
Characterization of Ictalurid herpesvirus 1 Glycoprotein ORF59 and Its Potential Role on Virus Entry into the Host Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell and Virus
2.2. Antibodies Preparation
2.3. Plasmid Construction
2.4. Design and Synthesis of Short Hairpin RNAs (shRNAs)
2.5. Plasmid Transfection
2.6. CCV DNA Synthesis Inhibition
2.7. Fluorescence Observation
2.8. Quantitative Real-Time Reverse Transcription PCR
2.9. Cell Plasma Membrane Protein Isolation
2.10. Virus Titration
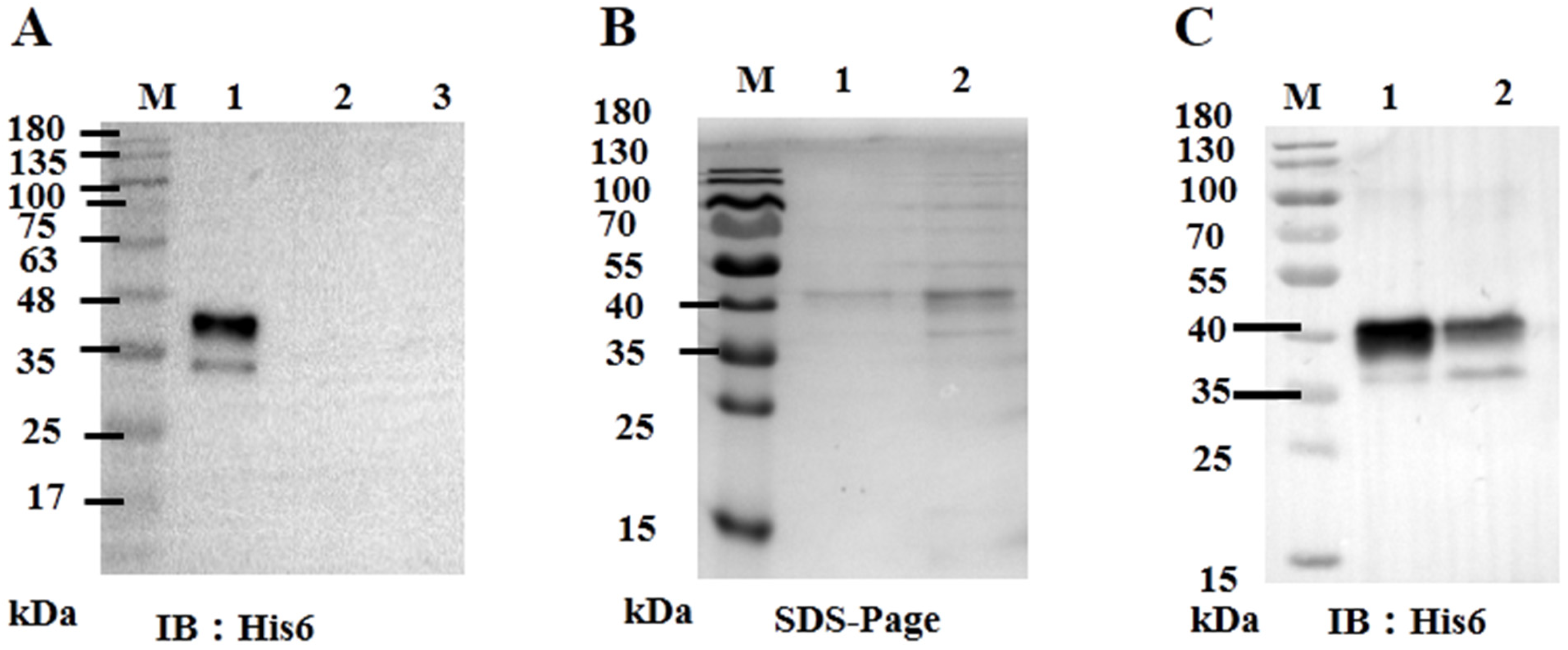
2.11. Baculovirus Expression of ORF59 Protein
2.12. Western Blotting Assay
2.13. Protein Blocking Assay
2.14. Statistical Analysis
3. Results
3.1. CCV ORF59 Is a Late Expression Gene during CCV Infection
3.2. CCV ORF59 Is a Membrane Associated Protein
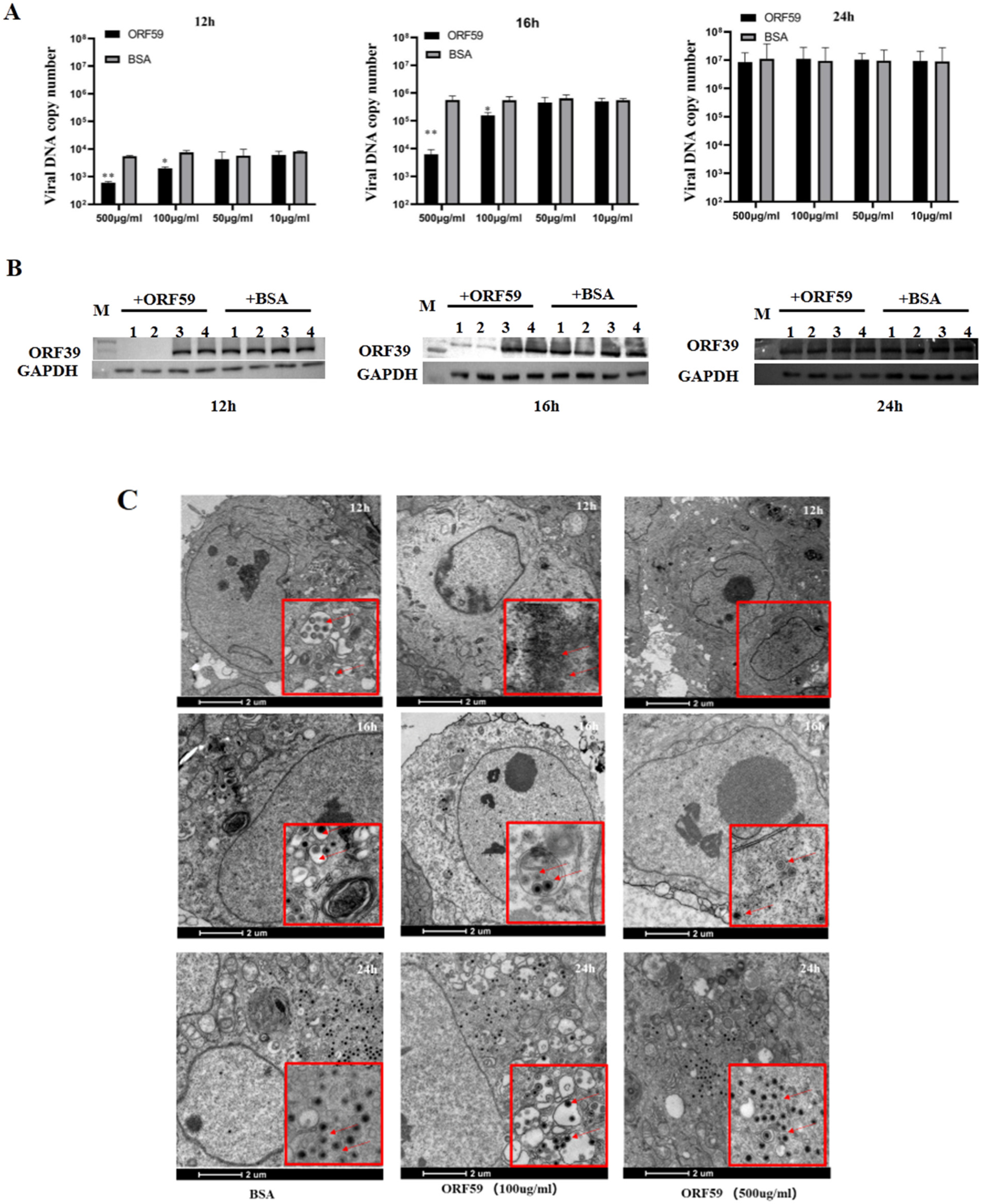
3.3. Recombinant ORF59 Protein Blocks CCV Infection by Abrogation of Cellular Binding
3.4. ORF59 Knockdown Reduces Production of Infectious Virus Particles in CCO Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hao, K.; Yuan, S.; Yu, F.; Chen, X.H.; Bian, W.J.; Feng, Y.H.; Zhao, Z. Acyclovir inhibits channel catfish virus replication and protects channel catfish ovary cells from apoptosis. Virus Res. 2021, 292, 198249. [Google Scholar] [CrossRef]
- Chousterman, S.; Lacasa, M.; Sheldrick, P.J. Physical Map of the Channel Catfish Virus Genome: Location of Sites for Restriction Endonucleases EcoRI, HindIII, HpaI, and XbaI. J. Virol. 1979, 31, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Kucuktas, H.; Brady, Y.; Tüzün, S. Cloning and expression of a putative glycoprotein gene of channel catfish virus using baculovirus expression system. Dis. Aquat. Org. 1998, 34, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Farber, F.E. Channel catfish virus: Physicochemical properties of the viral genome and identification of viral polypeptides. Virology 1980, 103, 267–278. [Google Scholar] [CrossRef]
