
Characterization of Four Novel dsRNA Viruses Isolated from Mucor hiemalis Strains

 , , , ,
, , , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Cultivation
2.2. Isolation of dsRNA Molecules
2.3. Purification and Examination of the Virus Particles
2.4. Sequencing Library Generation, NGS Sequencing and Bioinformatic Analysis of Sequencing Data
2.5. cDNA Synthesis and Sequencing of the dsRNA Molecules
2.6. Sequence and Phylogenetic Analysis
2.7. Hybridization Studies
3. Results
3.1. Presence of dsRNA Elements in the Tested Mucor Hiemalis Strains
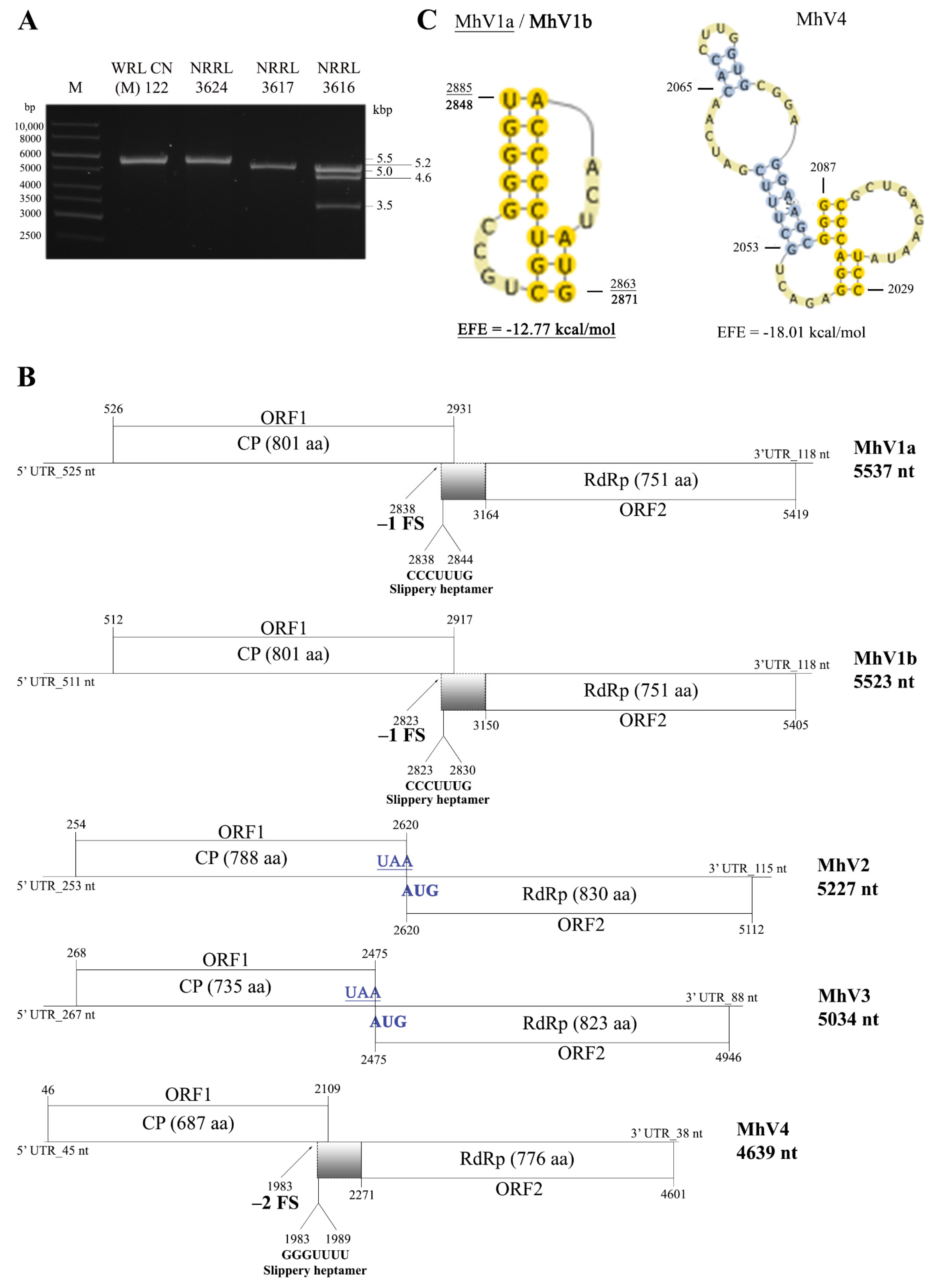
3.2. Sequence and Genome Organization of the Detected dsRNA Elements
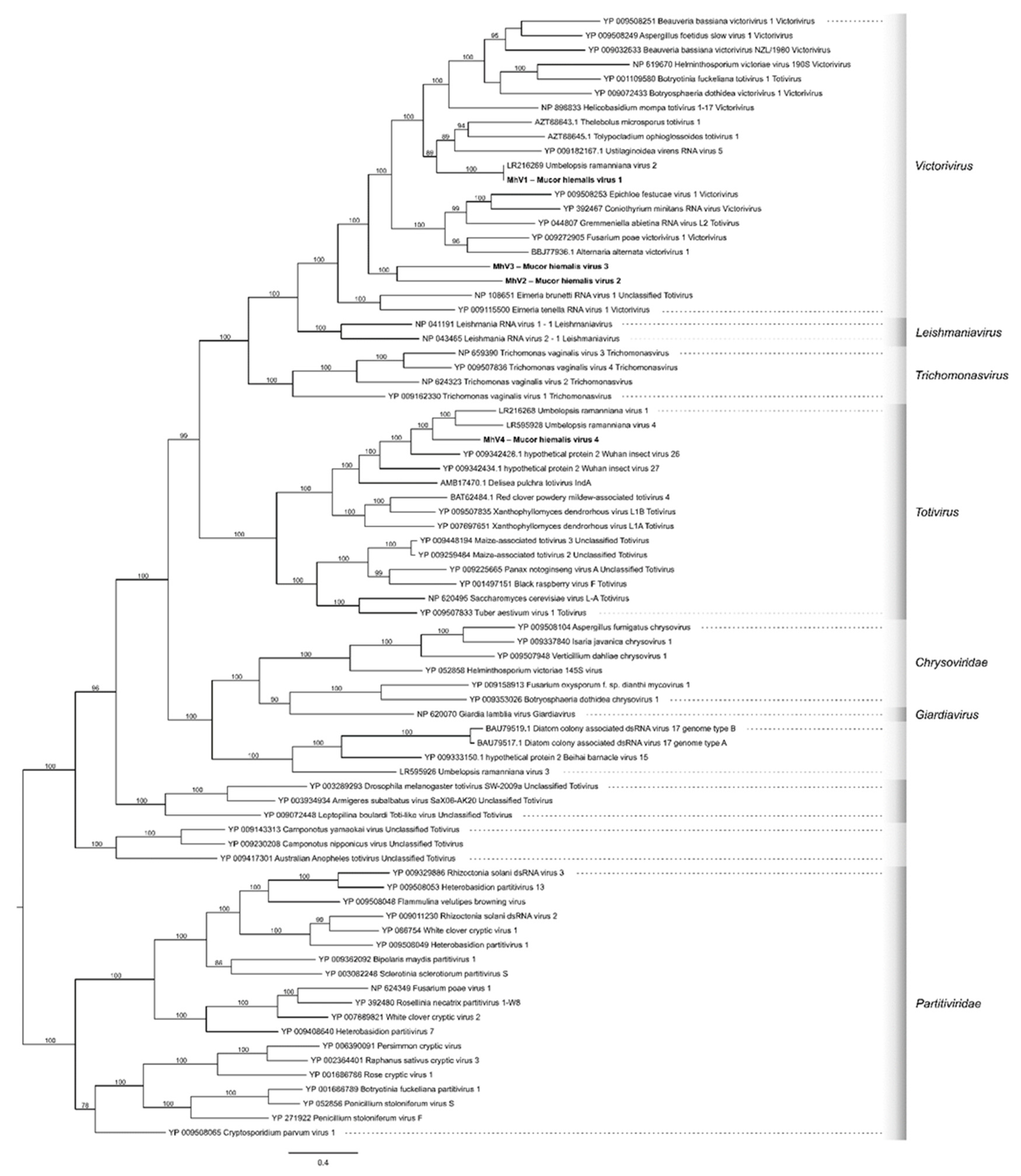
3.3. Phylogenetic Analysis of the M. hiemalis Viruses
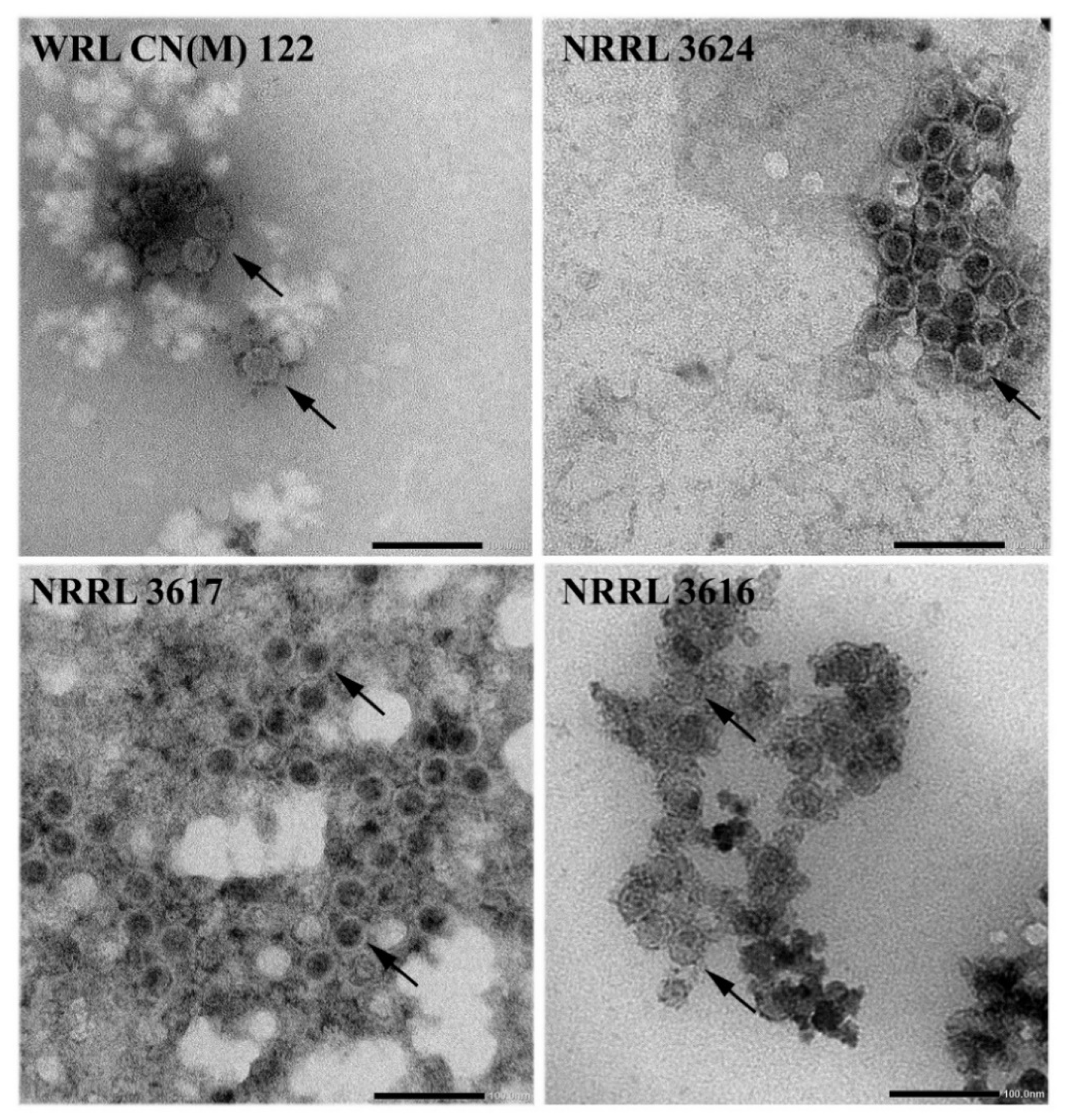
3.4. Detection of Virus Particles in M. hiemalis Strains
3.5. Hybridization Analysis of the dsRNA Patterns
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xie, J.; Jiang, D. New Insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [Green Version]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, M.; Yu, J.; Kim, K.H. Five questions about mycoviruses. PLoS Pathog. 2015, 11, e1005172. [Google Scholar] [CrossRef]
- Ghabrial, S.A. and Suzuki, N. Viruses of Plant Pathogenic Fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef]
- Ghabrial, S.A. Origin, Adaptation and Evolutionary Pathways of Fungal Viruses. Virus Genes 1998, 16, 119–131. [Google Scholar] [CrossRef]
- Wickner, R.B.; Ghabrial, S.A.; Nibert, M.L.; Patterson, J.L.; Wang, C.C. Family Totiviridae. In Ninth Report of the International Committee for the Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Virus Taxonomy; Elsevier Academic Press: San Diego, CA, USA, 2011; pp. 639–650. [Google Scholar]
