FBS) and 1× Penicillin−Streptomycin solution.
- 0.25% Trypsin with EDTA.
2.2.2. Human Neuroblastoma Cell Lines
- SH-SY5Y
Essential Media and Reagents
- Eagle’s Minimum Essential Medium (EMEM) and F-12K (Kaighn’s Modification of Ham’s F-12) medium in equal amounts (1:1) supplemented with 10% FBS and 1× Penicillin−Streptomycin solution.
- 0.25% Trypsin with EDTA.
Thawing of Cell Lines
Sub-Culturing
- These cells grow as a mixture of floating and adherent cells. Remove the medium with the floating cells.
- Rinse the adherent cells with fresh 0.25% trypsin 0.53 mM EDTA solution, add an additional 1 to 2 mL of trypsin solution and let the culture sit at room temperature (or at 37 °C) until the cells detach, and recover the cells by centrifugation (5 min at 500 Gs).
- Add fresh medium, aspirate, combine with the floating cells recovered above and dispense into new flasks. An inoculum of 3 × 103 to 1 × 105 cells/cm2 is recommended.
- Subculture when cell concentration is between 8 × 105 to 1 × 106 cells/cm2 (~80% confluency).
Culture Conditions
2.2.3. Human Neuronal Stem Cell Line (NSC)
Essential Media and Regents
- StemPro® NSC SFM complete medium (100 mL):KnockOut™ D-MEM/F-12 97 mLStemPro® NSC SEM, 200 mM 2 mLGlutaMAX ™-I Supplement 1 mLRecombinant Human Fibroblast Growth Factor Basic 20 µLEpidermal Growth Factor 20 µL
- Fibronectin
- Phosphate-Buffered Saline without Ca2+ and Mg2+ (DPBS)
- TrypLE™ Express Stable Trypsin Replacement Enzyme
Coating Culture Vessels with Fibronectin
- Dilute fibronectin in distilled water. Prepare 1 mg/mL stock solution. Store at −20 °C.
- Dilute stock solution in D-PBS to make working solution of 20 µg/mL.
- Add sufficient amount of working solution to the culture flask or plate to cover the bottom surface of the culture vessels (8–10 mL for T-75 flask or 1 mL per well of 6-well plate or 0.5 mL per well of 12- and 24-well plate.)
- Incubate for 1 h at 37 °C in humidified atmosphere of 5% CO2.
- Remove the vessel and store it at 4 °C until use.
- Discard Fibronectin solution immediately before use and wash with PBS.
Thawing of Cell Line
- Warm 10 mL of KnockOut™ D-MEM/F-12 media in 15 mL centrifuge tube at 37 °C.
- Remove the frozen vial of NSC from nitrogen tank. Immediately transfer to water bath.
- Mix the thawed cells into prewarmed media in a 15 mL centrifuge tube. Spin down the cells at 500 G for 4–5 min.
