
Cytomegalovirus Immunity, Inflammation and Cognitive Abilities in the Elderly


Abstract
:1. Introduction
2. Materials and Methods
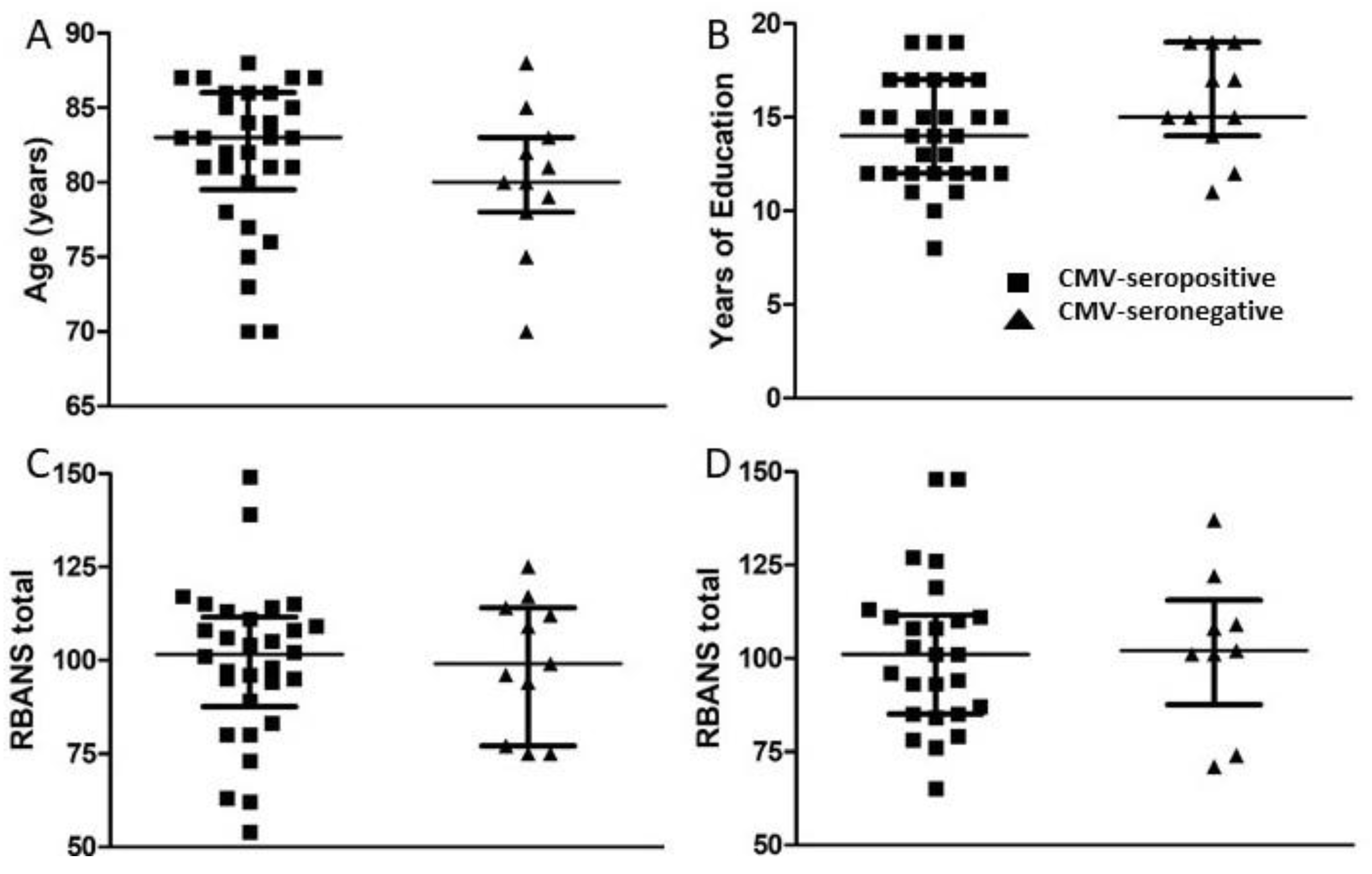
2.1. Study Subjects and Sample Processing
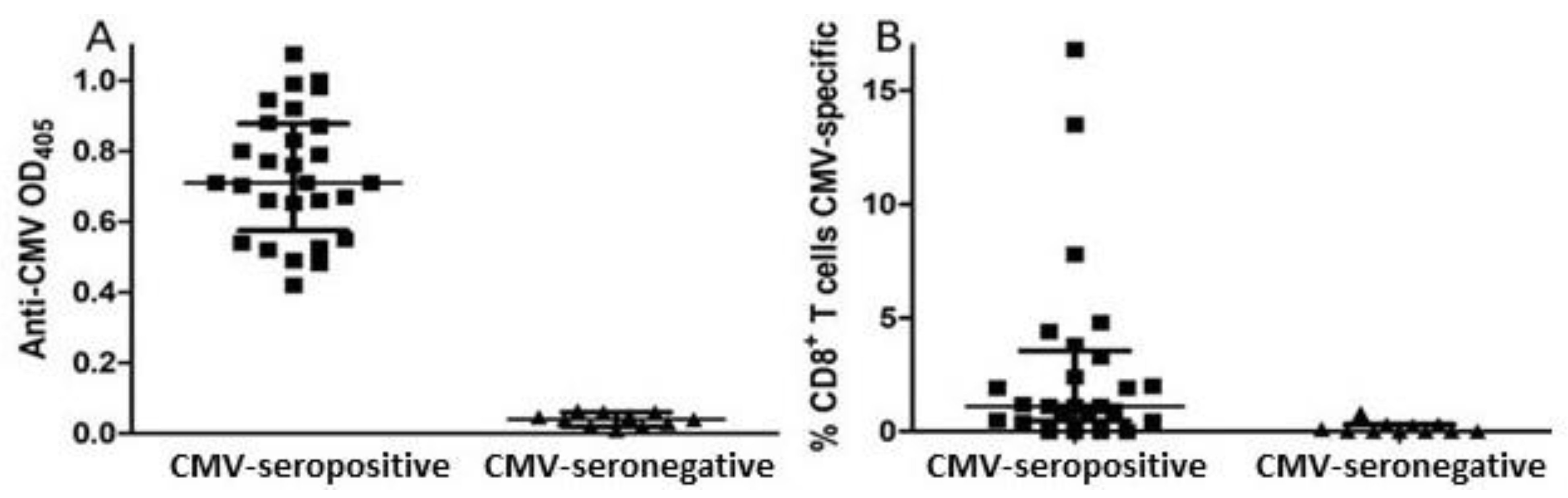
2.2. Anti-CMV IgG ELISA
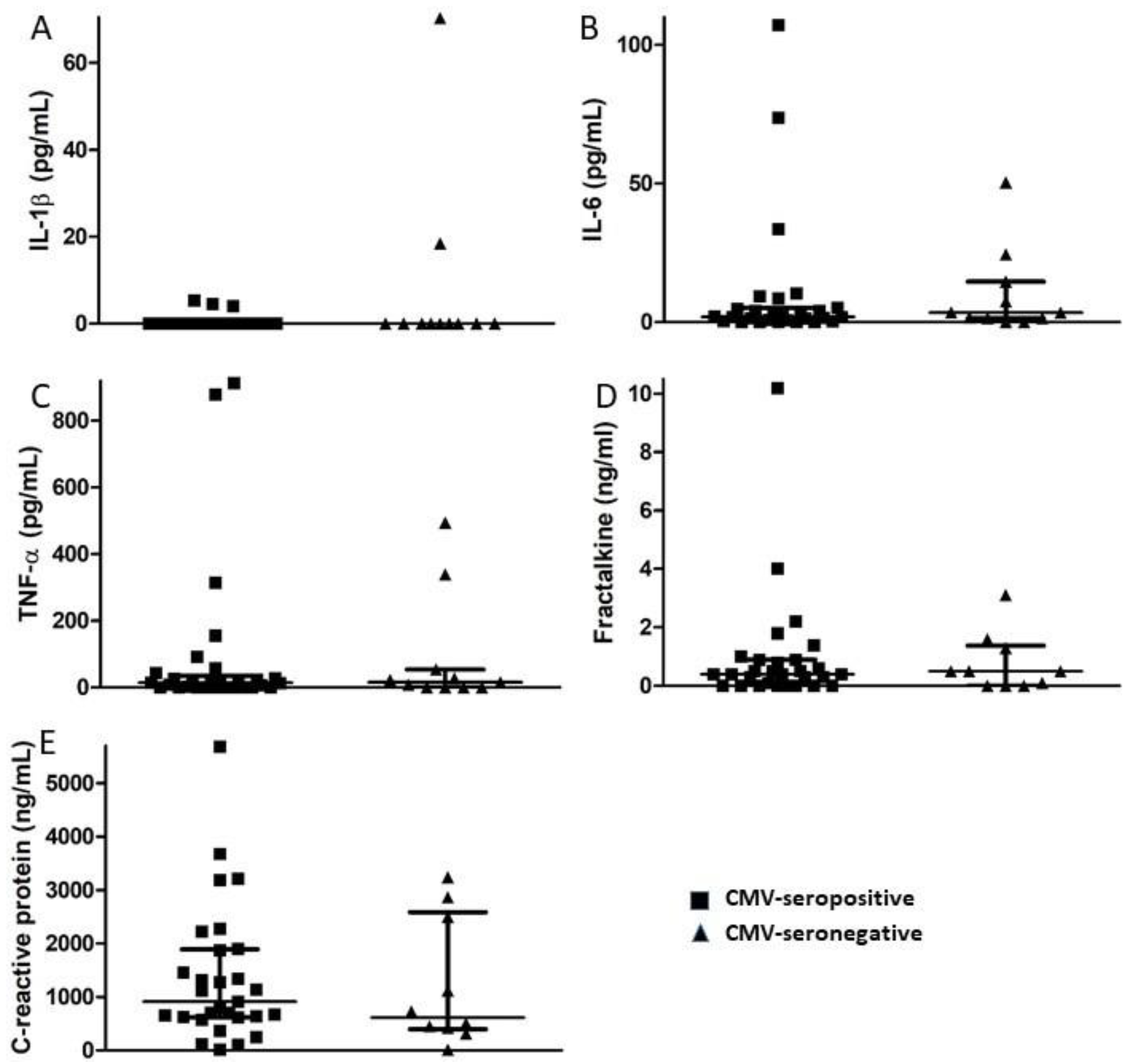
2.3. Measurement of Inflammatory Markers
2.4. Measurement of CMV-Specific CD8+ T Cell Responses
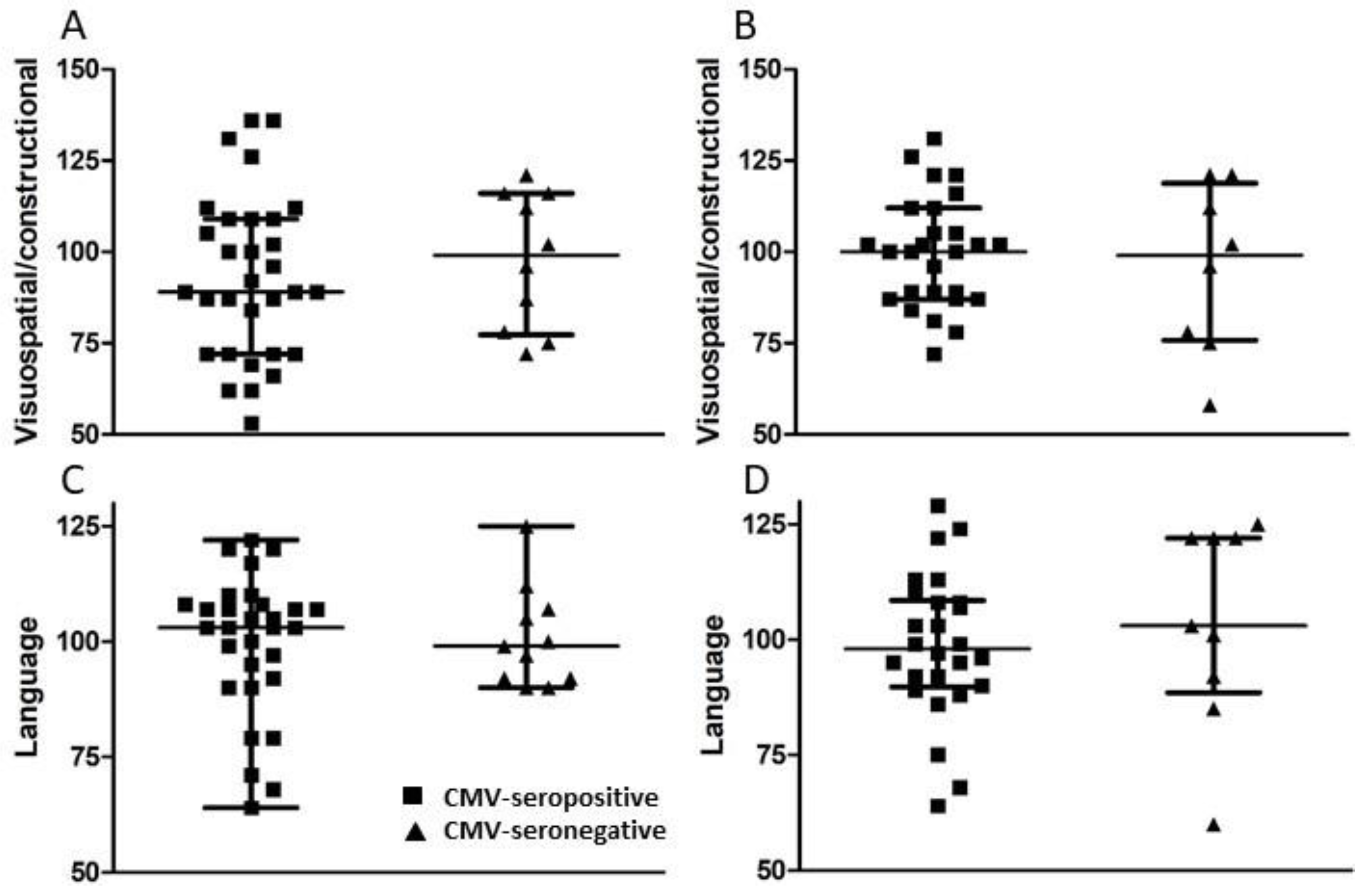
2.5. Neuropsychological Testing
2.6. Mood
2.7. Statistical Analysis
3. Results
3.1. Study Subjects
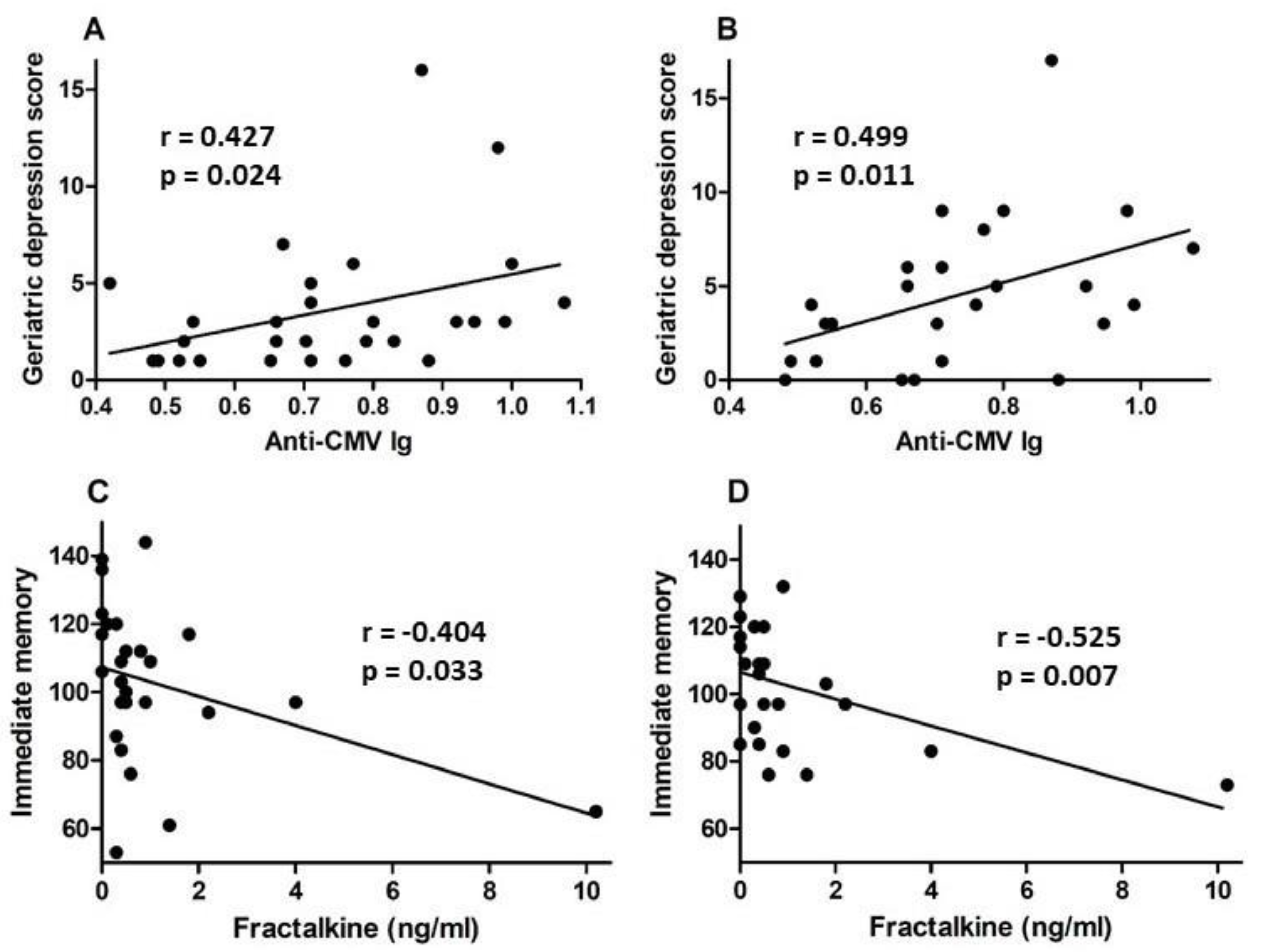
3.2. Plasma Markers of Inflammation
3.3. Psychological Testing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kanasi, E.; Ayilavarapu, S.; Jones, J. The aging population: Demographics and the biology of aging. Periodontology 2000 2016, 72, 13–18. [Google Scholar] [CrossRef]
- Hazra, N.C.; Rudisill, C.; Gulliford, M. Determinants of health care costs in the senior elderly: Age, comorbidity, impairment, or proximity to death? Eur. J. Heal. Econ. 2017, 19, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Borson, S.; Bartels, S.J.; Colenda, C.C.; Gottlieb, G.L.; Meyers, B. Geriatric mental health services research: Strategic Plan for an Aging Population: Report of the Health Services Work Group of the American Association for Geriatric Psychiatry. Am. J. Geriatr. Psychiatry 2001, 9, 191–204. [Google Scholar] [CrossRef]
- Baylis, D.; Bartlett, D.B.; Patel, H.P.; Roberts, H.C. Understanding how we age: Insights into inflammaging. Longev. Heal. 2013, 2, 1–8. [Google Scholar] [CrossRef]