- Davison, A.J.; Davison, M.D. Identification of Structural Proteins of Channel Catfish Virus by Mass Spectrometry. Virology 1995, 206, 1035–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuang, F.; Luo, Y.; Xiong, X.-P.; Weng, S.; Li, Y.; He, J.; Dong, C. Virions proteins of an RSIV-type megalocytivirus from spotted knifejaw Oplegnathus punctatus (SKIV-ZJ07). Virology 2013, 437, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, C.; Weng, S.; Li, W.; Li, X.; Yi, Y.; Liang, Q.; He, J. Characterization of a new cell line from caudal fin of koi, Cyprinus carpio koi, and first isolation of cyprinid herpesvirus 3 in China. Virus Res. 2011, 161, 140–149. [Google Scholar] [CrossRef]
- Banfield, B.W.; Leduc, Y.; Esford, L.; Schubert, K.; Tufaro, F. Sequential isolation of proteoglycan synthesis mutants by using herpes simplex virus as a selective agent: Evidence for a proteoglycan-independent virus entry pathway. J. Virol. 1995, 69, 3290–3298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spear, P.G. A First Step toward Understanding Membrane Fusion Induced by Herpes Simplex Virus. Mol. Cell 2001, 8, 2–4. [Google Scholar] [CrossRef]
- Banerjee, N.; Mukhopadhyay, S. Viral glycoproteins: Biological role and application in diagnosis. Virusdisease 2016, 27, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Reuter, N.; Kropff, B.; Schneiderbanger, J.K.; Alt, M.; Krawczyk, A.; Sinzger, C.; Winkler, T.H.; Britt, W.J.; Mach, M.; Thomas, M. Cell Fusion Induced by a Fusion-Active Form of Human Cytomegalovirus Glycoprotein B (gB) Is Inhibited by Antibodies Directed at Antigenic Domain 5 in the Ectodomain of gB. J. Virol. 2020, 94, e01276-20. [Google Scholar] [CrossRef] [PubMed]
- Vanderheijden, N.; Hanson, L.A.; Thiryc, E.; Martial, J.A. Channel Catfish Virus Gene 50 Encodes a Secreted, Mucin-like Glycoprotein. Virology 1999, 257, 220–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, L.; Subramaniam, K.; Waltzek, T.B.; Bowser, P.R.; Hanson, L. Cytochrome oxidase gene sequencing reveals channel catfish ovary cell line is contaminated with brown bullhead cells. J. Fish Dis. 2021, 44, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Li, S.; Chen, H.; Hao, K.; Meng, L.; Yang, J.; Zhao, Z. Multiple AT-rich sequences function as a cis -element in the ORF3 promoter in channel catfish virus ( Ictalurid herpesvirus 1). J. Fish Dis. 2021, 44, 1609–1617. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Yu, F.; Wang, L.; Li, W.; Lu, L. Identification of a novel membrane-associated protein from the S7 segment of grass carp reovirus. J. Gen. Virol. 2019, 100, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Caplen, N.J.; Zheng, Z.; Falgout, B.; Morgan, R.A. Inhibition of Viral Gene Expression and Replication in Mosquito Cells by dsRNA-Triggered RNA Interference. Mol. Ther. 2002, 6, 243–251. [Google Scholar] [CrossRef]
- Li, B.; Fan, Y.-D.; Li, Y.-Q.; Xu, J.; Zhou, Y.; Zeng, L.-B. Highly efficient inhibition on replication of grass carp reovirus mediated by chemically synthesized small interfering RNAs. Bing Du Xue Bao 2009, 25, 388–394. [Google Scholar] [CrossRef]
- Kumar, P.; Wu, H.; McBride, J.L.; Jung, K.-E.; Kim, M.H.; Davidson, B.; Lee, S.K.; Shankar, P.; Manjunath, N. Transvascular delivery of small interfering RNA to the central nervous system. Nat. Cell Biol. 2007, 448, 39–43. [Google Scholar] [CrossRef]
- Ma, Z.; Jacobs, S.R.; West, J.A.; Stopford, C.; Zhang, Z.; Davis, Z.; Barber, G.N.; Glaunsinger, B.A.; Dittmer, D.P.; Damania, B. Modulation of the cGAS-STING DNA sensing pathway by gammaherpesviruses. Proc. Natl. Acad. Sci. USA 2015, 112, E4306–E4315. [Google Scholar] [CrossRef] [Green Version]
- Stingley, R.L.; Gray, W.L. Transcriptional regulation of the channel catfish virus genome direct repeat region. J. Gen. Virol. 2000, 81, 2005–2010. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.P.; Melancon, J.M.; Baines, J.D.; Kousoulas, K.G. The Herpes Simplex Virus Type 1 UL20 Protein Modulates Membrane Fusion Events during Cytoplasmic Virion Morphogenesis and Virus-Induced Cell Fusion. J. Virol. 2004, 78, 5347–5357. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.C.; Chouljenko, V.N.; Chouljenko, D.V.; Boudreaux, M.J.; Kousoulas, K.G. The Herpes Simplex Virus Type 1 Glycoprotein D (gD) Cytoplasmic Terminus and Full-Length gE Are Not Essential and Do Not Function in a Redundant Manner for Cytoplasmic Virion Envelopment and Egress. J. Virol. 2009, 83, 6115–6124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Shen, B.; Wang, M.; Cheng, A.; Yang, Q.; Wu, Y.; Huang, J.; Tian, B.; Jia, R.; Liu, M.; et al. The intracellular domain of duck plague virus glycoprotein E affects UL11 protein incorporation into viral particles. Vet. Microbiol. 2021, 257, 109078. [Google Scholar] [CrossRef]
