
First Description of a Temperate Bacteriophage (vB_FhiM_KIRK) of Francisella hispaniensis Strain 3523

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media and Growth Conditions
2.2. DNA Techniques and PCR Analysis
2.3. Transformation of Bacteria
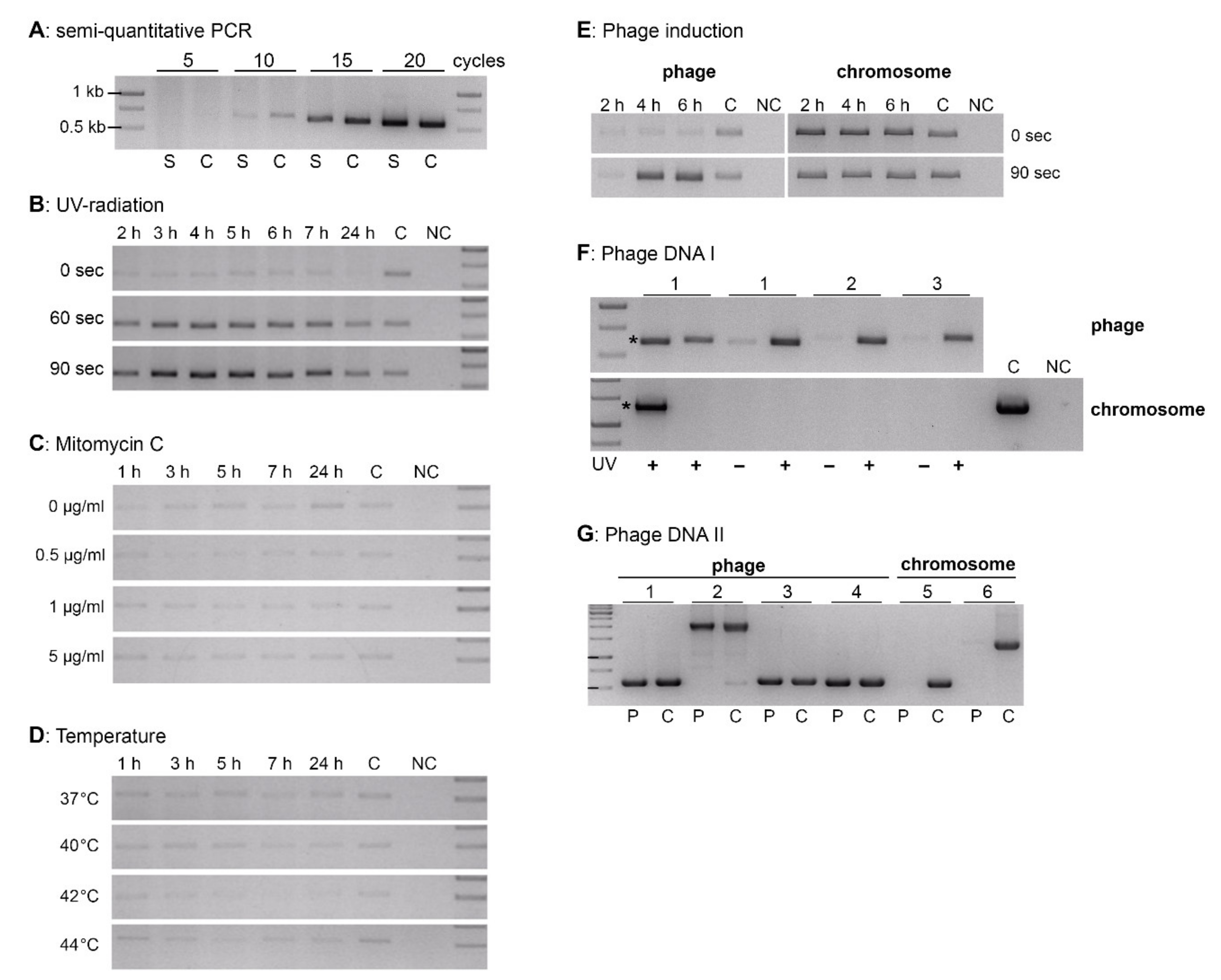
2.4. Phage Induction Experiments
2.5. Cell Disruption and Phage Purification
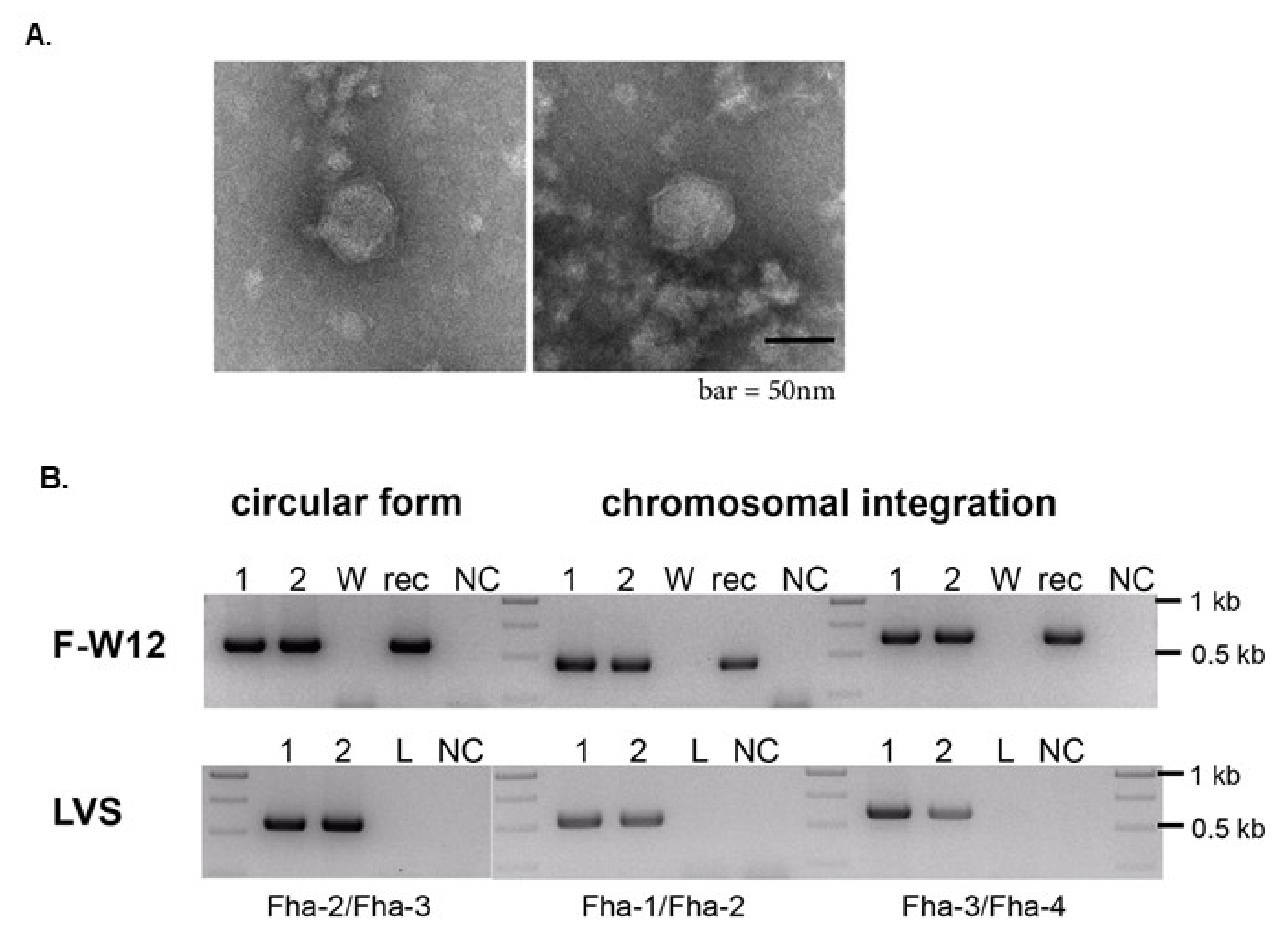
2.6. Electron Microscopy (EM)
2.7. Phage Plate (Spot) Test and Phage Infection Assays
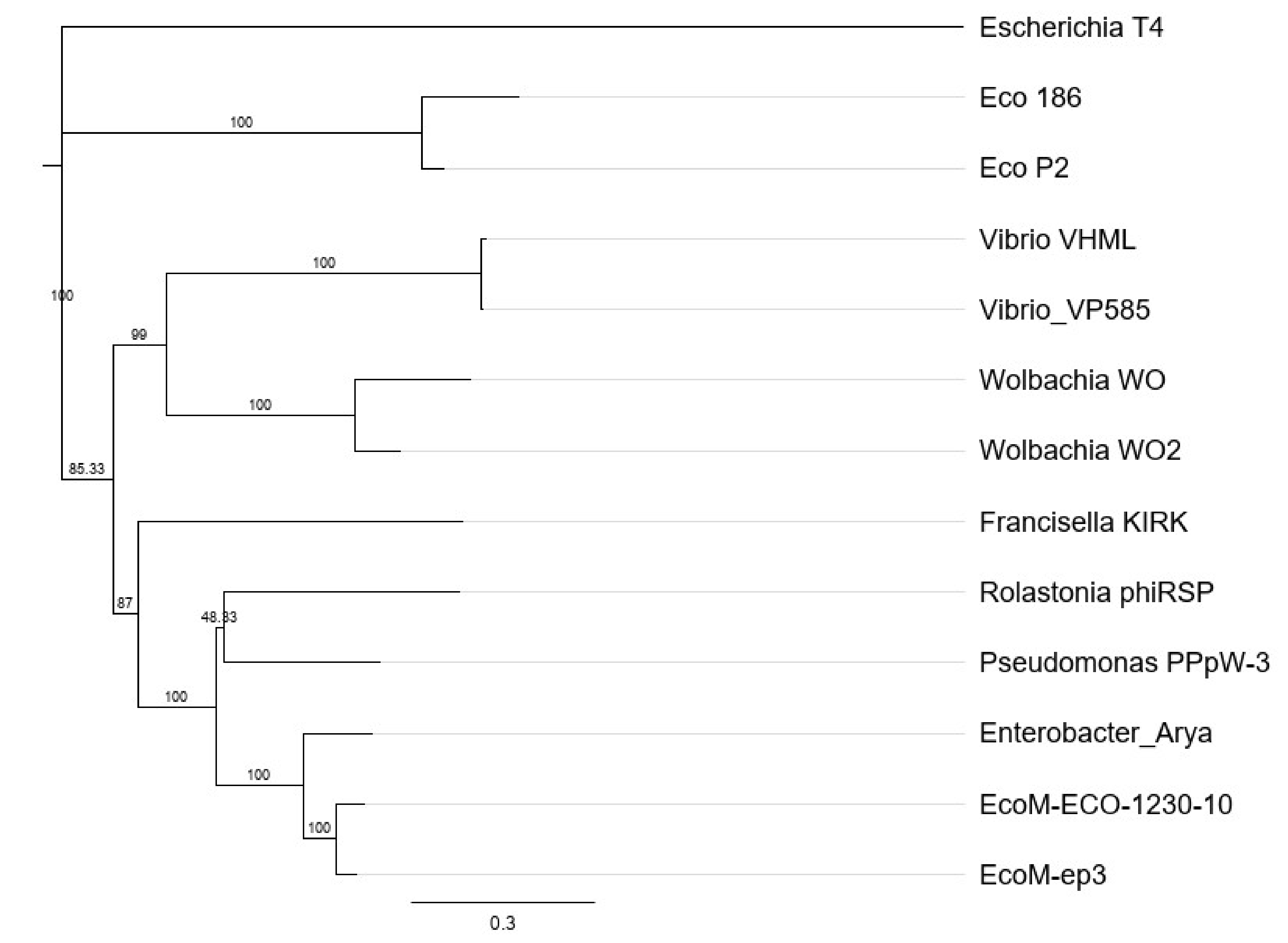
2.8. Phylogenetic Analysis
3. Results
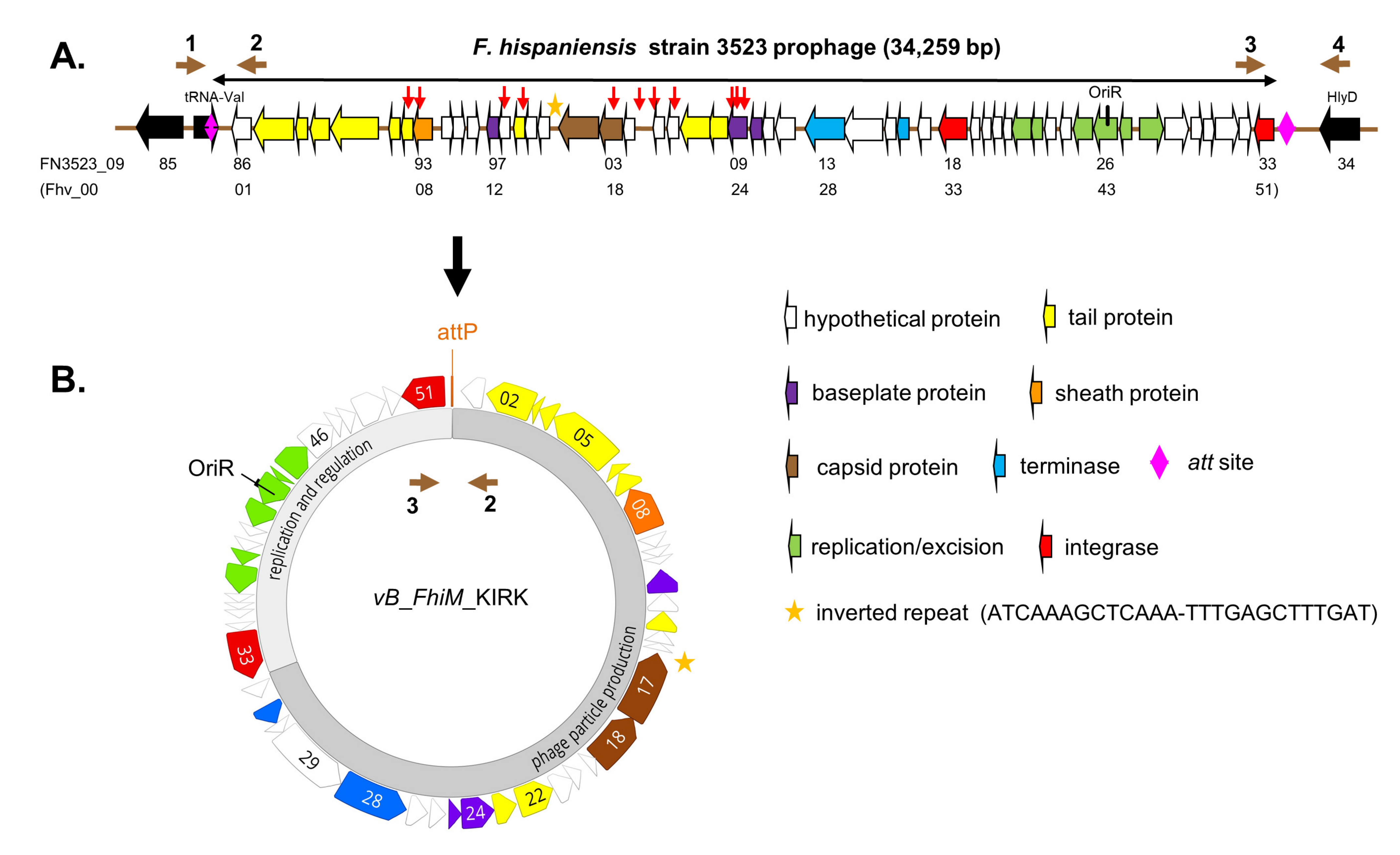
3.1. Genetic Organization and Open Reading Frames (ORFs)
3.2. Prophage Induction and Phage Characterization
3.3. The Recombinant KIRK (KIRKrec)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ellis, J.; Oyston, P.C.; Green, M.; Titball, R.W. Tularemia. Clin. Microbiol. Rev. 2002, 15, 631–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjostedt, A. Special Topic on Francisella tularensis and Tularemia. Front. Microbiol. 2011, 2, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keim, P.; Johansson, A.; Wagner, D.M. Molecular epidemiology, evolution, and ecology of Francisella. Ann. N. Y. Acad. Sci. 2007, 1105, 30–66. [Google Scholar] [CrossRef] [PubMed]
- Clarridge, J.E., 3rd; Raich, T.J.; Sjosted, A.; Sandstrom, G.; Darouiche, R.O.; Shawar, R.M.; Georghiou, P.R.; Osting, C.; Vo, L. Characterization of two unusual clinically significant Francisella strains. J. Clin. Microbiol. 1996, 34, 1995–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollis, D.G.; Weaver, R.E.; Steigerwalt, A.G.; Wenger, J.D.; Moss, C.W.; Brenner, D.J. Francisella philomiragia comb. nov. (formerly Yersinia philomiragia) and Francisella tularensis biogroup novicida (formerly Francisella novicida) associated with human disease. J. Clin. Microbiol. 1989, 27, 1601–1608. [Google Scholar] [CrossRef] [Green Version]
- Whipp, M.J.; Davis, J.M.; Lum, G.; de Boer, J.; Zhou, Y.; Bearden, S.W.; Petersen, J.M.; Chu, M.C.; Hogg, G. Characterization of a novicida-like subspecies of Francisella tularensis isolated in Australia. J. Med. Microbiol. 2003, 52, 839–842. [Google Scholar] [CrossRef]
- Sjodin, A.; Svensson, K.; Ohrman, C.; Ahlinder, J.; Lindgren, P.; Duodo, S.; Hnath, J.; Burans, J.P.; Johansson, A.; Colquhoun, D.J.; et al. Genome characterisation of the genus Francisella reveals insight into similar evolutionary paths in pathogens of mammals and fish. BMC Genom. 2012, 13, 268. [Google Scholar] [CrossRef] [Green Version]