- Kartali, T.; Nyilasi, I.; Szabó, B.; Kocsubé, S.; Patai, R.; Polgár, T.F.; Nagy, G.; Vágvölgyi, C.; Papp, T. Detection and molecular characterization of novel dsRNA viruses related to the Totiviridae family in Umbelopsis ramanniana. Front. Cell Infect. Microbiol. 2019, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Myers, J.M.; Bonds, A.E.; Clemons, R.A.; Thapa, N.A.; Simmons, D.R.; Carter-House, D.; Ortanez, J.; Liu, P.; Miralles-Durán, A.; Desirò, A.; et al. Survey of early-diverging lineages of fungi reveals abundant and diverse mycoviruses. mBio 2020, 11, e02027-20. [Google Scholar] [CrossRef]
- Papp, T.; Nyilasi, I.; Fekete, C.; Ferenczy, L.; Vágvölgyi, C. Presence of double-stranded RNA and virus-like particles in Rhizopus isolates. Can. J. Microbiol. 2001, 47, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Hafez, E.E.; Aseel, D.G.; Mostafa, S. Two novel mycoviruses related to Geminivirus isolated from the soil-borne fungi Macrophomina phaseolina (Tassi) Goid. and Mucor racemosus Bull. Biotechnol. Biotechnol. Equip. 2013, 27, 4222–4226. [Google Scholar] [CrossRef] [Green Version]
- Barata, R.R.; Vianez-Júnior, J.L.S.G.; Nunes, M.R.T. Transcriptomic analysis of Mucor irregularis containing a negative single-stranded RNA mycovirus. Microbiol. Resour. Announc. 2019, 8, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espino-Vázquez, A.N.; Bermúdez-Barrientos, J.R.; Cabrera-Rangel, J.F.; Córdova-López, G.; Cardoso-Martínez, F.; Martínez-Vázquez, A.; Camarena-Pozos, D.A.; Mondo, S.J.; Pawlowska, T.E.; Abreu-Goodger, C.; et al. Narnaviruses: Novel players in fungal–bacterial symbioses. ISME J. 2020, 14, 1743–1754. [Google Scholar] [CrossRef]
- Vágvölgyi, C.; Magyar, K.; Papp, T.; Vastag, M.; Ferenczy, L.; Hornok, L.; Fekete, C. Detection of double-stranded RNA molecules and virus-like particles in different Mucor species. Ant. Van. Leeuwenhoek 1998, 73, 207–210. [Google Scholar] [CrossRef]
- Leach, J.; Finkelstein, D.B.; Rambosek, J.A. Rapid miniprep of DNA from filamentous fungi. Fungal Genet. Rep. 1986, 33, 9. [Google Scholar] [CrossRef] [Green Version]