- Resuspend cells in StemPro® NSC SFM complete medium at 105 cells/mL and culture in a T-25 or T-75 flask coated with fibronectin as described in the previous section.
Culture Procedure
- When cells are about 80% confluent, aspirate the medium and wash with DPBS.
- Add 1 mL of TrypLE™ Express to the flask and incubate at room temperature (or 37 °C) until the cells detach. Add 5–6 mL of fresh media and recover it in a 15 mL centrifuge tube.
- Spin down cells by centrifugation at 500 G for 4–5 min.
- Count cell numbers using hemocytometer as described in Section 2.1.5.
- Plate cells in fresh StemPro® NSC SFM complete medium at specific cell count for experiments on fibronectin-coated plates.
3. Virus Infection Procedure
- Warm virus at room temperature or 37 °C for thawing. When thawed, place on ice.
- Dilute the virus in neuronal maintenance media based on final volumes below (iii). A multiplicity of infection (MOI) of 0.01–10.0 often works well for virus quantification and MTT/XTT assays. Keep dilution on ice.
- Add virus containing media to wells using below final volumes:
- For 6-well plate: 3 mL per well.
- For 12-well plate: 1–2 mL per well.
- For 24-well plate: 0.5–1 mL per well.
- For 48-well plate: 200–400 µL per well.
- For 96-well plate: 100–200 µL per well.
- Incubate at 37 °C for 1 h for virus attachment.
- Remove virus-containing media, wash with PBS and add fresh media at same volumes as described above.
- Place plates in 37 °C CO2 incubator for 2–5 days until 80–100% CPE.
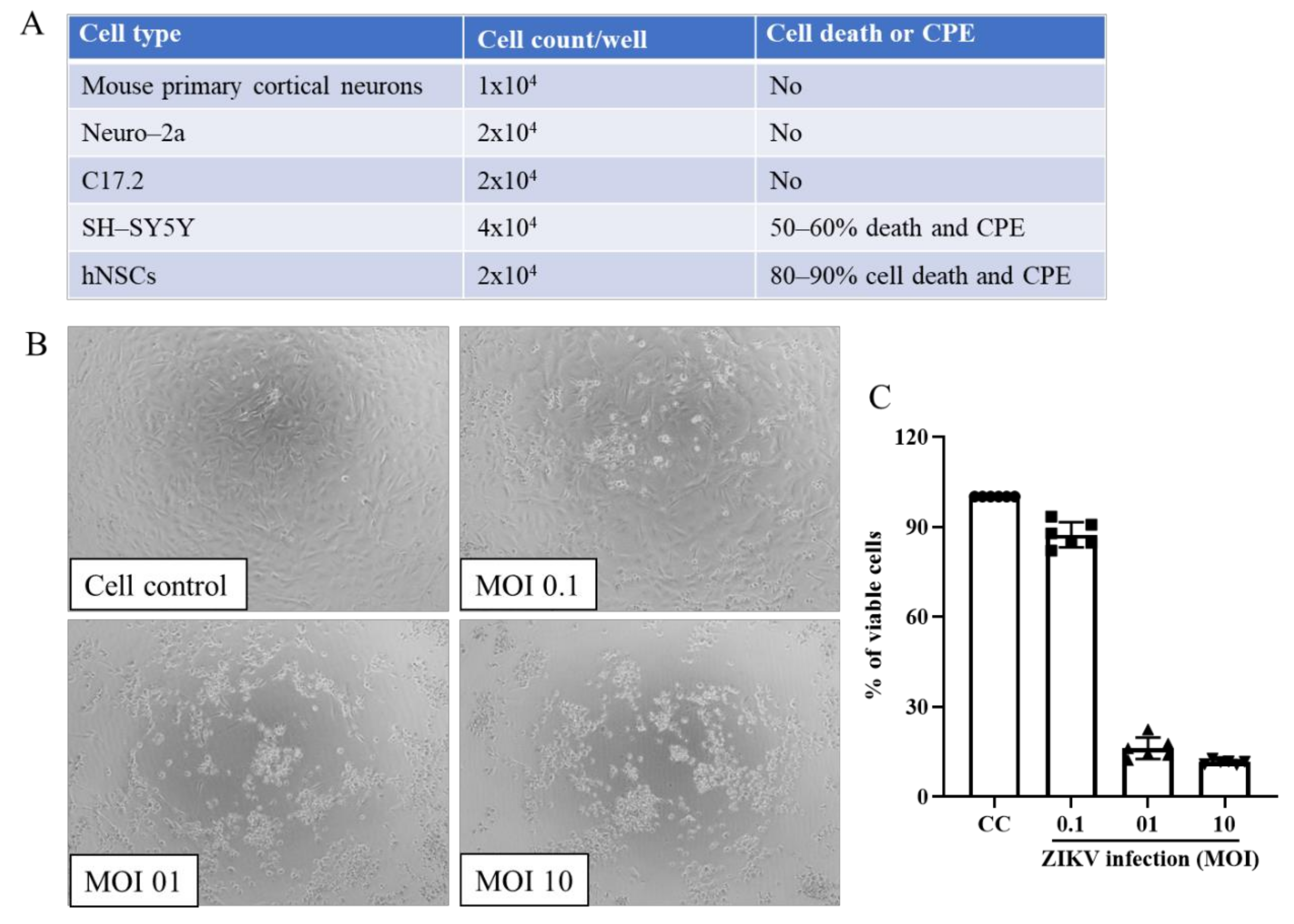
Selection of Cell Types for Drug Study against Zika Virus (ZIKV)
4. Screens of Selected Compounds to Determine Efficacy
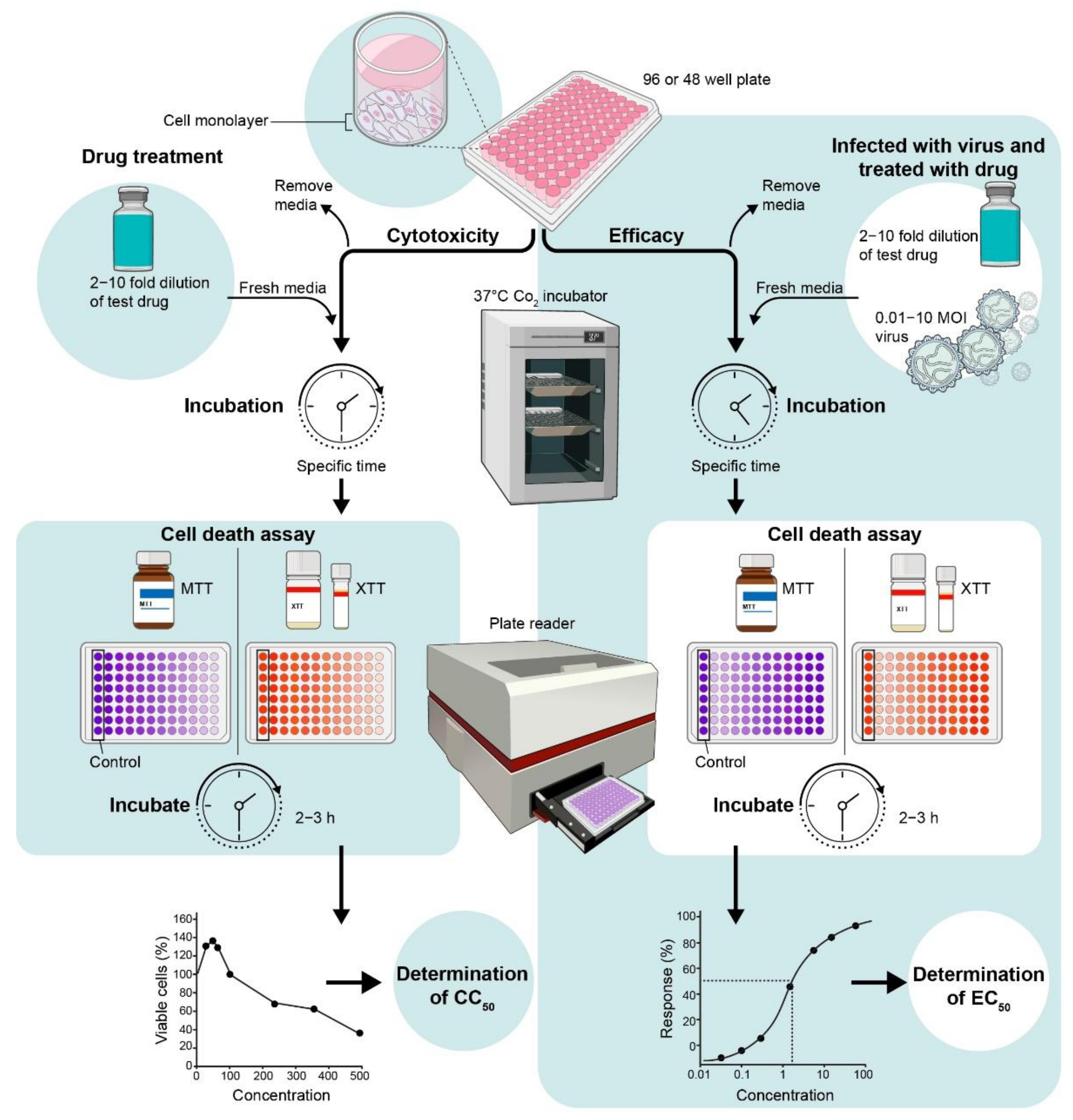
4.1. In Vitro Anti-Viral MTT or XTT Assay
4.1.1. MTT or XTT Reagent Preparation
- Stock concentration: 5 mg/mL in PBS. Dissolve 50 mg of MTT or XTT powder in 10 mL of PBS. Keep on magnetic stirrer with stir bar for 5–10 min to dissolve these reagents completely.
- Sterile filter through 0.22 µm filter. Protect the reconstituted MTT or XTT reagent from light and store at 4 °C.
- STOP solution: Use DMSO or 20% SDS in 50% dimethylformamide. Store at room temperature in fume hood.
4.1.2. Assay Protocol for Cytotoxicity
- Culture cells in 48- or 96-well plate using 103 to 105 cells per well (~80% confluent) and incubate for 4–6 h in 37 °C CO2 incubator.