- Escudero, R.; Elia, M.; Saez-Nieto, J.A.; Menendez, V.; Toledo, A.; Royo, G.; Rodriguez-Vargas, M.; Whipp, M.J.; Gil, H.; Jado, I.; et al. A possible novel Francisella genomic species isolated from blood and urine of a patient with severe illness. Clin. Microbiol. Infect. 2010, 16, 1026–1030. [Google Scholar] [CrossRef] [Green Version]
- Huber, B.; Escudero, R.; Busse, H.J.; Seibold, E.; Scholz, H.C.; Anda, P.; Kampfer, P.; Splettstoesser, W.D. Description of Francisella hispaniensis sp. nov., isolated from human blood, reclassification of Francisella novicida (Larson et al. 1955) Olsufiev et al. 1959 as Francisella tularensis subsp. novicida comb. nov. and emended description of the genus Francisella. Int. J. Syst. Evol. Microbiol. 2010, 60, 1887–1896. [Google Scholar] [CrossRef]
- Siddaramappa, S.; Challacombe, J.F.; Petersen, J.M.; Pillai, S.; Hogg, G.; Kuske, C.R. Common ancestry and novel genetic traits of Francisella novicida-like isolates from North America and Australia as revealed by comparative genomic analyses. Appl. Environ. Microbiol. 2011, 77, 5110–5122. [Google Scholar] [CrossRef] [Green Version]
- Schunder, E.; Rydzewski, K.; Grunow, R.; Heuner, K. First indication for a functional CRISPR/Cas system in Francisella tularensis. Int. J. Med. Microbiol. 2013, 303, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Deveau, H.; Garneau, J.E.; Moineau, S. CRISPR/Cas system and its role in phage-bacteria interactions. Annu. Rev. Microbiol. 2010, 64, 475–493. [Google Scholar] [CrossRef] [PubMed]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the immune system of bacteria and archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunin, V.; Sorek, R.; Hugenholtz, P. Evolutionary conservation of sequence and secondary structures in CRISPR repeats. Genome Biol. 2007, 8, R61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, K.S.; Aravind, L.; Wolf, Y.I.; Koonin, E.V. Unification of Cas protein families and a simple scenario for the origin and evolution of CRISPR-Cas systems. Biol. Direct. 2011, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, K.S.; Haft, D.H.; Barrangou, R.; Brouns, S.J.; Charpentier, E.; Horvath, P.; Moineau, S.; Mojica, F.J.; Wolf, Y.I.; Yakunin, A.F.; et al. Evolution and classification of the CRISPR-Cas systems. Nat. Rev. Microbiol. 2011, 9, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Rydzewski, K.; Tlapak, H.; Niehaus, I.P.; Dabrowski, P.W.; Grunow, R.; Heuner, K. Identification and characterization of episomal forms of integrative genomic islands in the genus Francisella. Int. J. Med. Microbiol. 2015, 305, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Tlapak, H.; Koppen, K.; Rydzewski, K.; Grunow, R.; Heuner, K. Construction of a New Phage Integration Vector pFIV-Val for Use in Different Francisella Species. Front. Cell. Infect. Microbiol. 2018, 8, 75. [Google Scholar] [CrossRef]