- Morris, T.J. and Dodds, J.A. Isolation and analysis of double-stranded RNA from virus-infected plant and fungal tissue. Phytopathology 1979, 69, 854–858. [Google Scholar] [CrossRef] [Green Version]
- Lot, H.; Marrou, J.; Quiot, J.B.; Esvan, C. Contribution a l′etude du virus de la mosaique du concombre (CMV). II. Methode de purification rapide du virus. Ann. Phytopathol. 1972, 4, 25–38. [Google Scholar]
- Langmead, B. and Salzberg, S. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maan, S.; Rao, S.; Maan, N.S.; Anthony, S.J.; Attoui, H.; Samuel, A.R. Rapid cDNA synthesis and sequencing techniques for the genetic study of bluetongue and other dsRNA viruses. J. Virol. Methods 2007, 143, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Potgieter, A.C.; Page, N.A.; Liebenberg, J.; Wright, I.M.; Landt, O.; van Dijk, A.A. Improved strategies for sequence-independent amplification and sequencing of viral double-stranded RNA genomes. J. Gen. Virol. 2009, 90, 1423–1432. [Google Scholar] [CrossRef]
- Darissa, O.; Willingmann, P.; Adam, G. Optimized approaches for the sequence determination of double-stranded RNA templates. J. Virol. Methods 2010, 169, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Datta, A. DotKnot: Pseudoknot prediction using the probability dot plot under a refined energy model. Nucleic Acids Res. 2010, 38, e103. [Google Scholar] [CrossRef] [PubMed]
- Byun, Y.; Han, K. PseudoViewer3: Generating planar drawings of large-scale RNA structures with pseudoknots. Bioinformatics 2009, 25, 1435–1437. [Google Scholar] [CrossRef]
- Löytynoja, A.; Vilella, J.A.; Goldman, N. Accurate extension of multiple sequence alignments using a phylogeny-aware graph algorithm. Bioinformatics 2012, 28, 1684–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Posada, D.; Kozlov, M.A.