- Howcroft, T.K.; Campisi, J.; Louis, G.B.; Smith, M.T.; Wise, B.; Wyss-Coray, T.; Augustine, A.D.; McElhaney, J.E.; Kohanski, R.; Sierra, F. The role of inflammation in age-related disease. Aging 2013, 5, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, D.J.; Wijshake, T.; Tchkonia, T.; Lebrasseur, N.K.; Childs, B.G.; Van De Sluis, B.; Kirkland, J.L.; Van Deursen, J.M. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature 2011, 479, 232–236. [Google Scholar] [CrossRef]
- Naylor, R.M.; Baker, D.J.; Van Deursen, J.M. Senescent Cells: A Novel Therapeutic Target for Aging and Age-Related Diseases. Clin. Pharmacol. Ther. 2012, 93, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Coppé, J.-P.; Desprez, P.-Y.; Krtolica, A.; Campisi, J. The Senescence-Associated Secretory Phenotype: The Dark Side of Tumor Suppression. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 99–118. [Google Scholar] [CrossRef] [Green Version]
- Staras, S.A.S.; Dollard, S.C.; Radford, K.W.; Flanders, W.D.; Pass, R.; Cannon, M.J. Seroprevalence of Cytomegalovirus Infection in the United States, 1988-1994. Clin. Infect. Dis. 2006, 43, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Emery, V.C.; Cope, A.V.; Bowen, E.F.; Gor, D.; Griffiths, P.D. The Dynamics of Human Cytomegalovirus Replication in Vivo. J. Exp. Med. 1999, 190, 177–182. [Google Scholar] [CrossRef]
- van de Berg, P.J.; van Stijn, A.; Ten Berge, I.J.; van Lier, R.A. A fingerprint left by cytomegalovirus infection in the human T cell compartment. J. Clin. Virol. 