- Casjens, S. Prophages and bacterial genomics: What have we learned so far? Mol. Microbiol. 2003, 49, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, M.; Kurokawa, K.; Hayashi, T. Diversification of Escherichia coli genomes: Are bacteriophages the major contributors? Trends Microbiol. 2001, 9, 481–485. [Google Scholar] [CrossRef]
- Banks, D.J.; Beres, S.B.; Musser, J.M. The fundamental contribution of phages to GAS evolution, genome diversification and strain emergence. Trends Microbiol. 2002, 10, 515–521. [Google Scholar] [CrossRef]
- Koliaditskaia, L.S.; Kuchina, K.V.; Shmurygina, A.A. [Tularemia bacteriophage; preliminary communication]. Zh. Mikrobiol. Epidemiol. Immunobiol. 1959, 30, 13–17. [Google Scholar] [PubMed]
- Lammertyn, E.; Vande Voorde, J.; Meyen, E.; Maes, L.; Mast, J.; Anne, J. Evidence for the presence of Legionella bacteriophages in environmental water samples. Microb. Ecol. 2008, 56, 191–197. [Google Scholar] [CrossRef]
- Grigor’ev, A.A.; Bondarev, V.P.; Borisevich, I.V.; Darmov, I.V.; Mironin, A.V.; Zolotarev, A.G.; Pogorel’skii, I.P.; Ianov, D.S. [Temperate Legionella bacteriophage: Discovery and characteristics]. Zh. Mikrobiol. Epidemiol. Immunobiol. 2008, 86–88. [Google Scholar]
- Alharby, A.M. Isolation and charakterization of a novel bacteriophage, ASC10, that lyses Francisella tularensis. Ph.D. Theses, Colorado State University, Fort Collins, CO, USA, 2014. [Google Scholar]
- Sampson, T.R.; Weiss, D.S. Cas9-dependent endogenous gene regulation is required for bacterial virulence. Biochem. Soc. Trans. 2013, 41, 1407–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddaramappa, S.; Challacombe, J.F.; Petersen, J.M.; Pillai, S.; Kuske, C.R. Comparative analyses of a putative Francisella conjugative element. Genome 2014, 57, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Sharma, A.; Kongshaug, H.; Nilsen, F.; Jonassen, I. Whole genome sequencing of the fish pathogen Francisella noatunensis subsp. orientalis Toba04 gives novel insights into Francisella evolution and pathogenecity. BMC Genom. 2012, 13, 598. [Google Scholar] [CrossRef] [Green Version]