- Remove the media and add fresh media containing two-fold dilution of the test drug.
- After 2 to 4 days incubation, remove media and wash with PBS (1×).
- Add fresh media containing 1/10 total volume of MTT stock reagent to each well.
- Incubate the plate at 37 °C in dark for 3–4 h. (Reduce incubation times to 2 h if cell density is high.)
- Add 100 to 200 µL of STOP solution into each well and place on shaker for 30 min to 1 h at room temperature to lyse the cells and dissolve the formazan completely. (Alternatively, you can pipet up and down several times to dissolve formazan but avoid air bubbles).
- Transfer 100 µL of the homogenous solution from each well into 96-well plates and measure the absorbance on an ELISA plate reader at 540 nm.
- Determine the percent viability of cells = (mean absorbance of drug treated sample/mean absorbance of control) × 100.
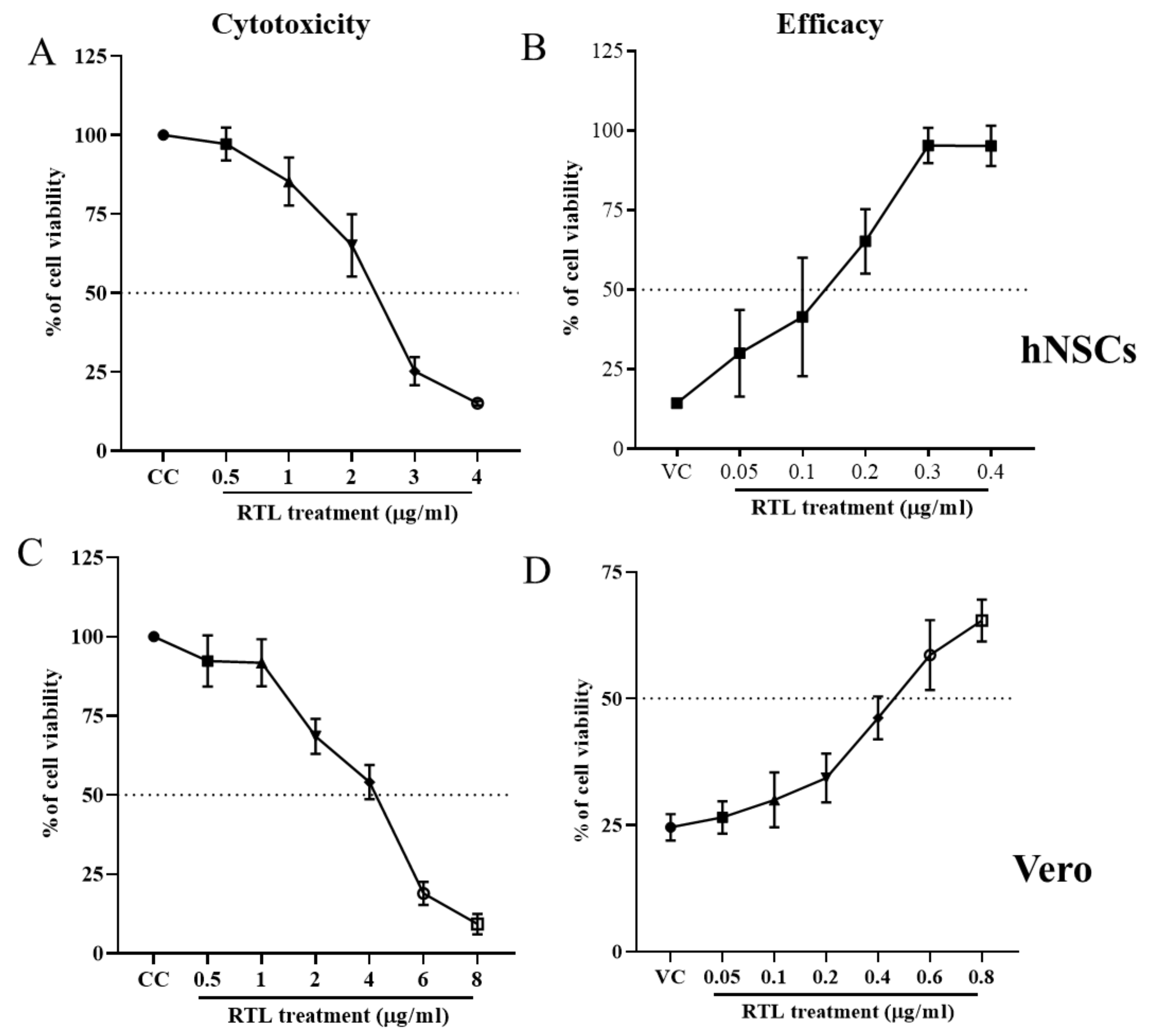
- Determine CC50 (the cytotoxic concentration of the drugs to cause death of 50% of viable cells) by extrapolating dose−response curve.
- After determining the CC50, this concentration should be used as highest dose of the drug for efficacy study. Starting at this initial highest dose, use two to ten (log)-fold dilutions of decreasing concentrations of the drug for dose–response curve in efficacy study.
4.1.3. Assay Protocol for In Vitro Efficacy
- Culture cells in 48- or 96-well plate using 103 to 105 cells per well (~80% confluent) and incubate for 4–6 h in 37 °C CO2 incubator.
- Remove the media and infect with virus at 0.01–10.0 MOI. Add two to ten-fold dilutions of the test drug to wells and incubate 1 h.
- Remove the media and wash with PBS.
- Add fresh media (containing the same amount of test drug as before) to each well and incubate 2–7 days.
- After 2–7 days of incubation, perform MTT/XTT assay (assay protocol for cytotoxicity) starting at step iii.
- After calculating the percentage of viable cells, determine the EC50 or IC50 and SI (CC50/EC50) from the dose–response curve.