- Wu, K.; Mu, Y.; Hu, J.; Lu, L.; Zhang, X.; Yang, Y.; Li, Y.; Liu, F.; Song, D.; Zhu, Y.; et al. Simultaneously inhibition of HIV and HBV replication through a dual small interfering RNA expression system. Antivir. Res. 2007, 74, 142–149. [Google Scholar] [CrossRef]
- Silverstein, P.S.; Bird, R.C.; van Santen, V.L.; Nusbaum, K.E. Immediate-early transcription from the channel catfish virus genome: Characterization of two immediate-early transcripts. J. Virol. 1995, 69, 3161–3166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nusbaum, K.E.; Smith, B.F.; DeInnocentes, P.; Bird, R. Protective immunity induced by DNA vaccination of channel catfish with early and late transcripts of the channel catfish herpesvirus (IHV-1). Vet. Immunol. Immunopathol. 2002, 84, 151–168. [Google Scholar] [CrossRef]
- Mody, P.H.; Pathak, S.; Hanson, L.K.; Spencer, J.V. Herpes Simplex Virus: A Versatile Tool for Insights Into Evolution, Gene Delivery, and Tumor Immunotherapy. Virol. Res. Treat. 2020, 11, 1178122X20913274. [Google Scholar] [CrossRef] [PubMed]
- Campadelli-Fiume, G.; Amasio, M.; Avitabile, E.; Cerretani, A.; Forghieri, C.; Gianni, T.; Menotti, L. The multipartite system that mediates entry of herpes simplex virus into the cell. Rev. Med. Virol. 2007, 17, 313–326. [Google Scholar] [CrossRef]
- Narayanan, K.K.; Procko, E. Deep Mutational Scanning of Viral Glycoproteins and Their Host Receptors. Front. Mol. Biosci. 2021, 8, 636660. [Google Scholar] [CrossRef]
- Antoine, T.E.; Park, P.J.; Shukla, D. Glycoprotein targeted therapeutics: A new era of anti-herpes simplex virus-1 therapeutics. Rev. Med. Virol. 2013, 23, 194–208. [Google Scholar] [CrossRef] [Green Version]
- Madavaraju, K.; Koganti, R.; Volety, I.; Yadavalli, T.; Shukla, D. Herpes Simplex Virus Cell Entry Mechanisms: An Update. Front. Cell. Infect. Microbiol. 2021, 10, 852. [Google Scholar] [CrossRef]
