
Identification of African Swine Fever Virus Transcription within Peripheral Blood Mononuclear Cells of Acutely Infected Pigs

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pigs
2.2. Challenge Virus
2.3. Study Design
2.4. Clinical Examination and Euthanasia
2.5. Sampling from the Inoculated Pigs
2.6. PBMC Isolation and Processing
2.7. RNA Purification
2.8. ASFV DNA Detection by Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.9. Data Analysis
Mapping of Sequence Reads to the Pig Genome and to the ASFV Genome
3. Results
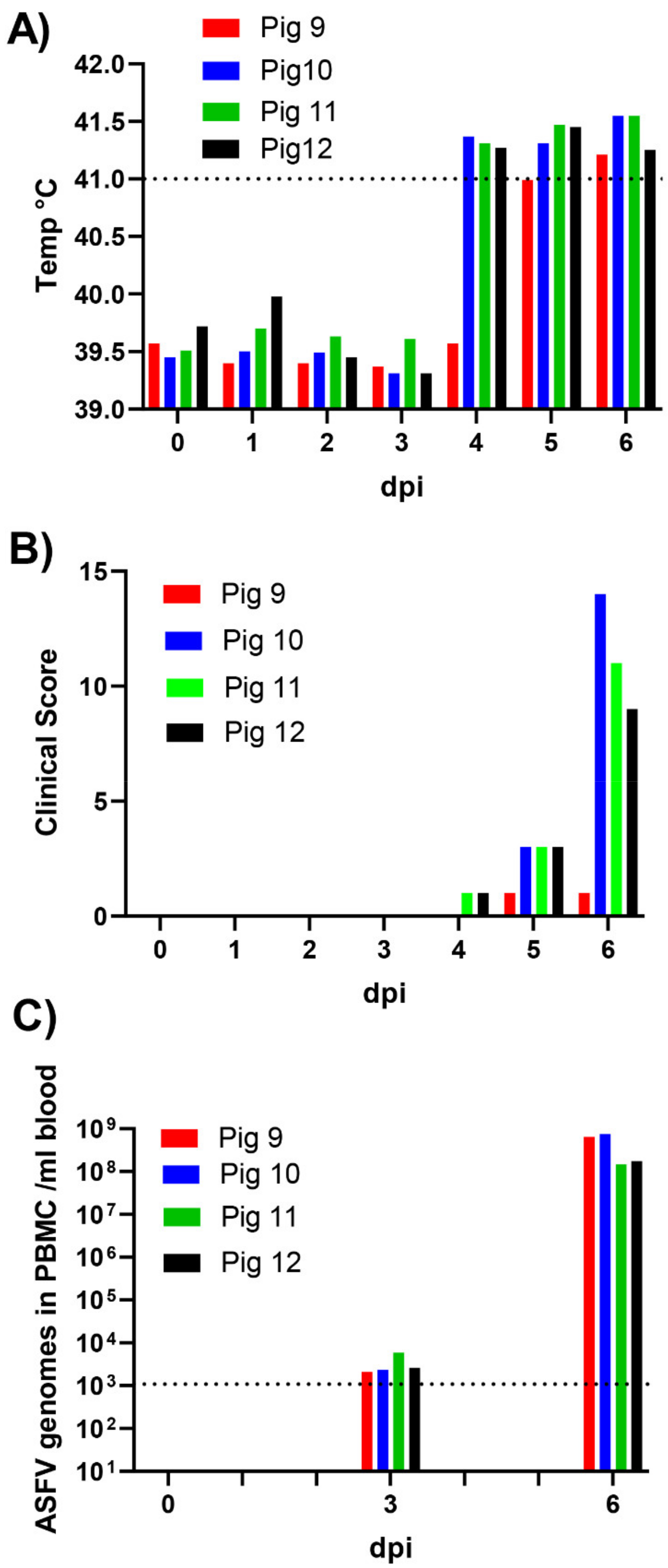
3.1. Course of Infection in the Inoculated Pigs
3.2. Virus Derived RNA Transcript Analysis from PBMCs of ASFV-Inoculated Pigs
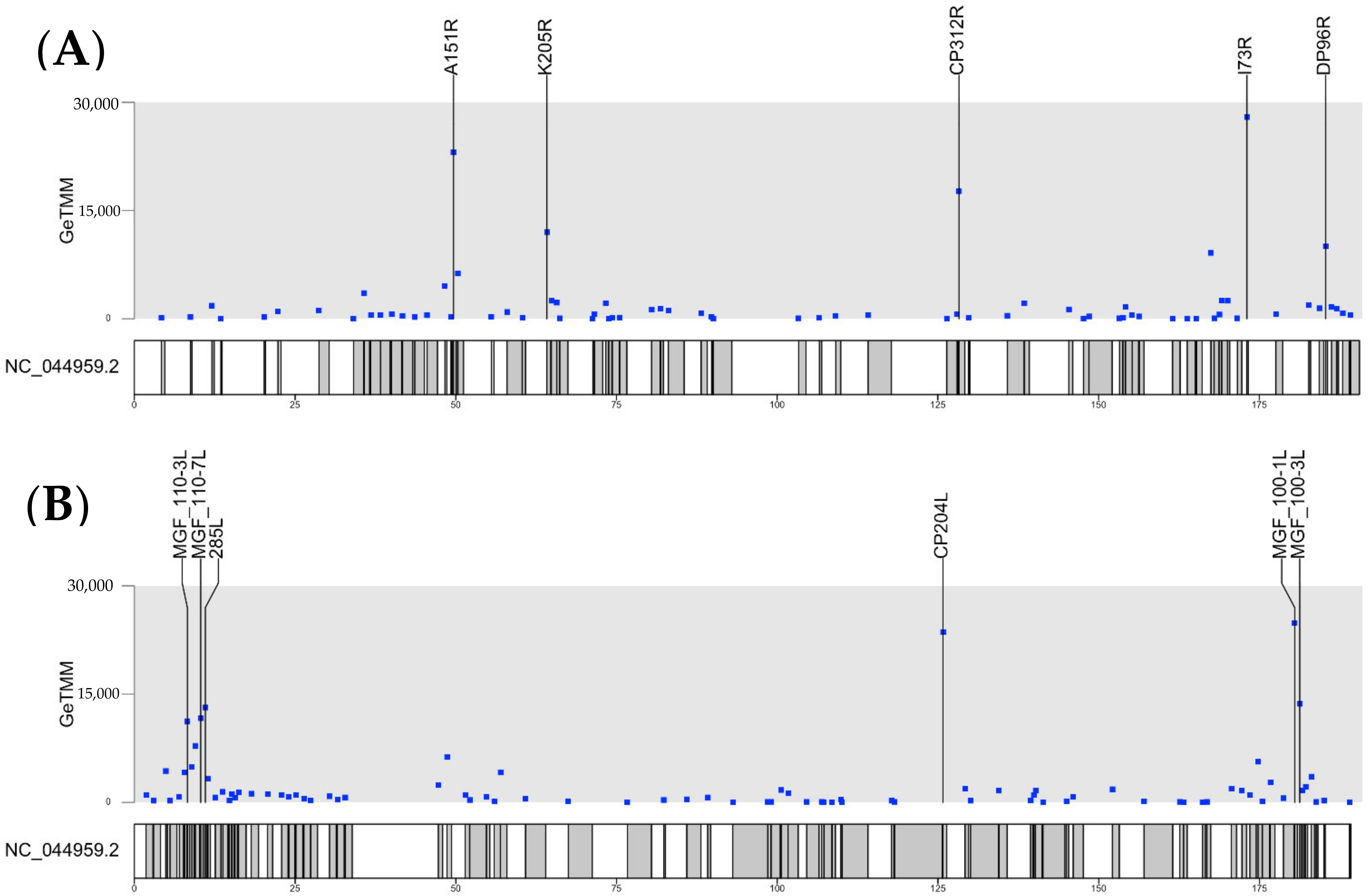
3.3. Transcription of Individual ASFV Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Understanding and Combatting African Swine Fever. 2021, pp. 25–61. Available online: https://agritrop.cirad.fr/598144/7/ID598144.pdf (accessed on 26 October 2021).