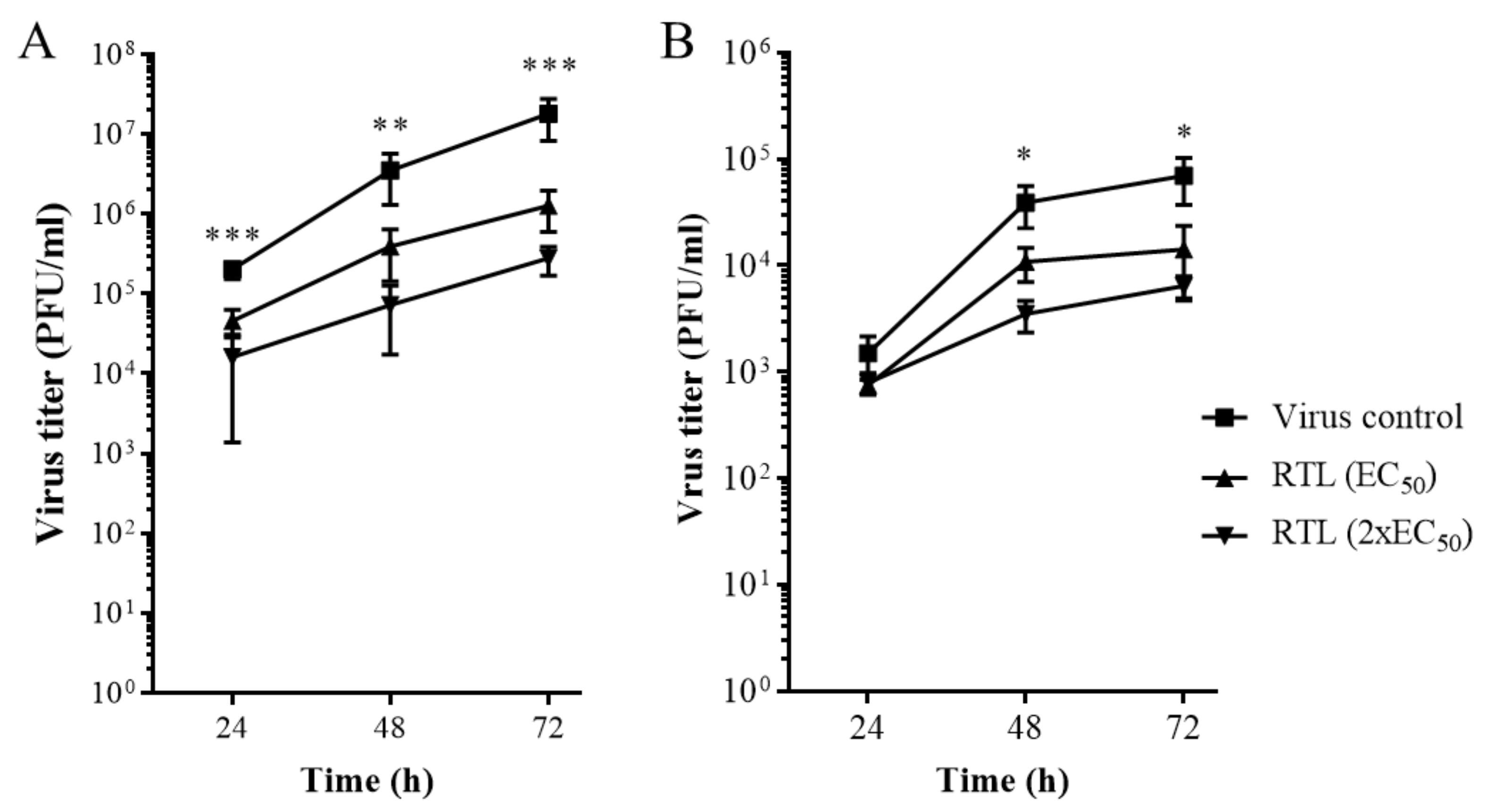
4.2. Determination of Intra-Cellular or Extra-Cellular Virus by Plaque Assay from Drug-Treated Cells
- Plate cells in 12- or 24well plate using 104 to 106 cells per well depending on well size (60–80% confluent). Incubate for 4–6 h in 37 °C CO2 incubator.
- Remove media and add fresh media containing virus at an MOI of 0.01–10.0 MOI. Add desired concentration of the test drug to wells and incubate 1 h.
- Remove media and wash with PBS or fresh media.
- Add fresh media containing same concentration of drug and incubate until desired time points.
4.2.1. For Extra-Cellular Virus
4.2.2. For Intra-Cellular Virus
- At desired time interval, remove the media and wash wells with fresh media.
- Remove cells by trypsin or scrapping and centrifuge.
- Freeze the cell suspension in dry ice or −80 °C freezer to disrupt the cell membrane.
- Prior to plaque assay, thaw the cells using a 37 °C water bath and vortex.
4.2.3. Reagent Preparation for Plaque Assay
- 10% FBS media: DMEM 500 mLPen/Strep 5 mLFBS 50 mL
- 2% FBS DMEM: DMEM 500 mLPen/Strep 5 mLFBS 10 mL
- 1.5% CMC: In a sterile hood, add 7.5 g of sterilized CMC sodium salt (see below) to 500 mL bottle of MEM media.Shake at 37 °C until CMC is dissolved.Sterile CMC sodium salt (Sigma Aldrich C4888): Measure 7.5 g into 20 mL beaker and cover with aluminum foil.Sterilize in autoclave at 250 °F for 20 min and dry cycle for 30 min.
- 10% formaldehyde: 32% formaldehyde 135 mL.Water 365 mL.
- 0.35% crystal violet: crystal violet 1.7 g into 95% ethanol (75 mL) until dissolved.Bring volume to 500 mL with nanopure water. Can be stored at room temperature indefinitely.
4.2.4. Procedure
- Grow Vero cells to confluency in T-25; trypsinize and resuspend cells in total 12.5 mL of 10% FBS media. Add 0.5 mL to each well of 24-well plate and incubate at 37 °C incubator. Can fill just one well and check density before doing entire plate. Keep cells well mixed in flask while dispensing.
- Allow plates to become confluent.
- Viral dilutions: Add 900 µL of 2% FBS-DMEM to small snap-cap tubes. Add 100 µL of sample to first tube, mix well, change pipette tip and transfer 100 µL to second tube. Repeat this process until desired dilutions are reached. Dilutions of 10−1 to 10−8 are usually sufficient to determine the amount of virus in each sample. Always run a positive control (known concentration of virus) and a negative control (media alone) for each assay.NOTE: If you want to conserve virus, make first dilution in 450 µL of media using 50 µL of virus. Rest of dilution scheme is the same. This dilution is rarely plated.
- Discard media from plated Vero cells into an appropriate waste container, leaving small amount of media to keep cell monolayer from drying out.
- Starting from highest dilution, add 200 µL of diluted sample to each of three wells. Mix tube well before adding media/virus to the cells.
- Place in incubator for 1 h for adsorption and infection.
- Add 0.5 mL of warm 1.5% CMC directly to wells with media/virus. Place back in incubator for 5 days.NOTE: Do not touch the pipette to the wells while adding CMC. Add CMC from a height, otherwise you will contaminate the CMC in the bottle with virus.
