
Efficacy of Oral Vaccine against Classical Swine Fever in Wild Boar and Estimation of the Disease Dynamics in the Quantitative Approach

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Wild Boar Data and Data Sources
2.2. Wild Boar Samples
2.3. RT-qPCR
2.4. Luciferase-Based SNT
2.5. Regression Analysis of Antibody Detection
2.6. Temporal Trend Analysis of Antibody Responses in Wild Boars Population
2.7. Classification of Quantitative Values
2.8. Multi-Distance Spatial Cluster Analysis and Kernel Density Analysis
3. Results
3.1. Quantitative Detection of Viral RNA in Wild Boar Serum
3.2. Detection of Antibodies against CSFV
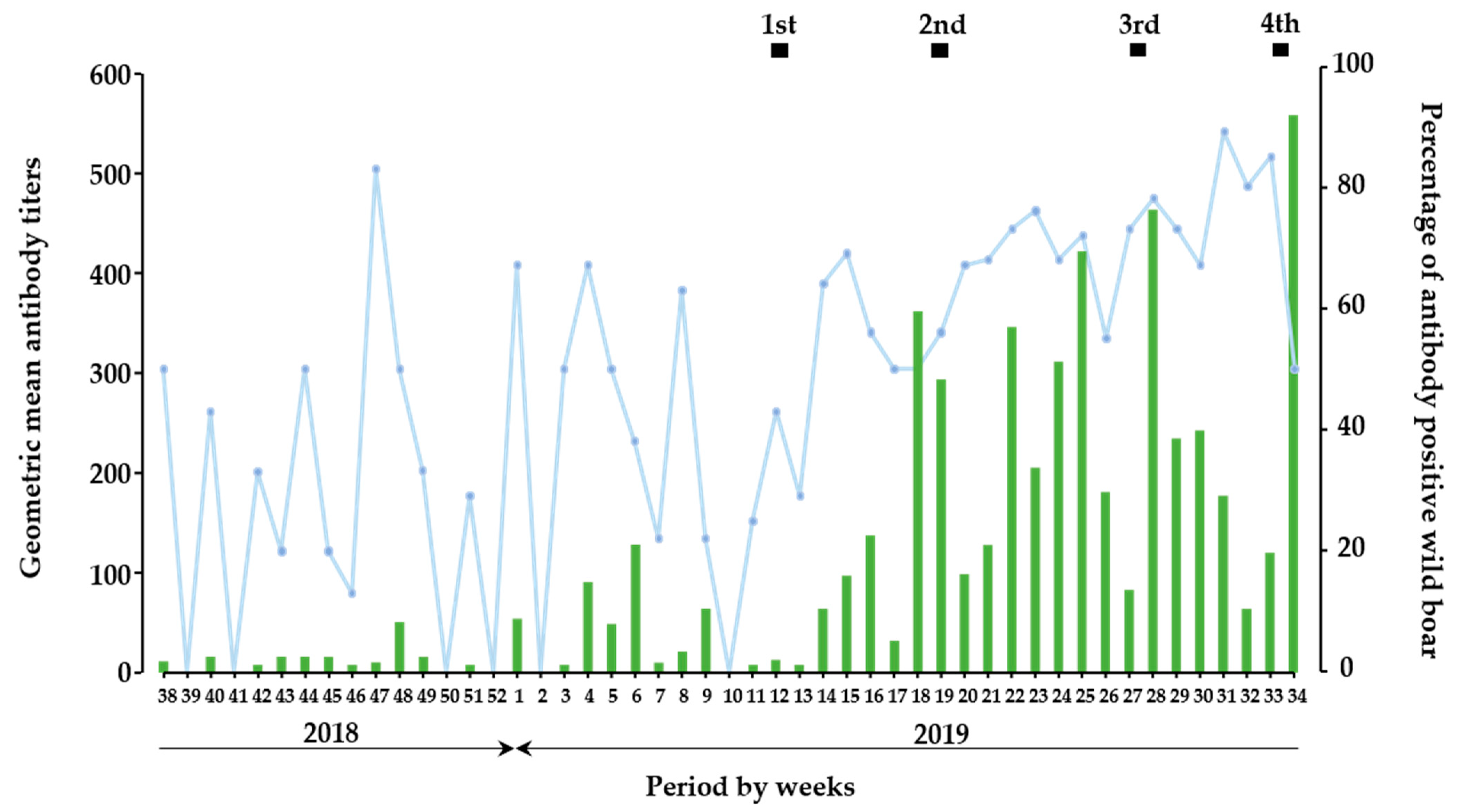
3.3. Temporal Trend Analysis of Antibody Response in Wild Boar during With- and Without-Vaccination Period