- Chouljenko, D.V.; Kim, I.-J.; Chouljenko, V.N.; Subramanian, R.; Walker, J.D.; Kousoulas, K.G. Functional Hierarchy of Herpes Simplex Virus 1 Viral Glycoproteins in Cytoplasmic Virion Envelopment and Egress. J. Virol. 2012, 86, 4262–4270. [Google Scholar] [CrossRef] [Green Version]
- Lye, M.F.; Wilkie, A.R.; Filman, D.J.; Hogle, J.M.; Coen, D.M. Getting to and through the inner nuclear membrane during herpesvirus nuclear egress. Curr. Opin. Cell Biol. 2017, 46, 9–16. [Google Scholar] [CrossRef]
- Dawar, F.U.; Hu, X.Q.; Zhao, L.J.; Dong, X.X.; Xiong, Y.; Zhou, M.R.; Liang, S.; Badu, S.; Li, J.; Mei, J.; et al. Transcriptomic analysis reveals differentially expressed genes and a unique apoptosis pathway in channel catfish ovary cells after infection with the channel catfish virus. J. Fish Dis. 2017, 71, 58–68. [Google Scholar] [CrossRef]
- Melancon, J.M.; Foster, T.P.; Kousoulas, K.G. Genetic Analysis of the Herpes Simplex Virus Type 1 UL20 Protein Domains Involved in Cytoplasmic Virion Envelopment and Virus-Induced Cell Fusion. J. Virol. 2004, 78, 7329–7343. [Google Scholar] [CrossRef] [Green Version]
- Dollery, S.J. Towards Understanding KSHV Fusion and Entry. Viruses 2019, 11, 1073. [Google Scholar] [CrossRef] [Green Version]
- Stegmann, C.; Rothemund, F.; Sampaio, K.L.; Adler, B.; Sinzger, C. The N Terminus of Human Cytomegalovirus Glycoprotein O Is Important for Binding to the Cellular Receptor PDGFRα. J. Virol. 2019, 93, e00138-19. [Google Scholar] [CrossRef] [Green Version]
- Atanasiu, D.; Saw, W.T.; Lazear, E.; Whitbeck, J.C.; Cairns, T.M.; Lou, H.; Eisenberg, R.J.; Cohen, G.H.J. Using antibod-ies and mutants to localize the presumptive gH/gL binding site on herpes simplex virus gD. J. Virol. 2018, 92, e01694-18. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) | Purpose |
|---|---|---|
| ORF59-F | AGGCGTATCACCAACTCACC | RT-PCR and PCR |
| ORF59-R | ACCGAACTGGTGAGGATCAG | RT-PCR and PCR |
| ORF3-F | GATGAGGGCGACGACACTAT | PCR |
| ORF-R | AGTCCCAGTCGGAAGTCTCA | PCR |
| ORF39-F | GAAGATAGCCCGTCTCACCG | PCR |
| ORF39-R | ATCTCGATCAGCATCTGGCG | PCR |
| 18sRNA-F | CGCCCCGCCCAACTCGCCTGAATA | RT-PCR |
| 18sRNA-R | CGAATGCCCCCGCCGTCCCTCTTA | RT-PCR |
| eGFP-ORF59F | CTCAAG CTTACCATG GTCGGCAAAGGTCTCC | ORF amplification |