- Dixon, L.; Sun, H.; Roberts, H. African swine fever. Antivir. Res. 2019, 165, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Olesen, A.S.; Belsham, G.J.; Rasmussen, T.B.; Lohse, L.; Bødker, R.; Halasa, T.; Boklund, A.; Bøtner, A. Potential routes for indirect transmission of African swine fever virus into domestic pig herds. Transbound. Emerg. Dis. 2020, 67, 1472–1484. [Google Scholar] [CrossRef]
- Achenbach, J.E.; Gallardo, C.; Pelegrin, E.N.; Rivera-Arroyo, B.; Degefa-Negi, T.; Arias, M.; Jenberie, S.; Mulisa, D.D.; Gizaw, D.; Gelaye, E.; et al. Identification of a New Genotype of African Swine Fever Virus in Domestic Pigs from Ethiopia. Transbound. Emerg. Dis. 2017, 64, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Bastos, A.D.S.; Penrith, M.-L.; Crucière, C.; Edrich, J.L.; Hutchings, G.; Roger, F.; Couacy-Hymann, E.; Thomson, G.R. Genotyping field strains of African swine fever virus by partial p72 gene characterisation. Arch. Virol. 2003, 148, 693–706. [Google Scholar] [CrossRef] [PubMed]
- Boshoff, C.; Bastos, A.; Gerber, L.; Vosloo, W. Genetic characterisation of African swine fever viruses from outbreaks in southern Africa (1973–1999). Vet. Microbiol. 2007, 121, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Lubisi, B.A.; Bastos, A.; Dwarka, R.M.; Vosloo, W. Molecular epidemiology of African swine fever in East Africa. Arch. Virol. 2005, 150, 2439–2452. [Google Scholar] [CrossRef]
- Quembo, C.J.; Jori, F.; Vosloo, W.; Heath, L. Genetic characterization of African swine fever virus isolates from soft ticks at the wildlife/domestic interface in Mozambique and identification of a novel genotype. Transbound. Emerg. Dis. 2018, 65, 420–431. [Google Scholar] [CrossRef] [Green Version]
- Costard, S.; Wieland, B.; De Glanville, W.; Jori, F.; Rowlands, R.; Vosloo, W.; Roger, F.; Pfeiffer, D.; Dixon, L.K. African swine fever: How can global spread be prevented? Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2683–2696. [Google Scholar] [CrossRef] [Green Version]
- OIE WAHIS Interface. 2021. Available online: https://wahis.oie.int/#/dashboards/country-or-disease-dashboard (accessed on 19 August 2021).
- FAO. ASF Situation in Asia Update. Available online: http://www.fao.org/ag/againfo/programmes/en/empres/ASF/situation_update.html (accessed on 1 June 2020).