2008, 41, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Wikby, A.; Nilsson, B.O.; Forsey, R.; Thompson, J.; Strindhall, J.; Lofgren, S.; Ernerudh, J.; Ernerudh, G.; Ferguson, F.; Johansson, V. The immune risk phenotype is associated with IL-6 in the terminal decline stage: Findings from the Swedish NONA immune longitudinal study of very late life func-tioning. Mech. Ageing Dev. 2006, 127, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Strindhall, J.; Nilsson, B.O.; Löfgren, S.; Ernerudh, J.; Pawelec, G.; Johansson, B.; Wikby, A. No Immune Risk Profile among indi-viduals who reach 100 years of age: Findings from the Swedish NONA immune longitudinal study. Exp. Gerontol. 2007, 42, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Hadrup, S.R.; Strindhall, J.; Kollgaard, T.; Seremet, T.; Johansson, B.; Pawelec, G.; Johansson, B.; Wikby, A. Longitudinal studies of clonally ex-panded CD8 T cells reveal a repertoire shrinkage predicting mortality and an increased number of dysfunctional cytomeg-alovirus-specific T cells in the very elderly. J. Immunol. 2006, 176, 2645–2653. [Google Scholar] [CrossRef] [Green Version]
- van de Berg, P.J.; Heutinck, K.M.; Raabe, R.; Minnee, R.C.; Young, S.L.; van Donselaar-van der Pant, K.A.; Bemelman, F.J.; van Lier Rene, A.; ten Berge, I.J. Human cyto-megalovirus induces systemic immune activation characterized by a type 1 cytokine signature. J. Infect. Dis. 2010, 202, 690–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaes, B.; Pasquet, A.; Wallemacq, P.; Rezzoug, N.; Mekouar, H.; Olivier, P.A.; Legrand, D.; Mathei, C.; Van Pottelbergh, G.; Degryse, J. The BELFRAIL (BFC80+) study: A population-based prospective cohort study of the very elderly in Belgium. BMC Geriatr. 