| eGFP-ORF59R | CCGGGATCCCGCCCGGGCAGGTGTGTAGT | ORF amplification |
| shRNA59-158F | CACCAACTCACCAAGCTACAAG | RT-PCR |
| shRNA59-158R | CCTACCAGGTCTATCACCGAAC | RT-PCR |
| shRNA59-257F | CGGTGATAGACCTGGTAGGG | RT-PCR |
| shRNA59-257R | GCGACGAAAACGATCATCAG | RT-PCR |
| shRNA59-422F | ACCTCTTTCGGGTTCGATGT | RT-PCR |
| shRNA59-422R | TCAGGATACTGAACACCGTGA | RT-PCR |
| shRNA59-948F | GGTTGGGGACAATAATCGAA | RT-PCR |
| ahRNA59-948R | TTCTCATACCGGGAATGGTG | RT-PCR |
| cVRT-F | GAAGATAGCCCGTCTCACCG | qPCR |
| cVRT-R | ATCTCGATCAGCATCTGGCG | qPCR |
| shORF3-F | CTGGAATCCTCCTCCTCCTT | qRT-PCR |
| shORF3-R | GTCGGAGACGGGAGAGTACA | qRT-PCR |
| shORF5-F | CCGTCTTCGTGTACCTGGAG | qRT-PCR |
| shORF5-R | CCACGCCTCGTATCTTTCG | qRT-PCR |
| shORF39-F | GGGTCTCATCTTTGCCGATA | qRT-PCR |
| shORF39-R | AGTTTGAGCGAGAACCCGTA | qRT-PCR |
| pBHTA-59F | ttttcagggcgccatggatccGATGGTCGGCAAAGGTCTCCC | ORF amplification |
| pBHTA-59R | tcgacgtaggcctttgaattcTCACGCCCGGGCAGGTGT | ORF amplification |
| pUC/M13F | CCCAGTCACGACGTTGTAAAACG | PCR |
| pUC/M13R | AGCGGATAACAATTTCACACAGG | PCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.-X.; Yu, F.; Chen, H.-X.; Zhang, X.-D.; Meng, L.-H.; Hao, K.; Zhao, Z. Characterization of Ictalurid herpesvirus 1 Glycoprotein ORF59 and Its Potential Role on Virus Entry into the Host Cells. Viruses 2021, 13, 2393. https://doi.org/10.3390/v13122393
Li S-X, Yu F, Chen H-X, Zhang X-D, Meng L-H, Hao K, Zhao Z. Characterization of Ictalurid herpesvirus 1 Glycoprotein ORF59 and Its Potential Role on Virus Entry into the Host Cells. Viruses. 2021; 13(12):2393. https://doi.org/10.3390/v13122393
Chicago/Turabian StyleLi, Shu-Xin, Fei Yu, Hong-Xun Chen, Xiao-Dong Zhang, Li-Hui Meng, Kai Hao, and Zhe Zhao. 2021. "Characterization of Ictalurid herpesvirus 1 Glycoprotein ORF59 and Its Potential Role on Virus Entry into the Host Cells" Viruses 13, no. 12: 2393. https://doi.org/10.3390/v13122393
APA StyleLi, S. -X., Yu, F., Chen, H. -X., Zhang, X. -D., Meng, L. -H., Hao, K., & Zhao, Z. (2021). Characterization of Ictalurid herpesvirus 1 Glycoprotein ORF59 and Its Potential Role on Virus Entry into the Host Cells. Viruses, 13(12), 2393. https://doi.org/10.3390/v13122393