- Busch, F.; Haumont, C.; Penrith, M.-L.; Laddomada, A.; Dietze, K.; Globig, A.; Guberti, V.; Zani, L.; Depner, K. Evidence-Based African Swine Fever Policies: Do We Address Virus and Host Adequately? Front. Vet. Sci. 2021, 8, 224. [Google Scholar] [CrossRef]
- Sánchez-Cordón, P.J.; Montoya, M.; Reis, A.L.; Dixon, L.K. African swine fever: A re-emerging viral disease threatening the global pig industry. Vet. J. 2018, 233, 41–48. [Google Scholar] [CrossRef]
- Gallardo, C.; Soler, A.; Nieto, R.; Cano, C.; Pelayo, V.; Sánchez, M.A.; Pridotkas, G.; Fernandez-Pinero, J.; Briones, V.; Arias, M. Experimental Infection of Domestic Pigs with African Swine Fever Virus Lithuania 2014 Genotype II Field Isolate. Transbound. Emerg. Dis. 2015, 64, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Guinat, C.; Reis, A.L.; Netherton, C.L.; Goatley, L.; Pfeiffer, D.U.; Dixon, L. Dynamics of African swine fever virus shedding and excretion in domestic pigs infected by intramuscular inoculation and contact transmission. Vet. Res. 2014, 45, 1–9. [Google Scholar] [CrossRef]
- Olesen, A.S.; Lohse, L.; Boklund, A.; Halasa, T.; Gallardo, C.; Pejsak, Z.; Belsham, G.; Rasmussen, T.B.; Bøtner, A. Transmission of African swine fever virus from infected pigs by direct contact and aerosol routes. Vet. Microbiol. 2017, 211, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Dixon, L.K.; Chapman, D.A.; Netherton, C.L.; Upton, C. African swine fever virus replication and genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Cackett, G.; Matelska, D.; Sýkora, M.; Portugal, R.; Malecki, M.; Bähler, J.; Dixon, L.; Werner, F. The African Swine Fever Virus Transcriptome. J. Virol. 2020, 94, e00119-20. [Google Scholar] [CrossRef] [Green Version]
- Jaing, C.; Rowland, R.R.R.; Allen, J.; Certoma, A.; Thissen, J.B.; Bingham, J.; Rowe, B.; White, J.; Wynne, J.; Johnson, D.; et al. Gene expression analysis of whole blood RNA from pigs infected with low and high pathogenic African swine fever viruses. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Zhu, J.J.; Ramanathan, P.; Bishop, E.A.; O’Donnell, V.; Gladue, D.P.; Borca, M.V. Mechanisms of African swine fever virus pathogenesis and immune evasion inferred from gene expression changes in infected swine macrophages. PLoS ONE 2019, 14, e0223955. [Google Scholar] [CrossRef]
- Ju, X.; Li, F.; Li, J.; Wu, C.; Xiang, G.; Zhao, X.; Nan, Y.; Zhao, D.; Ding, Q. Genome-wide transcriptomic analysis of highly virulent African swine fever virus infection reveals complex and unique virus host interaction. Vet. Microbiol. 2021, 261, 109211. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Villamandos, J.; Bautista, M.; Cordón, P.S.; Carrasco, L. Pathology of African swine fever: The role of monocyte-macrophage. Virus Res. 2013, 173, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Olesen, A.S.; Lohse, L.; Dalgaard, M.D.; Woźniakowski, G.; Belsham, G.J.; Bøtner, A.; Rasmussen, T.B. Complete genome sequence of an African swine fever virus (ASFV POL/2015/Podlaskie) determined directly from pig erythrocyte-associated nucleic acid. J. Virol. Methods 2018, 261, 14–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forth, J.H.; Forth, L.F.; King, J.; Groza, O.; Hübner, A.; Olesen, A.S.; Höper, D.; Dixon, L.K.; Netherton, C.L.; Rasmussen, T.B.; et al. A Deep-Sequencing Workflow for the Fast and Efficient Generation of High-Quality African Swine Fever Virus Whole-Genome Sequences. Viruses 2019, 11, 846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tignon, M.; Gallardo, C.; Iscaro, C.; Hutet, E.; Van der Stede, Y.; Kolbasov, D.; De Mia, G.M.; Le Potier, M.-F.; Bishop, R.P.; Arias, M.; et al. Development and inter-laboratory validation study of an improved new real-time PCR assay with internal control for detection and laboratory diagnosis of African swine fever virus. J. Virol. Methods 2011, 178, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Warr, A.; Affara, N.; Aken, B.; Beiki, H.; Bickhart, D.M.; Billis, K.; Chow, W.; Eory, L.; Finlayson, H.A.; Flicek, P.; et al. An improved pig reference genome sequence to enable pig genetics and genomics research. GigaScience 2020, 9, giaa051. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Liao, Y.; Smyth, G.K.; Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef] [Green Version]
- Smid, M.; van de Werken, H.J.G.; van Riet, J.; van Galen, A.; de Weerd, V.; van der Vlugt-Daane, M.; Bril, S.I.; Lalmahomed, Z.S.; Kloosterman, W.P.; Wilting, S.M.; et al. Gene length corrected trimmed mean of M-values (GeTMM) processing of RNA-seq data performs similarly in intersample analyses while improving intrasample comparisons. BMC Bioinform. 2018, 19, 236. [Google Scholar] [CrossRef] [Green Version]
- Menzel, P.; Ng, K.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef] [Green Version]
- Lazov, C.; Belsham, G.; Bøtner, A.; Rasmussen, T. Full-Genome Sequences of Alphacoronaviruses and Astroviruses from Myotis and Pipistrelle Bats in Denmark. Viruses 2021, 13, 1073. [Google Scholar] [CrossRef]
- Correia, S.; Ventura, S.; Parkhouse, R.M. Identification and utility of innate immune system evasion mechanisms of ASFV. Virus Res. 2013, 173, 87–100. [Google Scholar] [CrossRef]
- Alejo, A.; Matamoros, T.; Guerra, M.; Andrés, G. A Proteomic Atlas of the African Swine Fever Virus Particle. J. Virol. 2018, 92, e01293-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Feature | Score | Description |
|---|---|---|
| Alertness and recumbency | 0 | Alert |
| 1 | Depressed/lethargic | |
| 2 | Only gets up when touched | |
| 4 | Gets up slowly when touched | |
| 6 | Remains recumbent when touched | |
| Body condition | 0 | Normal, full stomach |
| 1 | Empty stomach, sunken flanks | |
| 2 | Empty stomach, sunken flanks, loss of muscle mass | |
| 3 | Emaciated | |
| Skin | 0 | Normal |
| 1 | Minimal area of the skin with observed bleeding (<10% of the body) | |