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, G. Estimating the Dimension of a Model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Le, S.Q. and Gascuel, O. An Improved General Amino Acid Replacement Matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, M.A.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Cole, T.E.; Brasier, C.M.; Buck, K.W. Novel structures of two virus-like RNA elements from a diseased isolate of the Dutch elm disease fungus. Ophiostoma Novo-Ulmi. Virol. 1998, 242, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor: New York, NY, USA, 1989. [Google Scholar]
- Guo, L.H.; Sun, L.; Chiba, S.; Araki, H.; Suzuki, N. Coupled termination/reinitiation for translation of the downstream open reading frame B of the prototypic hypovirus CHV1-EP713. Nucleic Acids Res. 2009, 37, 3645–3659. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S.; Lin, Y.H.; Kondo, H.; Kanematsu, S.; Suzuki, N. A novel victorivirus from a phytopathogenic fungus, Rosellinia necatrix, is infectious as particles and targeted by RNA silencing. J. Virol. 2013, 87, 6727–6738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, N.; Dueñas, E.; Quesada-Moraga, E.; Zabalgogeazcoa, I. Prevalence and diversity of viruses in the entomopathogenic fungus Beauveria bassiana. Appl. Env. Microbiol. 2012, 78, 8523–8530. [Google Scholar] [CrossRef] [Green Version]
- Mu, F.; Li, B.; Cheng, S.; Jia, J.; Jiang, D.; Fu, Y.; Cheng, J.; Lin, Y.; Chen, T.; Xie, J. Nine viruses from eight lineages exhibiting new evolutionary modes that co-infect a hypovirulent phytopathogenic fungus. PLoS Pathog. 2021, 17, e1009823. [Google Scholar] [CrossRef]
- Khalifa, M.E.; MacDiarmid, R.M. A Novel Totivirus Naturally Occurring in Two Different Fungal Genera. Front. Microbiol. 2019, 10, 2318. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Havens, W.M.; Nibert, M.L.; Ghabrial, S.A. RNA sequence determinants of a coupled termination-reinitiation strategy for downstream open reading frame translation in Helminthosporium victoriae virus 190S and other victoriviruses (family Totiviridae). J. Virol. 