- After 5 days, add 10% formaldehyde directly to fill wells, incubate at room temperature for at least 1 h.
- Pour off formaldehyde solution into appropriate container, rinse wells in tap water several times to remove all CMC. Do not spray directly into wells.
- Add enough 0.35% crystal violet to cover bottom of each well, and let sit for 10 min.
- Pour off stain, and again wash gently with tap water as in step ix.
- Allow to dry at room temperature.
- Plaque counting:
- ○
- Pick dilutions where plaques can be easily distinguished from each other for counting.
- ○
- Calculations: Virus titer (PFU/mL) = Number of Plaques x dilution factor x infection volume factor.
- ○
- Infection Volume factor:For, 200 µL = 5For, 100 µL = 10
5. Basics in Mode of Action Study
5.1. Determining When the Drug Affects Virus Infection: Time of Addition Assay
Assay Protocol for Time of Addition Assay
- Pre-treatment:
- Culture cells in 48- or 96-well plate at 104 to 105 cells per well and incubate for 6–9 h in incubator containing 5% CO2 at 37 °C.
- Remove the media and add fresh media containing 2- to k10-fold higher EC50 concentration of the test drug.
- After 1–3 h incubation, remove the media containing test drug and wash with PBS.
- Infect with virus at 0.01–10.0 MOI and incubate for 2–7 days.
- Read out of MTT/XTT assay and/or virus titers.
- Co-treatment:
- Culture cells in 48- or 96-well plate using 2 × 104 or 1 × 104 cells per well and incubate for 6–9 h in incubator containing 5% CO2 at 37 °C.
- Remove the media and infect with virus at 0.01–10.0 MOI as well as add 2-to-10-fold higher EC50 concentration of the test drug and incubate for 1 h.
- Remove the media containing unbound virus and wash with PBS.
- Add fresh media (without drug) and incubate for 2–7 days.
- Read out of MTT/XTT assay and/or virus titers.
- Post-treatment:
- Culture cells in 48- or 96- well plate using 103 to 106 cells per well and incubate for 6–9 h in incubator containing 5% CO2 at 37 °C.
- Remove the media and infect with virus at 0.01–10.0 MOI.
- Remove the media containing unbound virus and wash with PBS.
- Add fresh media containing 2-to-10-fold EC50 of test drug and incubate for 2–7 days.
- Read out of MTT/XTT assay and/or virus titers.
- Throughout-treatment: This is the combination of previously described three treatment regimens (Pre-, Co- and Post-treatment with drug).
5.2. Virus Attachment Assay
Assay Protocol for Virus Attachment Assay
- Grow cells in 12- or 24- well plate depending on well size.
- Incubate overnight.
- Prechill cells at 4 °C for 1 h.
- Mix same volume of the drug (at 4× EC50 concentration) and virus (2× specific MOI).
- Add drug and virus mixture to the cells and incubate for 3 h at 4 °C or at top of ice.
- Wash two to three time with ice-cold PBS to remove unbound virus.
- Add fresh medium to the cells and incubate for either an early time point (1 to 2 h) or late time point (24 or 48 h).
- For early time point: Remove supernatant and perform qRT-PCR or immunohistochemistry to detect efficacy of the drug.
- For late time point: Harvest cell supernatant after desired time points and titrate the viruses using previously described plaque assay.
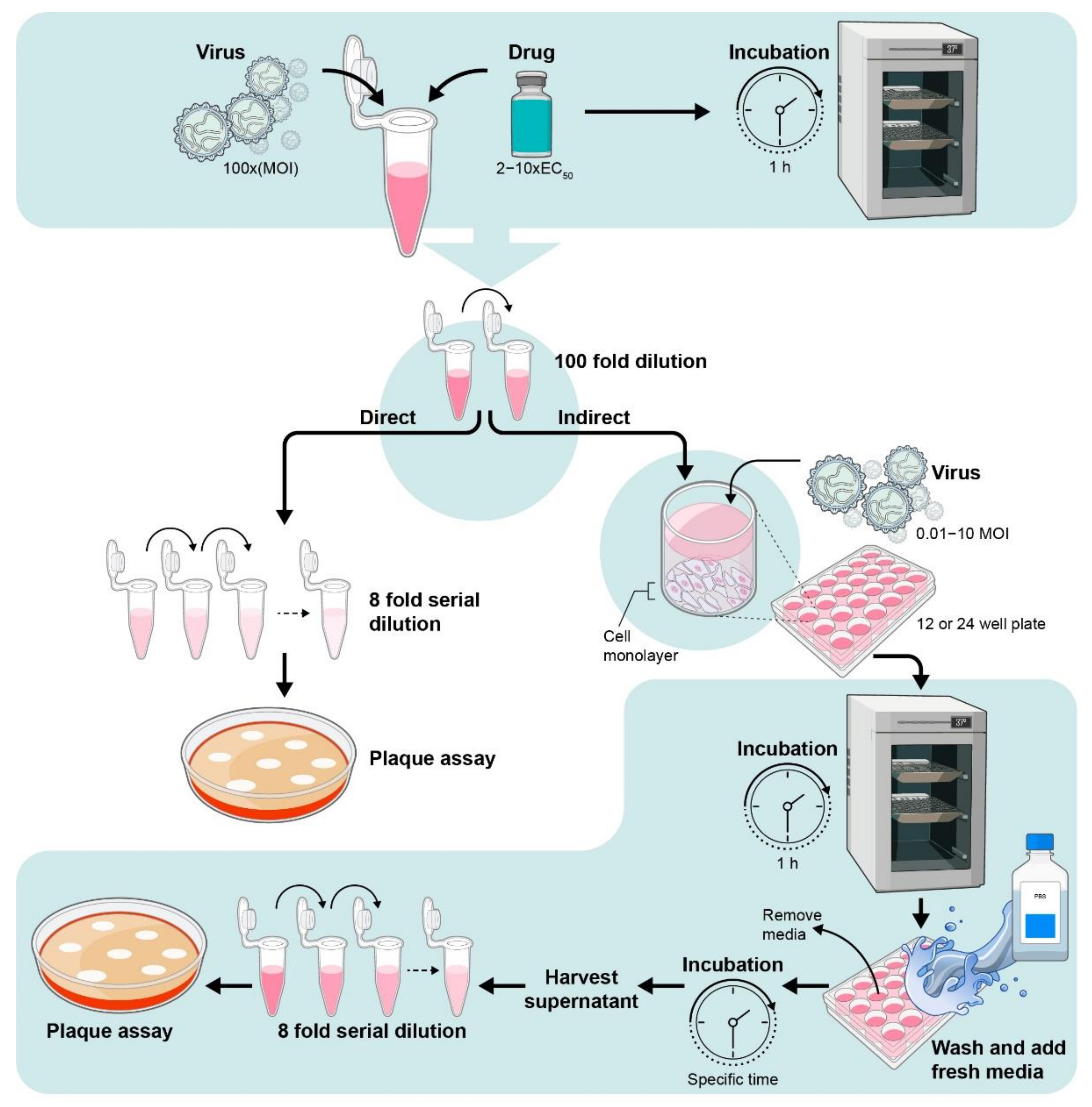
5.3. Viral Inactivation Assay
5.3.1. Assay Protocol for Viral Inactivation Assay
- Mix the drug (at 2–10-fold EC50 concentration) with virus (103 to 106 PFU/mL) and incubate for 1 h in 37 °C.