- Rydzewski, K.; Schulz, T.; Brzuszkiewicz, E.; Holland, G.; Luck, C.; Fleischer, J.; Grunow, R.; Heuner, K. Genome sequence and phenotypic analysis of a first German Francisella sp. isolate (W12-1067) not belonging to the species Francisella tularensis. BMC Microbiol. 2014, 14, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jepras, R.I.; Fitzgeorge, R.B.; Baskerville, A. A comparison of virulence of two strains of Legionella pneumophila based on experimental aerosol infection of guinea-pigs. Epidemiol. Infect. 1985, 95, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.; Lochau, P.; Jacob, D.; Heuner, K.; Grunow, R. Successful re-evaluation of broth medium T for the growth of Francisella tularensis ssp. and other higly pathogenic bacteria. J. Microbiol. Methods 2016, 121, 5–7. [Google Scholar] [CrossRef]
- Pavlovich, N.V.; Mishan’kin, B.N. [Transparent nutrient medium for culturing Francisella tularensis]. Antibiot. Med. Biotekhnol. 1987, 32, 133–137. [Google Scholar]
- Edelstein, P.H. Improved semiselective medium for isolation of Legionella pneumophila from contaminated clinical and environmental specimens. J. Clin. Microbiol. 1981, 14, 298–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, W.H.; Egan, J.B. Prophage induction of noninducible coliphage 186. J. Virol. 1974, 14, 1349–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hockett, K.L.; Baltrus, D.A. Use of the Soft-agar Overlay Technique to Screen for Bacterially Produced Inhibitory Compounds. J. Vis. Exp. 2017. [Google Scholar] [CrossRef] [Green Version]
- Schnos, M.; Zahn, K.; Inman, R.B.; Blattner, F.R. Initiation protein induced helix destabilization at the lambda origin: A prepriming step in DNA replication. Cell 1988, 52, 385–395. [Google Scholar] [CrossRef]
- Zimmermann, L.; Stephens, A.; Nam, S.Z.; Rau, D.; Kubler, J.; Lozajic, M.; Gabler, F.; Soding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Adriaenssens, E.; Brister, J.R. How to Name and Classify Your Phage: An Informal Guide. Viruses 2017, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigel, C.; Seitz, H. Bacteriophage replication modules. FEMS Microbiol. Rev. 2006, 30, 321–381. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, H.W. Bacteriophage observations and evolution. Res. Microbiol. 2003, 154, 245–251. [Google Scholar] [CrossRef]
- Kostyuchenko, V.A.; Chipman, P.R.; Leiman, P.G.; Arisaka, F.; Mesyanzhinov, V.V.; Rossmann, M.G. The tail structure of bacteriophage T4 and its mechanism of contraction. Nat. Struct. Mol. Biol. 2005, 12, 810–813. [Google Scholar] [CrossRef]