2011, 85, 7343–7352. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Havens, W.M.; Nibert, M.L.; Ghabrial, S.A. An RNA cassette from Helminthosporium victoriae virus 190S necessary and sufficient for stop/restart translation. Virology 2015, 474, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Yie, S.W.; Khalifa, M.E.; Hahn, T.; Pearson, M.N. Molecular characterization of a novel victorivirus from the entomopathogenic fungus Beauveria bassiana. Arch. Virol. 2014, 159, 1321–1327. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Zhang, T.; Zhu, Y.; Yuan, Y.; Wang, S.; Liu, J.; Liu, D. Isolation, and characterization of a novel mycovirus from Penicillium digitatum. Virology 2016, 494, 15–22. [Google Scholar] [CrossRef]
- Jamal, A.; Sato, Y.; Shahi, S.; Shamsi, W.; Kondo, H.; Suzuki, N. Novel victorivirus from a Pakistani isolate of Alternaria alternata lacking a typical translational stop/restart sequence signature. Viruses 2019, 11, 577. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Xia, Y.; Zhang, H.; Zhang, X.; Chen, H. A Victorivirus from Fusarium asiaticum, the pathogen of Fusarium head blight in China. Arch. Virol. 2019, 164, 313–316. [Google Scholar] [CrossRef]
- Ghabrial, S. Totiviruses. In Encyclopedia of Virology, 3rd ed.; Mahy, B.W.J., Van Regenmortel, M.H.V., Eds.; Elsevier: Oxford, UK, 2008; pp. 163–174. [Google Scholar]
- Atkins, J.F.; Loughran, G.; Bhatt, P.R.; Firth, A.E.; Baranov, P.V. Ribosomal frameshifting, and transcriptional slippage: From genetic steganography and cryptography to adventitious use. Nucleic. Acids Res. 2016, 44, 7007–7078. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.E.; Suh, J.M.; Scheffter, S.; Patterson, J.L.; Chung, I.K. Identification of a ribosomal frameshift in Leishmania RNA virus 1–4. J. Biochem. 