2010, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Roberts, E.T.; Haan, M.N.; Dowd, J.; Aiello, A.E. Cytomegalovirus Antibody Levels, Inflammation, and Mortality Among Elderly Latinos Over 9 Years of Follow-up. Am. J. Epidemiology 2010, 172, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Savva, G.M.; Pachnio, A.; Kaul, B.; Morgan, K.; Huppert, F.A.; Brayne, C.; Moss, P.A.H. The Medical Research Council Cognitive Function and Ageing Study Cytomegalovirus infection is associated with increased mortality in the older population. Aging Cell 2013, 12, 381–387. [Google Scholar] [CrossRef]
- Spyridopoulos, I.; Martin-Ruiz, C.; Hilkens, C.; Yadegarfar, M.E.; Isaacs, J.; Jagger, C.; Kirkwood, T.B.L.; Von Zglinicki, T. CMV seropositivity and T-cell senescence predict increased cardiovascular mortality in octogenarians: Results from the Newcastle 85+ study. Aging Cell 2015, 15, 389–392. [Google Scholar] [CrossRef] [Green Version]
- Vescovini, R.; Biasini, C.; Telera, A.R.; Basaglia, M.; Stella, A.; Magalini, F.; Bucci, L.; Monti, D.; Lazzarotto, T.; Monte, P.D.; et al. Intense Antiextracellular Adaptive Immune Response to Human Cytomegalovirus in Very Old Subjects with Impaired Health and Cognitive and Functional Status. J. Immunol. 2010, 184, 3242–3249. [Google Scholar] [CrossRef] [Green Version]
- Dickerson, F.; Stallings, C.; Origoni, A.; Katsafanas, E.; Schweinfurth, L.A.; Savage, C.L.; Yolken, R. Association between cytomegalovirus antibody levels and cognitive functioning in non-elderly adults. PLoS ONE 2014, 9, e95510. [Google Scholar] [CrossRef]
- Krogh, J.; Benros, M.E.; Jorgensen, M.B.; Vesterager, L.; Elfving, B.; Nordentoft, M. The association between depressive symptoms, cognitive function, and inflammation in major depression. Brain Behav Immun. 