- Maslowska, K.H.; Makiela-Dzbenska, K.; Fijalkowska, I.J. The SOS system: A complex and tightly regulated response to DNA damage. Environ. Mol. Mutagen. 2019, 60, 368–384. [Google Scholar] [CrossRef] [Green Version]
- Hochschild, A.; Lewis, M. The bacteriophage lambda CI protein finds an asymmetric solution. Curr. Opin. Struct. Biol. 2009, 19, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Wang, I.N.; Smith, D.L.; Young, R. Holins: The protein clocks of bacteriophage infections. Annu. Rev. Microbiol. 2000, 54, 799–825. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.L.; Saier, M.H., Jr. Topological and phylogenetic analyses of bacterial holin families and superfamilies. Biochim. Biophys. Acta 2013, 1828, 2654–2671. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk, B.; Bienkowska-Szewczyk, K.; Kozloff, L.M. Identification of T4 gene 25 product, a component of the tail baseplate, as a 15K lysozyme. Mol. Gen. Genet. 1986, 202, 363–367. [Google Scholar] [CrossRef]
- Cahill, J.; Young, R. Phage Lysis: Multiple Genes for Multiple Barriers. Adv. Virus Res. 2019, 103, 33–70. [Google Scholar] [CrossRef] [PubMed]
- Ratner, H.K.; Weiss, D.S. Francisella novicida CRISPR-Cas Systems Can Functionally Complement Each Other in DNA Defense while Providing Target Flexibility. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef]
- Chen, J.; Novick, R.P. Phage-mediated intergeneric transfer of toxin genes. Science 2009, 323, 139–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakasis, A.; Panitsa, G. Bacteriophage therapy as an alternative treatment for human infections. A comprehensive review. Int. J. Antimicrob. Agents 2019, 53, 16–21. [Google Scholar] [CrossRef]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage Applications for Food Production and Processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courchesne, N.M.; Parisien, A.; Lan, C.Q. Production and application of bacteriophage and bacteriophage-encoded lysins. Recent Pat. Biotechnol. 2009, 3, 37–45. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORF Number | Aa | Motif/Putative Function | putative Function in Phages According to HHpred # (Probability) | BLASTp Viruses (Identity) | BLASTp Francisella Group (Identity *) |
|---|---|---|---|---|---|
| Fhv_0001 (0986) ** | 197 | 2× internal repeat, HP | Major capsid protein (50%) | Aedes pseudoscutellaris reovirus VP6 (25%) | - |
| Fhv_0002 (0987) | 408 | Phage_GPD, GpD phage late control protein D | Tail protein (100%) ° | Wolbachia phage WO (30%) ° | Francisella sp. SYW-9 (47%) |
| Fhv_0003 (0988) | 066 | Phage_tail_X Phage tail protein | - | Wolbachia phage WO (55%) ° | Francisella sp. SYW-9 (55%) |