1996, 120, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Goodman, R.P.; Ghabrial, S.A.; Fichorova, R.N.; Nibert, M.L. Trichomonasvirus: A new genus of protozoan viruses in the family Totiviridae. Arch. Virol. 2011, 156, 171–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Collection Number | Other Collection Number | Origin/Other Information | dsRNA Size (kb) | Detected Viruses |
|---|---|---|---|---|---|
| Mucor hiemalis f. hiemalis | WRL CN(M) 122 a | SZMC 12,034 b | Oak-bark, UK | 5.5 | MhV1a e |
| Mucor hiemalis f. hiemalis | NRRL 3624 c | CBS 201.65 d | USA, Type strain of M. hiemalis | 5.5 | MhV1b f |
| Mucor hiemalis f. corticola | NRRL 3617 | CBS 366.68 | soil, Austria | 5.2 | MhV2 g |
| Mucor hiemalis f. corticola | NRRL 3616 | CBS 365.58 | soil, Austria | 5.0; 4.6; 3.5 | MhV3 h; MhV4 i |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kartali, T.; Nyilasi, I.; Kocsubé, S.; Patai, R.; Polgár, T.F.; Zsindely, N.; Nagy, G.; Bodai, L.; Lipinszki, Z.; Vágvölgyi, C.; et al. Characterization of Four Novel dsRNA Viruses Isolated from Mucor hiemalis Strains. Viruses 2021, 13, 2319. https://doi.org/10.3390/v13112319
Kartali T, Nyilasi I, Kocsubé S, Patai R, Polgár TF, Zsindely N, Nagy G, Bodai L, Lipinszki Z, Vágvölgyi C, et al. Characterization of Four Novel dsRNA Viruses Isolated from Mucor hiemalis Strains. Viruses. 2021; 13(11):2319. https://doi.org/10.3390/v13112319
Chicago/Turabian StyleKartali, Tünde, Ildikó Nyilasi, Sándor Kocsubé, Roland Patai, Tamás F. Polgár, Nóra Zsindely, Gábor Nagy, László Bodai, Zoltán Lipinszki, Csaba Vágvölgyi, and et al. 2021. "Characterization of Four Novel dsRNA Viruses Isolated from Mucor hiemalis Strains" Viruses 13, no. 11: 2319. https://doi.org/10.3390/v13112319
APA StyleKartali, T., Nyilasi, I., Kocsubé, S., Patai, R., Polgár, T. F., Zsindely, N., Nagy, G., Bodai, L., Lipinszki, Z., Vágvölgyi, C., & Papp, T. (2021). Characterization of Four Novel dsRNA Viruses Isolated from Mucor hiemalis Strains. Viruses, 13(11), 2319. https://doi.org/10.3390/v13112319