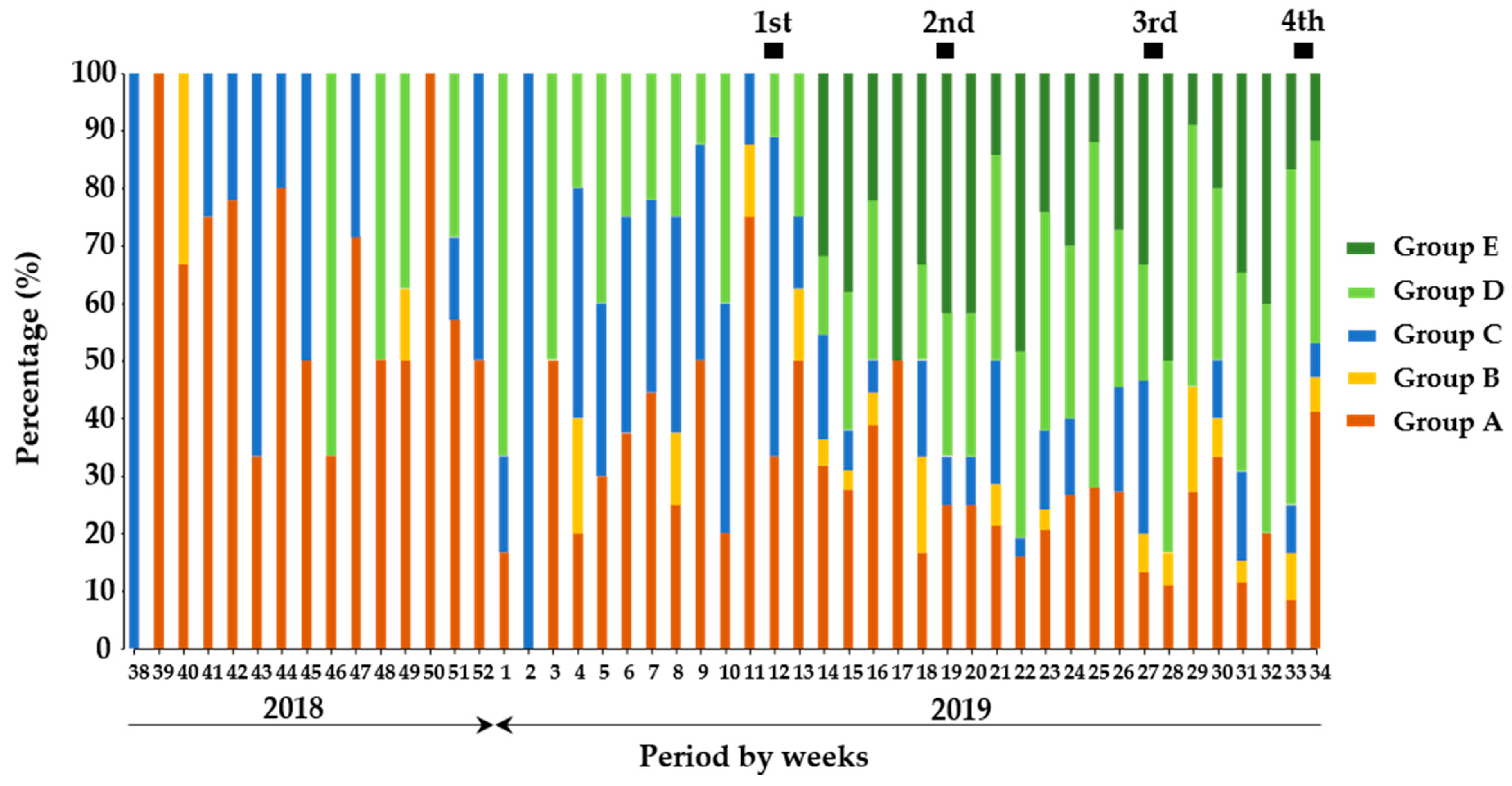
3.4. Divergent Quantitative Classifications among Wild Boar Population in Gifu Prefecture
3.5. Proportional Distribution of Diversity of Wild Boars
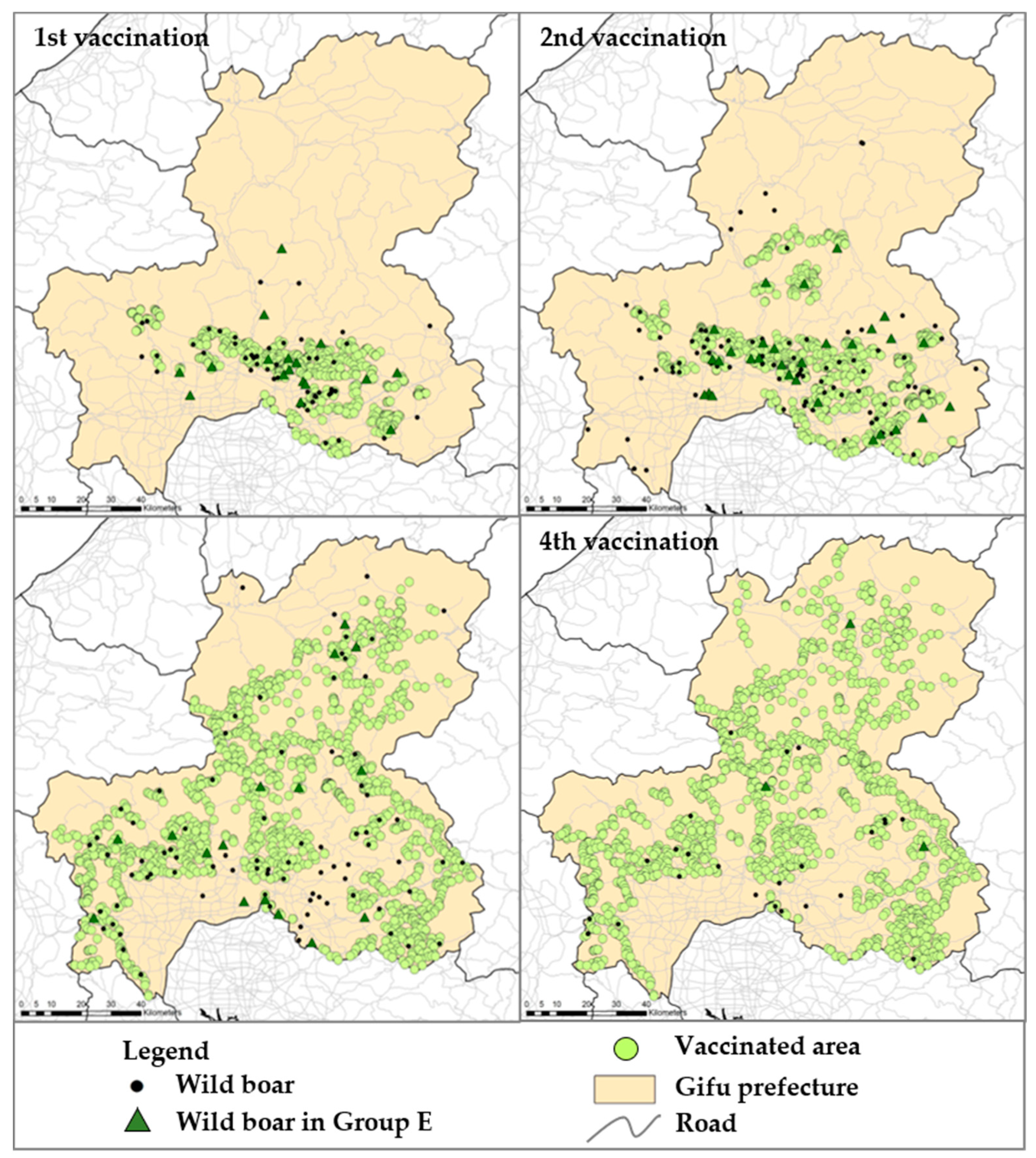
3.6. Efficacy of Oral Vaccine for Wild Boar
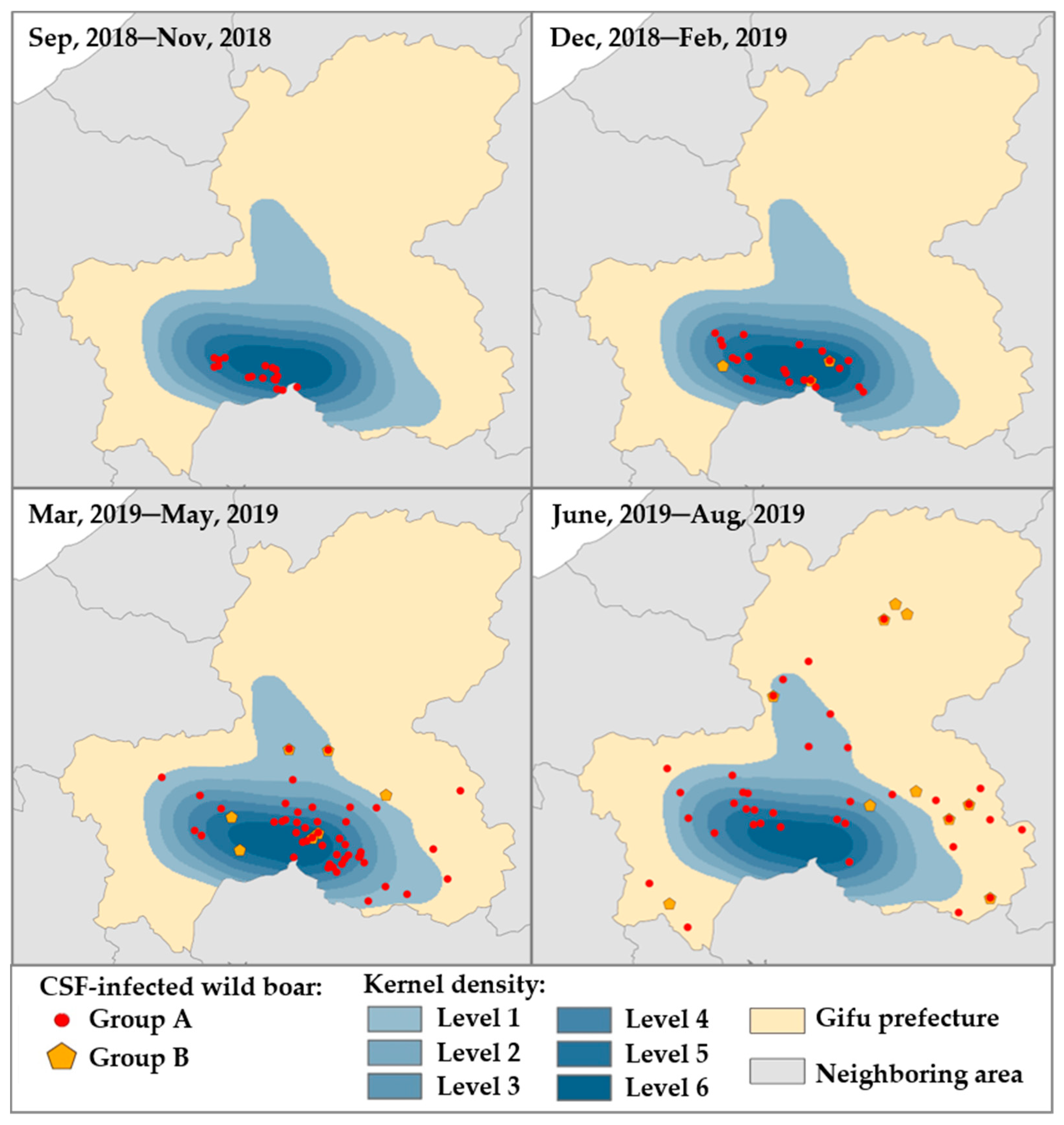
3.7. Kernel Density Estimation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Statements
References
- World Organization for Animal Health. Information on Aquatic and Terrestrial Animal Diseases: Classical Swine Fever (CSF). 2020. Available online: https://www.oie.int/en/animal-health-in-the-world/animal-diseases/Classical-swine-fever/ (accessed on 19 December 2020).