| Fhv_0004 (0989) | 138 | Phage_P2_GpU Phage tail assembly | Major tube protein gp53 (86%) | Caudovirales phage (29%) ° | Francisella sp. SYW-9 (43%) |
| Fhv_0005 (0990) | 607 | Coiled coil, 3× LCRs Phage tail tape measure protein | Tape Measure Protein gp57 (99%) | Vibrio phage VpKK5 (36%) | Francisella sp. SYW-9 (36%) |
| Fhv_0006 (0991) | 089 | Phage_TAC_7 Phage tail assembly chaperone, Myoviridae | Lambda integrase (48%) DNA-binding protein gp33 (41%) Tail assembly chaperone (38%) | Pseudomonas phage PPpW-3 (31%) ° | Francisella sp. SYW-9 (53%) |
| Fhv_0007 (0992) | 159 | Phage_tube Major tail tube protein | Tail tube protein gp19 (80%) ° | Wolbachia phage WO (35%) ° | Francisella sp. SYW-9 (51%) |
| Fhv_0008 (0993) | 385 | Phae_sheath_1, Phage_sheath_1C Phage tail sheath protein | Tail sheath protein Gp18 (100%) ° | Wolbachia phage WO (49%) ° | Francisella sp. SYW-9 (56%) |
| Fhv_0009 (0994) | 106 | HP | Integrase (21%) | - | - |
| Fhv_0010 (0995) | 081 | HP | Gene 9 protein Knob (23%) | - | - |
| Fhv_0011 (0996) | 066 | LCR, | arc repressor (39%) | Pseudomonas phage EL (50%) ° | F. tularensis subsp. novicida PA10-7858 (50%) |
| Fhv_0012 (0997) | 198 | Phage_base_V, Baseplate assembly protein V | Baseplate assembly protein V (100%) ° | Wolbachia phage WO (34%) ° | Francisella sp. SYW-9 (36%) |
| Fhv_0013 (0998) | 163 | HP | Minor tail protein U (97%) | Wolbachia phage WO (26%) ° | - |
| Fhv_0014 (0999) | 166 | 166 aa, LCR, minor tail_Z superfamily Phage minor tail protein | - | Halomonas virus HAP1 (26%) ° | Francisella sp. SYW-9 (40%) |
| Fhv_0015 (1000) | 111 | HP | Tail attachment protein (95%) | - | Francisella sp. SYW-9 (30%) |
| Fhv_0016 (1001) | 063 | HP | - | Mediterranean phage uvMED (36%) | - |
| Fhv_0017 (1002) | 600 | Peptidase_S78, Phage_capsid Major phage capsid protein | Major capsid protein (100%) | Escherichia phage vB_EcoM_ECO1230-10 (37%) ° | Francisella sp. SYW-9 (40%) |
| Fhv_0018 (1003) | 473 | Phage_portal_2, portal_lambda Phage hole protein, forming DNA-ejection hole | Portal protein (100%) | Enterobacter phage Arya (34%) ° | Francisella sp. SYW-9 (39%) |
| Fhv_0019 (1004) | 088 | HP, coiled coil | Head-to-tail joining protein W (89%) | - | - |
| Fhv_0020 (1005) | 110 | DUF1353, conserved HP | - | Fusobacterium phage Funu2 (36%) ° | F. marina (46%) |
| Fhv_0021 (1006) | 183 | HP, DUF4376 | Tail fiber assembly protein U (79%) ° | - | - |
| Fhv_0022 (1007) | 324 | DUF3751, Pfam_12571 Phage tail fibre protein | Long-tail fiber proximal subunit (75%) ° | Salmonella phage vB_SnwM_CGG4-1 (35%) ° | Francisella sp. SYW-9 (42%) |