| 2 | Moderate area of the skin with observed bleeding (10–25% of the body) | |
| 3 | Generalized skin bleeding (>25% of the body) | |
| Joints | 0 | No joint swelling |
| 1 | Swelling | |
| 4 | Severe swelling and lameness | |
| Respiration | 0 | Normal |
| 1 | Mildly labored | |
| 2 | Labored +/− cough | |
| 3 | Severely labored | |
| Eyes | 0 | Normal |
| 1 | Small amount of exudate | |
| 2 | Moderate amount of exudate | |
| Gastrointestinal and urinary tracts | 0 | No diarrhea |
| 1 | Mild diarrhea for less than 24 h | |
| 3 | Diarrhea for more than 24 h or vomiting | |
| 4 | Bloody diarrhea or blood in urine | |
| Neurology | 0 | No symptoms |
| 3 | Hesitant, unsteady walk, crossing-over of legs is corrected slowly | |
| 4 | Pronounced ataxia | |
| 6 | Paralysis or convulsions |
| Pig Number (Sampling Day) | Number of Input Reads | Number of Reads Uniquely Mapped on Pig Genome | Uniquely Mapped Reads (%) on Pig Genome | Number of Reads Mapped to ASFV Genome | Proportion of ASFV Reads/Total Reads (%) |
|---|---|---|---|---|---|
| Pig 9 (0 dpi) | 31979021 | 28863826 | 90.26 | 0 | 0 |
| Pig 9 (3 dpi) | 31807126 | 29207238 | 91.83 | 0 | 0 |
| Pig 9 (6 dpi) | 40093403 | 35472115 | 88.47 | 871681 | 2.17 |
| Pig 10 (0 dpi) | 32617753 | 29668789 | 90.96 | 0 | 0 |
| Pig 10 (3 dpi) | 46764030 | 43095231 | 92.15 | 638 | 0.0014 |
| Pig 10 (6 dpi) | 27095889 | 24288166 | 89.64 | 915507 | 3.38 |
| Pig 11 (0 dpi) | 45953582 | 39771420 | 86.55 | 0 | 0 |
| Pig 11 (3 dpi) | 46561959 | 41696207 | 89.55 | 616 | 0.0013 |
| Pig 11 (6 dpi) | 45970436 | 40372979 | 87.82 | 1939287 | 4.21 |
| Pig 12 (0 dpi) | 46326924 | 37609952 | 81.18 | 0 | 0 |
| Pig 12 (3 dpi) | 41406744 | 36549318 | 88.27 | 851 | 0.0021 |
| Pig 12 (6 dpi) | 45915279 | 41203225 | 89.74 | 1945879 | 4.23 |
| Gene or Feature Name | CDS Start (nt) | CDS End (nt) | Pig9- 6 dpi | Pig10- 6 dpi | Pig11- 6 dpi | Pig12- 6 dpi | 1 Mean Pigs 11–12 |
|---|---|---|---|---|---|---|---|
| CP312R | 128277 | 129200 | 66398 | 78462 | 138910 | 144010 | 141460 |
| CP204L | 125783 | 126367 | 59137 | 50204 | 128247 | 133890 | 131069 |
| MGF 100-1L | 180479 | 180904 | 50566 | 30183 | 108077 | 98680 | 103379 |
| A151R | 49652 | 50107 | 45198 | 45603 | 96565 | 89945 | 93255 |
| K205R | 64174 | 64791 | 29906 | 34834 | 64872 | 65805 | 65339 |
| I73R | 173088 | 173306 | 23887 | 24501 | 59167 | 54809 | 56988 |
| MGF 360-15R | 50346 | 51215 | 24275 | 23647 | 47350 | 49844 | 48597 |
| MGF 110-5L-6L | 9490 | 10107 | 18740 | 20840 | 42820 | 46048 | 44434 |
| A240L | 48633 | 49343 | 18353 | 15318 | 41882 | 45882 | 43882 |
| MGF 110-7L | 10314 | 10727 | 20780 | 20790 | 43090 | 42632 | 42861 |
| E165R | 167468 | 167965 | 17095 | 20759 | 40914 | 41374 | 41144 |
| MGF 100-3L | 181269 | 181577 | 15716 | 18157 | 36029 | 42209 | 39119 |
| F334L | 56956 | 57960 | 17153 | 17920 | 38103 | 39044 | 38574 |
| MGF 110-3L | 8239 | 8613 | 16201 | 22633 | 35842 | 34045 | 34944 |