- Ganges, L.; Crooke, H.R.; Bohorquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical swine fever virus: The past, present and future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef] [PubMed]
- Cabezon, O.; Colom-Cadena, A.; Munoz-Gonzalez, S.; Perez-Simo, M.; Bohorquez, J.A.; Rosell, R.; Marco, I.; Domingo, M.; Lavin, S.; Ganges, L. Post-Natal Persistent Infection With Classical Swine Fever Virus in Wild Boar: A Strategy for Viral Maintenance? Transbound Emerg. Dis. 2017, 64, 651–655. [Google Scholar] [CrossRef]
- Shimizu, Y.; Hayama, Y.; Murato, Y.; Sawai, K.; Yamaguchi, E.; Yamamoto, T. Epidemiology of classical swine fever in Japan-a descriptive analysis of the outbreaks in 2018–2019. Front Vet. Sci. 2020, 7, 573480. [Google Scholar] [CrossRef]
- Kameyama, K.I.; Nishi, T.; Yamada, M.; Masujin, K.; Morioka, K.; Kokuho, T.; Fukai, K. Experimental infection of pigs with a classical swine fever virus isolated in Japan for the first time in 26 years. J. Vet. Med. Sci. 2019, 81, 1277–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choe, S.; Cha, R.M.; Yu, D.S.; Kim, K.S.; Song, S.; Choi, S.H.; Jung, B.I.; Lim, S.I.; Hyun, B.H.; Park, B.K.; et al. Rapid Spread of Classical Swine Fever Virus among South Korean Wild Boars in Areas near the Border with North Korea. Pathogens 2020, 9, 244. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Ji, S.; Liu, Y.; Lei, J.L.; Xia, S.L.; Wang, Y.; Du, M.L.; Shao, L.; Meng, X.Y.; Zhou, M.; et al. Isolation and characterization of a moderately virulent classical swine fever virus emerging in China. Transbound Emerg. Dis. 2017, 64, 1848–1857. [Google Scholar] [CrossRef]
- Abe, M. External measurements of the Japanese wild boar (Sus scrofa leucomystax Temminck). J. Mamm. Soc. Jpn. 1986, 11, 147–154. [Google Scholar]
- Geocoding. Available online: https://www.geocoding.jp/ (accessed on 26 November 2020).
- Vilcek, S.; Herring, A.J.; Herring, J.A.; Nettleton, P.F.; Lowings, J.P.; Paton, D.J. Pestiviruses isolated from pigs, cattle and sheep can be allocated into at least three genogroups using polymerase chain reaction and restriction endonuclease analysis. Arch. Virol. 1994, 136, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Sakoda, Y.; Wakamoto, H.; Tamura, T.; Nomura, T.; Naito, M.; Aoki, H.; Morita, H.; Kida, H.; Fukusho, A. Development and evaluation of indirect enzyme-linked immunosorbent assay for a screening test to detect antibodies against classical swine fever virus. Jpn. J. Vet. Res. 2012, 60, 85–94. [Google Scholar]
- Hoffmann, B.; Beer, M.; Schelp, C.; Schirrmeier, H.; Depner, K. Validation of a real-time RT-PCR assay for sensitive and specific detection of classical swine fever. J. Virol. Methods 2005, 130, 36–44. [Google Scholar] [CrossRef]
- Meyer, D.; Petrov, A.; Becher, P. Inactivation of Classical Swine Fever Virus in Porcine Serum Samples Intended for Antibody Detection. Pathogens 2019, 8, 286. [Google Scholar] [CrossRef] [Green Version]
- Tetsuo, M.; Matsuno, K.; Tamura, T.; Fukuhara, T.; Kim, T.; Okamatsu, M.; Tautz, N.; Matsuura, Y.; Sakoda, Y. Development of a High-Throughput Serum Neutralization Test Using Recombinant Pestiviruses Possessing a Small Reporter Tag. Pathogens 2020, 9, 188. [Google Scholar] [CrossRef] [Green Version]
- Prabhakaran, S. R-Statistics. Available online: http://r-statistics.co/ (accessed on 20 December 2020).