- Dilute 100-fold (10–1000 PFU/well) with media containing 2% FBS to get subtherapeutic concentration of the drug.
5.3.2. Direct Method
- Titrate the viruses using previously described plaque assay.
5.3.3. Indirect Method
- Plate cells in 12- or 24-well plate using 104 × 106 cells per well depending on well size.
- Incubate cells overnight.
- Remove media and add diluted virus and drug mix and incubate 1 h.
- Remove media and wash with PBS or fresh media.
- Add fresh media and reincubate cells in incubator.
- Harvest cell supernatant after desired time points
- Titrate the viruses using previously described plaque assay.
- The 100-fold dilution helps to minimize the effect of the drugs in its effective dose and prevent significant interaction with host cells.
- Use neutralizing antibody as a positive control [24].
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kennedy, P.G.E. Viral Encephalitis: Causes, Differential Diagnosis, and Management. J. Neurol. Neurosurg. Psychiatry 2004, 7, i10–i15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ames, J.; Yadavalli, T.; Suryawanshi, R.; Hopkins, J.; Agelidis, A.; Patil, C.; Fredericks, B.; Tseng, H.; Valyi-Nagy, T.; Shukla, D. OPTN is a host intrinsic restriction factor against neuroinvasive HSV-1 infection. Nat. Commun. 2021, 12, 5401. [Google Scholar] [CrossRef] [PubMed]
- Arribas, J.R.; Storch, G.A.; Clifford, D.B.; Tselis, A.C. Cytomegalovirus encephalitis. Ann. Intern. Med. 1996, 125, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Doja, A.; Bitnun, A.; Ford Jones, E.L.; Richardson, S.; Tellier, R.; Petric, M.; Heurter, H.; MacGregor, D. Pediatric Epstein–Barr virus—Associated encephalitis: 10-year review. J. Child. Neurol. 2006, 21, 384–391. [Google Scholar] [CrossRef]
- Lizzi, J.; Hill, T.; Jakubowski, J. Varicella Zoster Virus Encephalitis. Clin. Pract. Cases Emerg. Med. 2019, 3, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Anastasina, M.; Domanska, A.; Palm, K.; Butcher, S. Human picornaviruses associated with neurological diseases and their neutralization by antibodies. J. Gen. Virol. 2017, 98, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Enders, G. Paramyxoviruses. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; Chapter 59. [Google Scholar]
- Wong, S.C.; Ooi, M.H.; Wong, M.N.L.; Tiob, P.H.; Solomonc, T.; Cardosab, M.J. Late presentation of Nipah virus encephalitis and kinetics of the humoral immune response. J Neurol. Neurosurg. Psychiatry 2001, 71, 552–554. [Google Scholar] [CrossRef]
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.B.; Peterson, K.E. Throw out the Map: Neuropathogenesis of the Globally Expanding California Serogroup of Orthobunyaviruses. Viruses 2019, 11, 794. [Google Scholar] [CrossRef] [Green Version]
- Burrell, C.J.; Howard, C.R.; Murphy, F.A. Chapter 35—Togaviruses. In Fenner and White’s Medical Virology, 5th ed.; Burrell, C.J., Howard, C.R., Murphy, F.A., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 477–491. [Google Scholar]
- Rupprecht, C.E. Rhabdoviruses: Rabies virus. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; Chapter 61. [Google Scholar]
- Romero, J.R.; Simonsen, K.A. Powassan encephalitis and Colorado tick fever. Infect. Dis. Clin. N. Am. 2008, 22, 545–559. [Google Scholar] [CrossRef]
- Deleidi, M.; Isacson, O. Viral and inflammatory triggers of neurodegenerative diseases. Sci. Transl. Med. 2012, 4, 121ps3. [Google Scholar] [CrossRef] [Green Version]
- Berth, S.H.; Leopold, P.L.; Morfini, G.N. Virus-induced neuronal dysfunction and degeneration. Front. Biosci. 2009, 14, 5239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyuncu, O.O.; Hogue, I.B.; Enquist, L.W. Virus infections in the nervous system. Cell Host Microbe 2013, 13, 379–393. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.P.; Enquist, L.W. Axonal spread of neuroinvasive viral infections. Trends Microbiol. 2015, 23, 283–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksamit, A.J., Jr. Treatment of Viral Encephalitis. Neurol. Clin. 2021, 39, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Lehman, C.W.; Stewart, A.K.; Panny, L.; Bracci, N.; Wright, J.L.C.; Paige, M.; Strangman, W.K.; Kehn-Hall, K. Homoseongomycin, a compound isolated from marine actinomycete bacteria K3-1, is a potent inhibitor of encephalitic alphaviruses. Antivir. Res. 2021, 191, 105087. [Google Scholar] [CrossRef] [PubMed]