| Fhv_0023 (1008) | 195 | Tail_P2_I, Phage tail protein I | Baseplate wedge protein gp6 (77%) ° | Ralstonia phage phiRSP (31%) ° | Francisella sp. SYW-9 (47%) |
| Fhv_0024 (1009) | 281 | Baseplate_J, (P2 phage), gpJ Phage-related baseplate assembly protein | Baseplate wedge protein gp6 (100%) ° | Pseudomonas phage PPpW-3 (36%) ° | Francisella sp. SYW-9 (52%) |
| Fhv_0025 (1010) | 112 | GPW_gp25 T4 phage, V1_zyme Phage baseplate protein, lysozyme activity | Baseplate wedge protein gp25 (100%) ° | Ralstonia phage phiRSP (48%) ° | Francisella sp. SYW-9 (53%) F. philomiragia (37%) |
| Fhv_0026 (1011) | 131 | HP, coiled coil, TM | Fibritin (56%) ° | - | - |
| Fhv_0027 (1012) | 169 | HP, LCR, TM | - | - | F. marina (47%) |
| Fhv_0028 (1013) | 602 | Terminase_GpA, Phage terminase large subunit, DNA packaging | Terminase (100%) | Enterobacter phage Arya (48%) ° | Francisella sp. SYW-9 (59%) |
| Fhv_0029 (1014) | 671 | HP, 6x LCR | - | - | - |
| Fhv_0030 (1015) | 093 | HP | - | - | F. marina (40%) |
| Fhv_0031 (1016) | 172 | Phage_Nu1 SF Minor subunit Nu1 of terminase | Regulatory protein cox (98%) ° | Mediterranean phage uvMED (29%) | F. philomiragia (57%) F. novicida (57%) F. salina (57%) |
| Fhv_0032 (1017) | 128 | HP | Middle operon regulator (100%) ° | - | Francisella novicida (33%) |
| Fhv_0033 (1018) | 422 | Arm-DNA-bind_3, Phage_Int_P4, Phage integrase | Integrase (100%) | Pseudomonas phage phiAH14b (32%) | F. salina (54%) F. novicida (54%) F. philomiragia (56%) Francisella sp. SYW-9 (48%) |
| Fhv_0034 (1019) | 106 | HP, LCR | - | - | - |
| Fhv_0035 (1020) | 072 | HP | - | - | F. marina (70%) |
| Fhv_0036 (n.a.) | 061 | HP | - | Megaviridae environmental sample (41%) | Francisella sp. FSC1006 (43%) F. marina (45%) |
| Fhv_0037 (n.a.) | 075 | HP, TM, conju_TIGR03752, Integrating conjugative element protein | Fusion of phage phi29 Gp7 protein and Cell division protein FtsB (64%) | - | Francisella sp. SYW-9 (50%) |
| Fhv_0038 (1021) | 240 | Phage_pRha, ANT Phage regulatory and anti-repressor protein | Anti-sigma effector (45%) | Lactobacillus phage phiEF-1.1 (66%) | Francisella sp. SYW-9 (54%) F. marina (71%) |
| Fhv_0039 (1022) | 120 | Phage_TIGR01671, YopX Putative phage protein | HP ORF041 (Staphylococcus phage, 100%) | Clostridium phage phiCT19406C (41%) | - |
| Fhv_0040 (1023) | 100 | HP, coiled coil | Long tail fiber distal subunit (24%) ° | - | - |
| Fhv_0041 (1024) | 085 | HP | Regulatory protein cox (99%) | - | Francisella sp. SYW-9 (64%) F. philomiragia (48%) |