| 285L | 11042 | 11326 | 17175 | 14716 | 34762 | 32868 | 33815 |
| I215L | 174794 | 175432 | 16341 | 14199 | 32361 | 32736 | 32549 |
| MGF 505-3R | 35760 | 36602 | 13836 | 11627 | 25567 | 30317 | 27942 |
| DP96R | 185339 | 185629 | 12107 | 10550 | 27516 | 28141 | 27829 |
| ASFV G ACD 00600 | 48000 | 48152 | 9902 | 7709 | 22675 | 22563 | 22619 |
| K196R | 65113 | 65703 | 9812 | 14044 | 22163 | 22183 | 22173 |
| Gene or Feature Name | CDS Start (nt) | CDS End (nt) | Pig9- 6 dpi | Pig10- 6 dpi | Pig11- 6 dpi | Pig12- 6 dpi | 1 Mean Pigs 11–12 | Gene Product Properties (If Known) |
|---|---|---|---|---|---|---|---|---|
| ASFV G ACD 01960 | 187401 | 187532 | 141 | 114 | 292 | 254 | 273 | |
| ASFV G ACD 00190 | 12456 | 12581 | 81 | 122 | 265 | 276 | 271 | |
| E423R | 163803 | 165074 | 93 | 126 | 240 | 285 | 263 | |
| H171R | 153250 | 153765 | 100 | 117 | 264 | 245 | 255 | |
| ASFV G ACD 00090 | 7647 | 7760 | 107 | 150 | 229 | 236 | 233 | |
| L11L | 183821 | 184102 | 75 | 79 | 233 | 204 | 219 | |
| B407L | 107261 | 108499 | 52 | 119 | 189 | 185 | 187 | |
| O61R | 129795 | 129980 | 60 | 126 | 197 | 177 | 187 | P12 attachment protein |
| B117L | 106907 | 107254 | 66 | 81 | 196 | 165 | 181 | TR containing protein |
| MGF 505-2R | 34093 | 35673 | 66 | 100 | 197 | 155 | 176 | |
| B318L | 96276 | 97232 | 51 | 69 | 152 | 182 | 167 | Prenyltransferase |
| E301R | 165225 | 166130 | 34 | 74 | 146 | 159 | 153 | Proliferating cell nuclear antigen-like protein |
| EP153R | 73808 | 74284 | 77 | 38 | 139 | 129 | 134 | C-type lectin-like |
| ASFV G ACD 01020 | 92901 | 93059 | 47 | 37 | 140 | 88 | 114 | |
| B119L | 95936 | 96295 | 38 | 39 | 75 | 109 | 92 | FAD-dependent thiol oxidase |
| B175L | 108527 | 109054 | 28 | 47 | 82 | 100 | 91 | Late TF VLTF-2 |
| S273R | 147670 | 148491 | 20 | 37 | 70 | 89 | 80 | SUMO-1-like protease |
| E183L | 163218 | 163772 | 28 | 46 | 84 | 69 | 77 | P54, Virus entry |
| ASFV G ACD 01870 | 182604 | 182741 | 20 | 12 | 85 | 66 | 76 | |
| S183L | 147058 | 147609 | 18 | 44 | 55 | 76 | 66 | |
| E146L | 166164 | 166604 | 25 | 45 | 69 | 56 | 63 | PSP |
| ASFV G ACD 00360 | 20169 | 20285 | 14 | 14 | 56 | 48 | 52 | |
| ASFV G ACD 00210 | 13461 | 13652 | 18 | 17 | 29 | 51 | 40 | |
| ASFV G ACD 00240 | 14570 | 14680 | 13 | 11 | 26 | 40 | 33 | |
| EP84R | 71306 | 71560 | 7 | 13 | 13 | 26 | 20 | PSP |
| 1. Gene Name | Gene Product Properties (If Known) | Start | End | Pig11- 3 dpi | Pig12- 3 dpi | Pig9- 6 dpi | Pig10- 6 dpi | Pig11- 6 dpi | Pig12- 6 dpi | Mean Pigs 11 and 12 |
|---|---|---|---|---|---|---|---|---|---|---|
| I73R | Tandem repeat sequence | 173088 | 173306 | 9 | 13 | 15236 | 22260 | 37512 | 29371 | 33441 |
| MGF 100-1L | 180479 | 180904 | 9 | 16 | 16544 | 14066 | 35147 | 27124 | 31136 | |
| CP204L | P32 (P30) phosphoprotein | 125783 | 126367 | 9 | 16 | 14080 | 17027 | 30351 | 26783 | 28567 |
| A151R | Redox pathway | 49652 | 50107 | 8 | 13 | 13812 | 19851 | 29333 | 23093 | 26213 |