2014, 35, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Heath, J.; Newhook, N.; Comeau, E.; Gallant, M.; Fudge, N.; Grant, M. NKG2C+CD57+ Natural Killer Cell Expansion Parallels Cytomegalovirus-Specific CD8+ T Cell Evolution towards Senescence. J. Immunol. Res. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randolph, C.; Tierney, M.C.; Mohr, E.; Chase, T.N. The Repeatable Battery for the Assessment of Neuropsychological Status (RBANS): Preliminary Clinical Validity. J. Clin. Exp. Neuropsychol. 1998, 20, 310–319. [Google Scholar] [CrossRef]
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated dis-eases. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69 (Suppl. 1), S4–S9. [Google Scholar] [CrossRef]
- Limatola, C.; Lauro, C.; Catalano, M.; Ciotti, M.T.; Bertollini, C.; Di Angelantonio, S.; Ragozzino, D.; Eusebi, F. Chemokine CX3CL1 protects rat hippocampal neurons against glutamate-mediated excitotoxicity. J. Neuroimmunol. 2005, 166, 19–28. [Google Scholar] [CrossRef]
- Beisser, P.S.; Laurent, L.; Virelizier, J.-L.; Michelson, S. Human Cytomegalovirus Chemokine Receptor Gene US28 Is Transcribed in Latently Infected THP-1 Monocytes. J. Virol. 2001, 75, 5949–5957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finneran, D.J.; Nash, K.R. Neuroinflammation and fractalkine signaling in Alzheimer’s disease. J. Neuroinflammation 2019, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafe, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Vasto, S.; Candore, G.; Balistreri, C.R.; Caruso, M.; Colonna-Romano, G.; Grimaldi, M.P.; Listi, F.; Nuzzo, D.; Lio, D.; Caruso, C. Inflammatory networks in ageing, age-related diseases and longevity. Mech. Ageing Dev. 2007, 128, 83–91. [Google Scholar] [CrossRef]