- Isoda, N.; Baba, K.; Ito, S.; Ito, M.; Sakoda, Y.; Makita, K. Dynamics of Classical Swine Fever Spread in Wild Boar in 2018–2019, Japan. Pathogens 2020, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Ministry of Agriculture, Forestry and Fisheries. Guidelines for Field Spraying of CSF Oral Vaccine. 2019. Available online: https://www.maff.go.jp/j/syouan/douei/csf/attach/pdf/domestic-109.pdf (accessed on 11 December 2020).
- Kodera, Y.C.S.F. Prevention of Epidemics from a Point of View of the Ecology of Wild Boar. J. Vet. Epidemiol. 2019, 23, 4–8. [Google Scholar] [CrossRef] [Green Version]
- Institute, E.S.R. Multi-Distance Spatial Cluster Analysis (Ripley’s K Function). Available online: https://pro.arcgis.com/en/pro-app/tool-reference/spatial-statistics/multi-distance-spatial-cluster-analysis.htm (accessed on 4 December 2020).
- Ito, S.; Jurado, C.; Bosch, J.; Ito, M.; Sanchez-Vizcaino, J.M.; Isoda, N.; Sakoda, Y. Role of wild boar in the spread of classical swine fever in Japan. Pathogens 2019, 8, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaden, V.; Hänel, A.; Renner, C.; Gossger, K. Oral immunisation of wild boar against classical swine fever in Baden-Württemberg: Development of the seroprevalences based on the hunting bag. Eur. J. Wildl. Res. 2005, 51, 101–107. [Google Scholar] [CrossRef]
- World Organization for Animal Health. Classical Swine Fever. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. 2019. Available online: https://www.oie.int/standard-setting/terrestrial-manual/access-online/ (accessed on 20 November 2020).
- Kaden, V.; Lange, E.; Kuster, H.; Muller, T.; Lange, B. An update on safety studies on the attenuated “RIEMSER Schweinepestoralvakzine” for vaccination of wild boar against classical swine fever. Vet. Microbiol. 2010, 143, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Moennig, V.; Floegel-Niesmann, G.; Greiser-Wilke, I. Clinical signs and epidemiology of classical swine fever: A review of new knowledge. Vet. J. 2003, 165, 11–20. [Google Scholar] [CrossRef]
- Fritzemeier, J.; Teuffert, J.; Greiser-Wilke, I.; Staubach, C.; Schluter, H.; Moennig, V. Epidemiology of classical swine fever in Germany in the 1990s. Vet. Microbiol. 2000, 77, 29–41. [Google Scholar] [CrossRef]
- Coronado, L.; Bohorquez, J.A.; Munoz-Gonzalez, S.; Perez, L.J.; Rosell, R.; Fonseca, O.; Delgado, L.; Perera, C.L.; Frias, M.T.; Ganges, L. Investigation of chronic and persistent classical swine fever infections under field conditions and their impact on vaccine efficacy. BMC Vet. Res. 2019, 15, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Staubach, C.; Blome, S.; Guberti, V.; Thulke, H.H.; Vos, A.; Koenen, F.; Le Potier, M.F. Controlling of CSFV in European wild boar using oral vaccination: A review. Front. Microbiol. 2015, 6, 1141. [Google Scholar] [CrossRef] [PubMed]
- An, D.J.; Lim, S.I.; Choe, S.; Kim, K.S.; Cha, R.M.; Cho, I.S.; Song, J.Y.; Hyun, B.H.; Park, B.K. Evolutionary dynamics of classical swine fever virus in South Korea: 1987–2017. Vet. Microbiol. 2018, 225, 79–88. [Google Scholar] [CrossRef]
- Hayama, Y.; Shimizu, Y.; Murato, Y.; Sawai, K.; Yamamoto, T. Estimation of infection risk on pig farms in infected wild boar areas-epidemiological analysis for the reemergence of classical swine fever in Japan in 2018. Prev. Vet. Med. 2020, 175, 104873. [Google Scholar] [CrossRef]