| CP312R | Immunodominant protein | 128277 | 129200 | 6 | 8 | 10003 | 16837 | 20801 | 18227 | 19514 |
| MGF 100-3L | 181269 | 181577 | 4 | 7 | 7095 | 11676 | 16168 | 16009 | 16088 | |
| 285L | 11042 | 11326 | 3 | 8 | 8409 | 10263 | 16917 | 13520 | 15219 | |
| K205R | In virus factories | 64174 | 64791 | 3 | 8 | 6740 | 11182 | 14532 | 12459 | 13495 |
| MGF 110-7L | 10314 | 10727 | 7 | 3 | 6996 | 9970 | 14420 | 12059 | 13239 | |
| DP96R | 185339 | 185629 | 6 | 4 | 5805 | 7205 | 13114 | 11336 | 12225 | |
| MGF 110-3L | 8239 | 8613 | 4 | 4 | 6023 | 11986 | 13245 | 10634 | 11940 | |
| E165R | dUTPase | 167468 | 167965 | 3 | 5 | 4783 | 8273 | 11378 | 9725 | 10551 |
| MGF 110-5L-6L | 9490 | 10107 | 3 | 4 | 4223 | 6690 | 9592 | 8719 | 9155 | |
| A240L | Thymidylate kinase | 48633 | 49343 | 1 | 3 | 3594 | 4273 | 8153 | 7549 | 7851 |
| MGF 360-15R | 50346 | 51215 | 2 | 3 | 3884 | 5390 | 7531 | 6701 | 7116 | |
| I215L | Ubiquitin conjugating enzyme | 174794 | 175432 | 2 | 3 | 3561 | 4408 | 7010 | 5994 | 6502 |
| L83L | 4878 | 5123 | 3 | 5 | 2775 | 2009 | 6343 | 5185 | 5764 | |
| MGF 110-4L | Has KDEL-like domain | 8927 | 9301 | 2 | 3 | 2689 | 4421 | 6542 | 4684 | 5613 |
| A104R | Histone-like | 48322 | 48636 | 2 | 4 | 2532 | 3481 | 6119 | 4943 | 5531 |
| F334L | Ribonucleotide reductase subunit | 56956 | 57960 | 2 | 2 | 2376 | 3535 | 5245 | 4543 | 4894 |
| K196R | Thymidine kinase | 65113 | 65703 | 1 | 2 | 2312 | 4715 | 5192 | 4392 | 4792 |
| MGF 110-2L | 7828 | 8142 | 2 | 1 | 2486 | 3884 | 4694 | 4298 | 4496 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olesen, A.S.; Kodama, M.; Lohse, L.; Accensi, F.; Rasmussen, T.B.; Lazov, C.M.; Limborg, M.T.; Gilbert, M.T.P.; Bøtner, A.; Belsham, G.J. Identification of African Swine Fever Virus Transcription within Peripheral Blood Mononuclear Cells of Acutely Infected Pigs. Viruses 2021, 13, 2333. https://doi.org/10.3390/v13112333
Olesen AS, Kodama M, Lohse L, Accensi F, Rasmussen TB, Lazov CM, Limborg MT, Gilbert MTP, Bøtner A, Belsham GJ. Identification of African Swine Fever Virus Transcription within Peripheral Blood Mononuclear Cells of Acutely Infected Pigs. Viruses. 2021; 13(11):2333. https://doi.org/10.3390/v13112333
Chicago/Turabian StyleOlesen, Ann Sofie, Miyako Kodama, Louise Lohse, Francesc Accensi, Thomas Bruun Rasmussen, Christina M. Lazov, Morten T. Limborg, M. Thomas P. Gilbert, Anette Bøtner, and Graham J. Belsham. 2021. "Identification of African Swine Fever Virus Transcription within Peripheral Blood Mononuclear Cells of Acutely Infected Pigs" Viruses 13, no. 11: 2333. https://doi.org/10.3390/v13112333
APA StyleOlesen, A. S., Kodama, M., Lohse, L., Accensi, F., Rasmussen, T. B., Lazov, C. M., Limborg, M. T., Gilbert, M. T. P., Bøtner, A., & Belsham, G. J. (2021). Identification of African Swine Fever Virus Transcription within Peripheral Blood Mononuclear Cells of Acutely Infected Pigs. Viruses, 13(11), 2333. https://doi.org/10.3390/v13112333