| Fhv_0042 (1025) | 212 | Inhibitor_G39P Blocking G40P replicative helicase | Replisome organizer (100%) ° | - | Francisella sp. FSC1006 (46%) F. philomiragia (34%) |
| Fhv_0043 (1026) | 240 | Phg_2220_C, (internal repeat) Phage replication protein | DNA-binding protein TubR (97%) | Lactobacillus prophage Lj771 (50%) ° | Francisella sp. FSC1006 (57%) |
| Fhv_0044 (1027) | 056 | P22_Cro (lytic growth), Cro protein, phage_CI_repressor | Repressor protein (98%) | Erwinia phage vB_EhrS_59 (45%) | Francisella sp. FSC1006 (45%) |
| Fhv_0045 (1028) | 265 | Peptidase_S24_S26 SF, CI-type HTH_XRE domain, Pro-phage repressor (CI), LexA protein domain, | Lambda Repressor (100%) | Streptococcus phage PH15 (48%) | F. philomiragia (54%) F. salina (52%) F. marina (51%) F. novicida (57%) |
| Fhv_0046 (1029) | 293 | HP | - | Marinobacter phage AS1 (23%) | - |
| Fhv_0047 (1030) | 112 | HP, DUF4325 | - | Campylobacter phage CP30A (27%) ° | - |
| Fhv_0048(n.a.) | 125 | HP, TM | - | - | - |
| Fhv_0049 (1031) | 264 | HP | - | - | F. philomiragia (45%) F. noatunensis (45%) |
| Fhv_0050 (1032) | 130 | TM, bPH_2 Putative transmembrane protein | Endolysin (75%) ° | Serratia phage phiMAM1 (39%) | Francisella sp. W12-1067 (46%) |
| Fhv_0051 (1033) | 375 | Phage_integrase, Arm-DNA-bind_3 Site-spezific integrase/recombinase | Integrase (100%) | Prokaryotic dsDNA virus sp. (30%) | F. adeliensis (62%) F. philomiragia (56%) F. marina (52%) F. salina (51% )Francisella sp. SYW-9 (49%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Köppen, K.; Prensa, G.I.; Rydzewski, K.; Tlapák, H.; Holland, G.; Heuner, K. First Description of a Temperate Bacteriophage (vB_FhiM_KIRK) of Francisella hispaniensis Strain 3523. Viruses 2021, 13, 327. https://doi.org/10.3390/v13020327
Köppen K, Prensa GI, Rydzewski K, Tlapák H, Holland G, Heuner K. First Description of a Temperate Bacteriophage (vB_FhiM_KIRK) of Francisella hispaniensis Strain 3523. Viruses. 2021; 13(2):327. https://doi.org/10.3390/v13020327
Chicago/Turabian StyleKöppen, Kristin, Grisna I. Prensa, Kerstin Rydzewski, Hana Tlapák, Gudrun Holland, and Klaus Heuner. 2021. "First Description of a Temperate Bacteriophage (vB_FhiM_KIRK) of Francisella hispaniensis Strain 3523" Viruses 13, no. 2: 327. https://doi.org/10.3390/v13020327
APA StyleKöppen, K., Prensa, G. I., Rydzewski, K., Tlapák, H., Holland, G., & Heuner, K. (2021). First Description of a Temperate Bacteriophage (vB_FhiM_KIRK) of Francisella hispaniensis Strain 3523. Viruses, 13(2), 327. https://doi.org/10.3390/v13020327