- Letendre, S.; Bharti, A.; Perez-Valero, I.; Hanson, B.; Franklin, D.; Woods, S.P.; Gianella, S.; Faria de Oliveira, M.; Heaton, R.K.; Grant, I.; et al. Higher Anti-Cytomegalovirus Immunoglobulin G Concentrations Are Associated With Worse Neurocognitive Performance During Suppressive Antiretroviral Therapy. Clin. Infect. Dis. 2018, 67, 770–777. [Google Scholar] [CrossRef]
- Ballegaard, V.; Pedersen, K.K.; Pedersen, M.; Braendstrup, P.; Kirkby, N.; Buus, A.S.; Ryder, L.P.; Gerstoft, J.; Nielsen, S.D. Cytomegalovirus-Specific CD4+ T-cell Responses and CMV-IgG Levels Are Associated With Neurocognitive Impairment in People Living With HIV. J. Acquir. Immune Defic. Syndr. 2018, 79, 117–125. [Google Scholar] [CrossRef]
- Naeger, D.; Martin, J.N.; Sinclair, E.; Hunt, P.W.; Bangsberg, D.R.; Hecht, F.; Hsue, P.; McCune, J.M.; Deeks, S.G. Cytomegalovirus-Specific T Cells Persist at Very High Levels during Long-Term Antiretroviral Treatment of HIV Disease. PLoS ONE 2010, 5, e8886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, M.L.; Mudd, J.C.; Shive, C.L.; Younes, S.-A.; Panigrahi, S.; Sieg, S.F.; Lee, S.A.; Hunt, P.W.; Calabrese, L.H.; Gianella, S.; et al. CD8 T-Cell Expansion and Inflammation Linked to CMV Coinfection in ART-treated HIV Infection. Clin. Infect. Dis. 2015, 62, 392–396. [Google Scholar] [CrossRef]
- Barrett, L.; Fudge, N.J.; Heath, J.J.; Grant, M.D. Cytomegalovirus Immunity and Exhaustive CD8+ T Cell Proliferation in Treated Human Immunodeficiency Virus Infection. Clin. Infect. Dis. 2016, 62, 1467–1468. [Google Scholar] [CrossRef] [Green Version]