- Fukai, K.; Nishi, T.; Yamada, M.; Ikezawa, M. Toward better control of classical swine fever in wild boars: Susceptibility of boar-pig hybrids to a recent Japanese isolate and effectiveness of a bait vaccine. Vet. Res. 2020, 51, 96–103. [Google Scholar] [CrossRef]
- Kaden, V.; Lange, B. Oral immunisation against classical swine fever (CSF): Onset and duration of immunity. Vet. Microbiol. 2001, 82, 301–310. [Google Scholar] [CrossRef]
- Postel, A.; Austermann-Busch, S.; Petrov, A.; Moennig, V.; Becher, P. Epidemiology, diagnosis and control of classical swine fever: Recent developments and future challenges. Transbound Emerg. Dis. 2017, 65, 248–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saïd, S.; Tolon, V.; Brandt, S.; Baubet, E. Sex effect on habitat selection in response to hunting disturbance: The study of wild boar. Eur. J. Wildl. Res. 2011, 58, 107–115. [Google Scholar] [CrossRef]
- Boitani, L.; Mattei, L.; Nonis, D.; Corsi, F. Spatial and activity patterns of wild boars in Tuscany, Italy. J. Mammal. 1994, 75, 600–612. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age Group | Total | Gender | Status | |||
|---|---|---|---|---|---|---|
| Male | Female | Unknown | Captured | Dead | ||
| ≤1 year | 540 | 271 | 266 | 3 | 478 | 62 |
| 1–2 years | 360 | 176 | 181 | 3 | 328 | 32 |
| >2 years | 534 | 302 | 231 | 1 | 463 | 71 |
| Unknown | 9 | 5 | 4 | 0 | 9 | 0 |
| Total | 1443 | 754 | 682 | 7 | 1278 | 165 |
| Groups | Cutoff Values | Total | Age | Status | |||||
|---|---|---|---|---|---|---|---|---|---|
| RT-qPCR | SNT | ≤1 | 1–2 | >2 | Un- known | Captured | Dead | ||
| Group A | <30 | <8 | 168 | 57 | 45 | 66 | 0 | 130 | 38 |
| Group B | <30 | 8–32 | 23 | 11 | 5 | 7 | 0 | 16 | 7 |
| Group C | 30–40 | ≤128 | 81 | 18 | 22 | 41 | 0 | 54 | 27 |
| Group D | ≥30 | >128 | 187 | 24 | 62 | 101 | 0 | 185 | 2 |
| Group E | ≥40 | 8–128 | 83 | 15 | 27 | 41 | 0 | 78 | 5 |
| No infection | ≥40 | <8 | 624 | 329 | 130 | 156 | 9 | 606 | 18 |
| Total | 1166 | 454 | 291 | 412 | 9 | 1069 | 97 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazarragchaa, E.; Isoda, N.; Kim, T.; Tetsuo, M.; Ito, S.; Matsuno, K.; Sakoda, Y. Efficacy of Oral Vaccine against Classical Swine Fever in Wild Boar and Estimation of the Disease Dynamics in the Quantitative Approach. Viruses 2021, 13, 319. https://doi.org/10.3390/v13020319
Bazarragchaa E, Isoda N, Kim T, Tetsuo M, Ito S, Matsuno K, Sakoda Y. Efficacy of Oral Vaccine against Classical Swine Fever in Wild Boar and Estimation of the Disease Dynamics in the Quantitative Approach. Viruses. 2021; 13(2):319. https://doi.org/10.3390/v13020319
Chicago/Turabian StyleBazarragchaa, Enkhbold, Norikazu Isoda, Taksoo Kim, Madoka Tetsuo, Satoshi Ito, Keita Matsuno, and Yoshihiro Sakoda. 2021. "Efficacy of Oral Vaccine against Classical Swine Fever in Wild Boar and Estimation of the Disease Dynamics in the Quantitative Approach" Viruses 13, no. 2: 319. https://doi.org/10.3390/v13020319
APA StyleBazarragchaa, E., Isoda, N., Kim, T., Tetsuo, M., Ito, S., Matsuno, K., & Sakoda, Y. (2021). Efficacy of Oral Vaccine against Classical Swine Fever in Wild Boar and Estimation of the Disease Dynamics in the Quantitative Approach. Viruses, 13(2), 319. https://doi.org/10.3390/v13020319