- Warrell, M.; Warrell, D.A.; Tarantola, A. The Imperative of Palliation in the Management of Rabies Encephalomyelitis. Trop. Med. Infect. Dis. 2017, 2, 52. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, W. Clinical features and surgical treatment of epilepsy after viral encephalitis. Brain Sci. Adv. 2019, 5, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Piret, J.; Boivin, G. Resistance of herpes simplex viruses to nucleoside analogues: Mechanisms, prevalence, and management. Antimicrob. Agents Chemother. 2011, 55, 459–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lycke, J.; Malmestrom, C.; Stahle, L. Acyclovir levels in serum and cerebrospinal fluid after oral administration of valacyclovir. Antimicrob. Agents Chemother. 2003, 47, 2438–2441. [Google Scholar] [CrossRef] [Green Version]
- Ojha, D.; Winker, W.C.; Leung, J.M.; Woods, T.A.; Chen, C.Z.; Nair, V.; Taylor, K.; Yeh, C.D.; Tawa, G.J.; Larson, C.L.; et al. Rottlerin inhibits La Crosse virus-induced encephalitis in mice and blocks release of replicating virus from the Golgi body in neuronsn. Nat. Microbiol. 2021, 6, 1398–1409. [Google Scholar] [CrossRef] [PubMed]
- Barrows, N.J.; Campos, R.K.; Powell, S.T.; Prasanth, K.R.; Schott-Lerner, G.; Soto-Acosta, R.; Galarza-Muñoz, G.; McGrath, E.L.; Urrabaz-Garza, R.; Gao, J.; et al. A Screen of FDA-Approved Drugs for Inhibitors of Zika Virus Infection. Cell Host Microbe. 2016, 20, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Severson, W.E.; Shindo, N.; Sosa, M.; Fletcher, T., 3rd; White, E.L.; Ananthan, S.; Jonsson, C.B. Development and validation of a high-throughput screen for inhibitors of SARS CoV and its application in screening of a 100,000-compound library. J. Biomol. Screen. 2007, 12, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, H.; Baba, M.; Shigeta, S. An application of tetrazolium (MTT) colorimetric assay for the screening of anti-herpes simplex virus compounds. J. Virologic. Meth. 1991, 33, 61–71. [Google Scholar] [CrossRef]
- Müller, J.A.; Harms, M.; Schubert, A.; Mayer, B.; Jansen, S.; Herbeuval, J.P.; Michel, D.; Mertens, T.; Vapalahti, O.; Schmidt-Chanasit, J.; et al. Development of a high-throughput colorimetric Zika virus infection assay. Med. Microbiol. Immunol. 2017, 206, 175–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, C.J.; Li, C.L.; Tai, C.J.; Wang, C.K.; Lin, L.T. Early Viral Entry Assays for the Identification and Evaluation of Antiviral Compounds. J. Vis. Exp. 2015, 105, e53124. [Google Scholar] [CrossRef] [Green Version]
- Green, N.; Ott, R.D.; Isaacs, R.J.; Fang, H. Cell-based Assays to Identify Inhibitors of Viral Disease. Expert Opin. Drug Discov. 2008, 3, 671–676. [Google Scholar] [CrossRef] [Green Version]
- Kovalevich, J.; Langford, D. Considerations for the Use of SH-SY5Y Neuroblastoma Cells in Neurobiology. Methods Mol. Bio. 2016, 1078, 9–21. [Google Scholar]
- Ryder, E.F.; Snyder, E.Y.; Cepko, C.L. Establishment and characterization of multipotent neural cell lines using retrovirus vector-mediated oncogene transfer. J. Neurobiol. 1990, 21, 356–375. [Google Scholar] [CrossRef] [PubMed]
- Butchi, N.B.; Du, M.; Peterson, K.E. Interactions between TLR7 and TLR9 agonists and receptors regulate innate immune responses by astrocytes and microglia. Glia 2010, 58, 650–664. [Google Scholar] [CrossRef] [Green Version]
- Jang, Y.; Shin, J.S.; Lee, M.K.; Jung, E.; An, T.; Kim, U.-I.; Kim, K.; Kim, M. Comparison of Antiviral Activity of Gemcitabine with 2′-Fluoro-2′-Deoxycytidine and Combination Therapy with Remdesivir against SARS-CoV-2. Int. J. Mol. Sci. 2021, 22, 1581. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Shin, J.S.; Yoon, Y.S.; Go, Y.Y.; Lee, H.W.; Kwon, O.S.; Park, S.; Park, M.-S.; Kim, M. Salinomycin Inhibits Influenza Virus Infection by Disrupting Endosomal Acidification and Viral Matrix Protein 2 Function. J. Virol. 2018, 92, e01441-18. [Google Scholar] [CrossRef] [Green Version]
- Qiao, J.; Li, Y.-S.; Zeng, R.; Liu, F.-L.; Luo, R.-H.; Huang, C.; Wang, Y.-F.; Zhang, J.; Quan, B.; Shen, C.; et al. SARS-CoV-2 Mpro inhibitors with antiviral activity in a transgenic mouse model. Science 2021, 371, 1374–1378. [Google Scholar] [CrossRef] [PubMed]
- Gowen, B.B.; Wong, M.H.; Jung, K.H.; Sanders, A.B.; Mendenhall, M.; Bailey, K.W.; Furuta, Y.; Sidwell, R.W. In vitro and in vivo activities of T-705 against arenavirus and bunyavirus infections. Antimicrob. Agents Chemother. 2007, 51, 3168–3176. [Google Scholar] [CrossRef] [Green Version]
- Food and Drug Administration. Guidance for Industry: Antiviral Product Development: Conducting and Submitting Virology Studies to the Agency. 2006. Available online: https://www.fda.gov/ucm/groups/fdagov-public/@fdagov-drugs-gen/documents/document/ucm070953.pdf (accessed on 16 November 2021).