- Donzis, E.J.; Tronson, N.C. Modulation of learning and memory by cytokines: Signaling mechanisms and long term consequences. Neurobiol. Learn. Mem. 2014, 115, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Bradburn, S.; Sarginson, J.; Murgatroyd, C. Association of Peripheral Interleukin-6 with Global Cognitive Decline in Non-demented Adults: A Meta-Analysis of Prospective Studies. Front. Aging Neurosci. 2018, 9, 438. [Google Scholar] [CrossRef]
- Shah, P.J.; Ebmeier, K.P.; Glabus, M.F.; Goodwin, G.M. Cortical grey matter reductions associated with treatment-resistant chronic unipolar depression. Br. J. Psychiatry 1998, 172, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Stertz, L.; Magalhães, P.V.; Kapczinski, F. Is bipolar disorder an inflammatory condition? The relevance of microglial activation. Curr. Opin. Psychiatry 2013, 26, 19–26. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant ID | Age at Entry | Years of Education | Sex | CMV Status |
|---|---|---|---|---|
| 3001 | 83 | 12 | Female | Seronegative |
| 3002 | 87 | 10 | Female | Seropositive |
| 3003 | 84 | 12 | Male | Seropositive |
| 3004 | 83 | 14 | Female | Seropositive |
| 3005 | 81 | 15 | Male | Seronegative |
| 3006 | 88 | 12 | Male | Seropositive |
| 3007 | 80 | 17 | Female | Seropositive |
| 3008 | 81 | 15 | Male | Seropositive |
| 3009 | 83 | 12 | Female | Seropositive |
| 3010 | 87 | 12 | Female | Seropositive |
| 3011 * | 82 | 11 | Female | Seronegative |
| 3012 | 87 | 8 | Female | Seropositive |
| 3013 | 85 | 14 | Female | Seropositive |
| 3014 | 81 | 19 | Male | Seropositive |
| 3015 | 84 | 15 | Female | Seropositive |
| 3016 * | 83 | 11 | Male | Seropositive |
| 3017 | 85 | 19 | Male | Seropositive |
| 3018 * | 88 | 15 | Male | Seronegative |