- Gupta, R.; Hill, E.L.; McClernon, D.; Davis, G.; Selke, S.; Corey, L.; Wald, A. Acyclovir Sensitivity of Sequential Herpes Simplex Virus Type 2 Isolates from the Genital Mucosa of Immunocompetent Women. J. Infec. Dis. 2005, 192, 1102–1107. [Google Scholar] [CrossRef] [Green Version]
- Tse, E.G.; Korsik, M.; Todd, M.H. The past, present and future of anti-malarial medicines. Malar. J. 2019, 18, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delvecchio, R.; Higa, L.M.; Pezzuto, P.; Valadão, A.L.; Garcez, P.P.; Monteiro, F.L.; Loiola, E.C.; Dias, A.A.; Silva, F.J.M.; Aliota, M.T.; et al. Chloroquine, an Endocytosis Blocking Agent, Inhibits Zika Virus Infection in Different Cell Models. Viruses 2016, 8, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maioli, E.; Greci, L.; Soucek, k.; Hyzdalova, M.; Pecorelli, A.; Fortino, V.; Valacchet, G. Rottlerin inhibits ROS formation and prevents NFkappaB activation in MCF-7 and HT-29 cells. J. Biomed. Biotechnol. 2009, 2009, 742936. [Google Scholar] [CrossRef] [Green Version]
- Ishii, R.; Horie, M.; Saito, K.; Arisawa, M.; Kitanaka, S. Inhibitory effects of phloroglucinol derivatives from Mallotus japonicus on nitric oxide production by a murine macrophage-like cell line, RAW 264.7, activated by lipopolysaccharide and interferon-gamma. Biochim. Biophys. Acta 2001, 1568, 74–82. [Google Scholar] [CrossRef]
- Ishii, R.; Horie, M.; Saito, K.; Arisawa, M.; Kitanaka, S. Prostaglandin E(2) production and induction of prostaglandin endoperoxide synthase-2 is inhibited in a murine macrophage-like cell line, RAW 264.7, by Mallotus japonicus phloroglucinol derivatives. Biochim. Biophys. Acta 2002, 1571, 115–123. [Google Scholar] [CrossRef]
- Shivshankar, P.; Lei, L.; Wang, J.; Zhong, G. Rottlerin inhibits chlamydial intracellular growth and blocks chlamydial acquisition of sphingolipids from host cells. Appl. Environ. Microbiol. 2008, 74, 1243–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, S.F.H.; Yoshida, I.; Butt, F.; Yusuf, M.A.; Usmanghani, K.; Kadowaki, M.; Sugiyama, T. Potent bactericidal constituents from Mallotus philippinensis against clarithromycin and metronidazole resistant strains of Japanese and Pakistani Helicobacter pylori. Biol. Pharm. Bull. 2009, 32, 631–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maioli, E.; Torricelli, C.; Valacchi, G. Rottlerin and cancer: Novel evidence and mechanisms. Sci. World J. 2012, 2012, 350826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.; Wang, L.; Yin, X.; Zhao, Z.; Hou, Y.; Ye, X.; Zhou, X.; Wang, Z. Rottlerin exhibits anti-cancer effect through inactivation of S phase kinase-associated protein 2 in pancreatic cancer cells. Am. J. Cancer Res. 2016, 6, 2178–2191. [Google Scholar]
- Gschwendt, M.; Müller, H.J.; Kielbassa, K.; Zang, R.; Kittstein, W.; Rincke, G.; Marks, F. Rottlerin, a novel protein kinase inhibitor. Biochem. Biophys. Res. Commun. 1994, 199, 93–98. [Google Scholar] [CrossRef]
- Bain, J.; Plater, L.; Elliott, M.; Shpiro, N.; Hastie, C.J.; McLauchlan, H.; Klevernic, I.; Arthur, J.S.C.; Alessi, D.R.; Cohen, P. The selectivity of protein kinase inhibitors: A further update. Biochem. J. 2007, 408, 297–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viegas, D.J.; Edwards, T.G.; Bloom, D.C.; Abreu, P.A. Virtual screening identified compounds that bind to cyclin dependent kinase 2 and prevent herpes simplex virus type 1 replication and reactivation in neurons. Antivir. Res. 2019, 172, 104621. [Google Scholar] [CrossRef]
- Bhatia, H.K.; Singh, H.; Grewal, N.; Natt, N.K. Sofosbuvir: A novel treatment option for chronic hepatitis C infection. J. Pharmacol. Pharmacother. 2014, 5, 278–284. [Google Scholar]
- Baz, M.; Boivin, G. Antiviral Agents in Development for Zika Virus Infections. Pharmaceuticals 2019, 12, 101. [Google Scholar] [CrossRef] [Green Version]
- Reznik, S.E.; Ashby, C.R., Jr. Sofosbuvir: An antiviral drug with potential efficacy against Zika infection. Int. J. Infect. Dis. 2017, 55, 29–30. [Google Scholar] [CrossRef] [Green Version]
- Bullard-Feibelman, K.M.; Govero, J.; Zhu, Z.; Salazar, V.; Veselinovic, M.; Diamond, M.S.; Geiss, B.J. The FDA-approved drug sofosbuvir inhibits Zika virus infection. Antivir. Res. 2017, 137, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.-T.; Chen, T.-Y.; Chung, C.-Y.; Noyce, R.S.; Grindley, T.B.; McCormick, C.; Lin, T.-C.; Wang, G.-H.; Lin, C.-C.; Richardson, C.D. Hydrolyzable tannins (chebulagic acid and punicalagin) target viral glycoprotein-glycosaminoglycan interactions to inhibit Herpes Simplex virus 1 entry and cell-to-cell spread. J. Virol. 2011, 85, 4386–4398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogbole, O.O.; Akinleye, T.E.; Segun, P.A.; Faleye, T.C.; Adeniji, A.J. In vitro antiviral activity of twenty-seven medicinal plant extracts from Southwest Nigeria against three serotypes of echoviruses. Virol. J. 2018, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Keyaerts, E.; Li, S.; Vijgen, L.; Rysman, E.; Verbeeck, J.; Ranst, M.V.; Maes, P. Antiviral Activity of Chloroquine against Human Coronavirus OC43 Infection in Newborn Mice. Antimicrob. Agents Chemother. 2009, 53, 3416–3421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheahan, T.P.; Sims, A.C.; Zhou, S.; Graham, R.L.; Pruijssers, A.J.; Agostini, M.L.; Leist, S.R.; Schäfer, A.; Dinnon, K.H., 3rd; Stevens, L.J.; et al. An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 in human airway epithelial cell cultures and multiple coronaviruses in mice. Sci. Transl. Med. 2020, 12, eabb5883. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Point(s) | Possible Outcome |
|---|---|
| Pre-treatment |
|
| Pre- and Co- treatment |
|
| Co-treatment |
|
| Co- and Post- treatment |
|
| Post-treatment |
|
| Throughout- treatment | Positive control for all other time of addition as it combines all time points. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ojha, D.; Woods, T.A.; Peterson, K.E. Drug-Screening Strategies for Inhibition of Virus-Induced Neuronal Cell Death. Viruses 2021, 13, 2317. https://doi.org/10.3390/v13112317
Ojha D, Woods TA, Peterson KE. Drug-Screening Strategies for Inhibition of Virus-Induced Neuronal Cell Death. Viruses. 2021; 13(11):2317. https://doi.org/10.3390/v13112317
Chicago/Turabian StyleOjha, Durbadal, Tyson A. Woods, and Karin E. Peterson. 2021. "Drug-Screening Strategies for Inhibition of Virus-Induced Neuronal Cell Death" Viruses 13, no. 11: 2317. https://doi.org/10.3390/v13112317
APA StyleOjha, D., Woods, T. A., & Peterson, K. E. (2021). Drug-Screening Strategies for Inhibition of Virus-Induced Neuronal Cell Death. Viruses, 13(11), 2317. https://doi.org/10.3390/v13112317