| 3019 | 80 | 15 | Female | Seronegative |
| 3020 * | 87 | 14 | Female | Seropositive |
| 3021 | 86 | 17 | Female | Seropositive |
| 3022 | 85 | 14 | Female | Seronegative |
| 3023 | 86 | 12 | Female | Seropositive |
| 3024 | 82 | 15 | Male | Seropositive |
| 3025 * | 86 | 11 | Female | Seropositive |
| 3026 | 83 | 15 | Male | Seropositive |
| 3027 | 78 | 19 | Male | Seropositive |
| 3028 | 75 | 17 | Female | Seronegative |
| 3029 | 70 | 17 | Female | Seronegative |
| 3030 | 78 | 19 | Male | Seronegative |
| 3031 | 80 | 19 | Male | Seronegative |
| 3032 | 82 | 17 | Female | Seropositive |
| 3033 | 79 | 19 | Male | Seronegative |
| 3034 | 73 | 15 | Female | Seropositive |
| 3035 | 76 | 17 | Male | Seropositive |
| 3036 | 75 | 13 | Female | Seropositive |
| 3037 | 81 | 12 | Female | Seropositive |
| 3038 | 70 | 12 | Female | Seropositive |
| 3039 | 70 | 17 | Female | Seropositive |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hesson, J.; Fudge, N.; Grant, M. Cytomegalovirus Immunity, Inflammation and Cognitive Abilities in the Elderly. Viruses 2021, 13, 2321. https://doi.org/10.3390/v13112321
Hesson J, Fudge N, Grant M. Cytomegalovirus Immunity, Inflammation and Cognitive Abilities in the Elderly. Viruses. 2021; 13(11):2321. https://doi.org/10.3390/v13112321
Chicago/Turabian StyleHesson, Jacqueline, Neva Fudge, and Michael Grant. 2021. "Cytomegalovirus Immunity, Inflammation and Cognitive Abilities in the Elderly" Viruses 13, no. 11: 2321. https://doi.org/10.3390/v13112321
APA StyleHesson, J., Fudge, N., & Grant, M. (2021). Cytomegalovirus Immunity, Inflammation and Cognitive Abilities in the Elderly. Viruses, 13(11), 2321. https://doi.org/10.3390/v13112321