
Transmission of Bluetongue Virus Serotype 8 by Artificial Insemination with Frozen–Thawed Semen from Naturally Infected Bulls

 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. BTV-8 Contaminated Semen
2.3. Sperm Quality Assessment
2.4. Study Design
2.4.1. Oestrus Synchronization and Artificial Insemination
2.4.2. Experiment 1
2.4.3. Experiment 2
2.4.4. Pregnancy Diagnosis and Follow-up
2.4.5. Sampling and Clinical Examination
2.4.6. Environmental Monitoring and Vector Control
2.5. Bluetongue Virus Laboratory Diagnosis
2.5.1. BTV Genome Detection
2.5.2. BTV Antibody Detection
2.5.3. BTV Isolation and Titration
2.6. Foetal Examination
2.7. Statistical Analysis
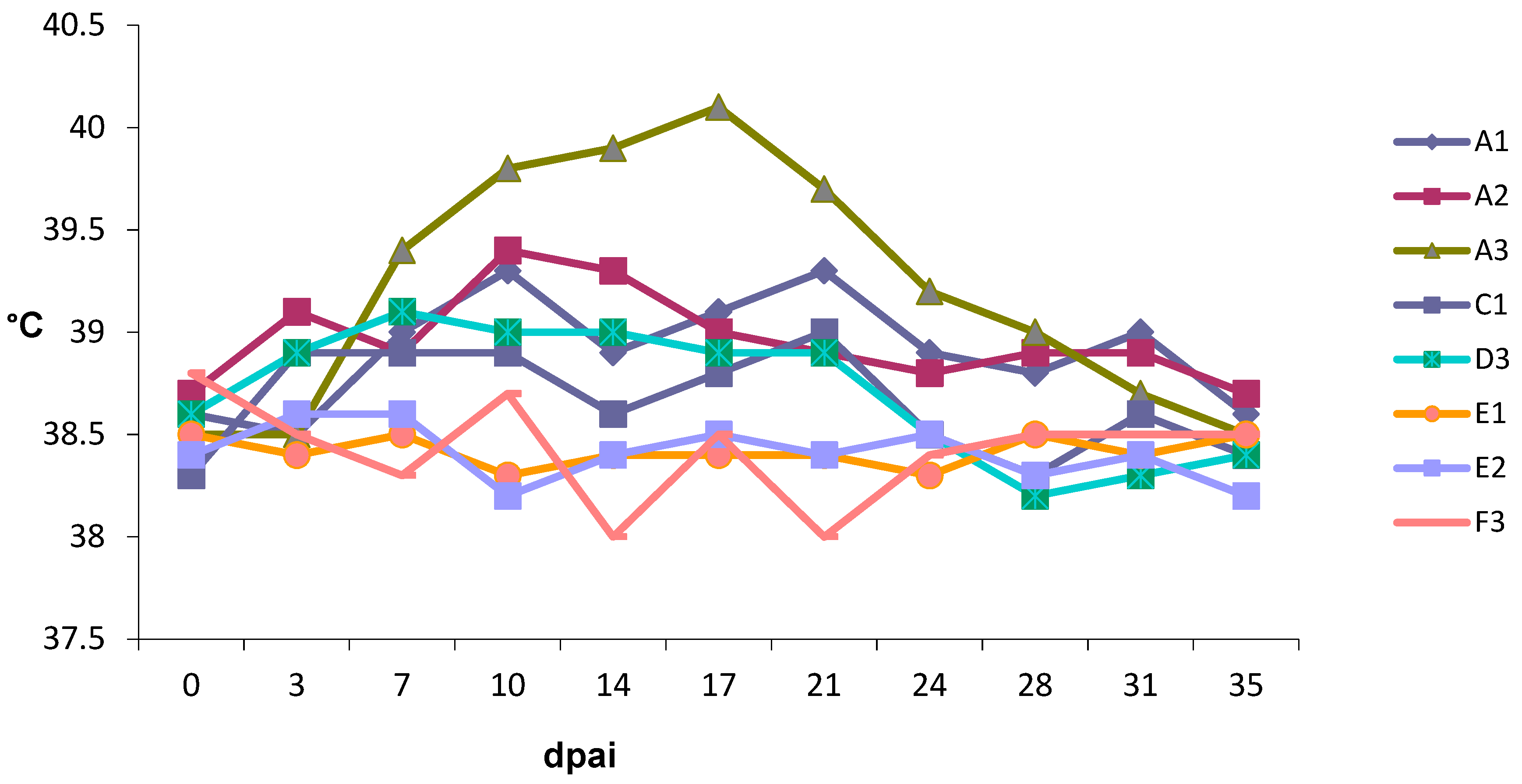
3. Results
3.1. Animals
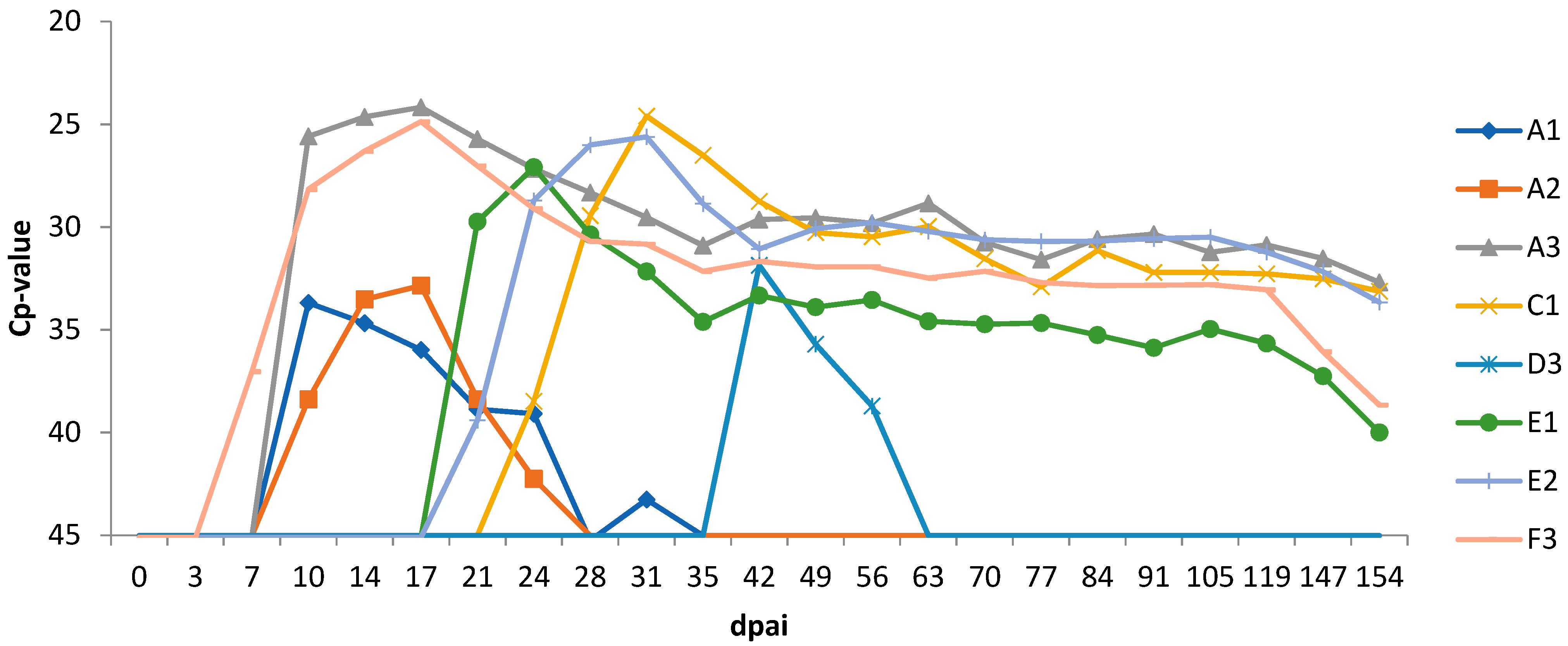
3.2. Real-Time RT-PCR Analysis and Viral Titration of the Semen Batches
3.3. Sperm Quality Assessment
3.4. Experiment 1
3.5. Experiment 2
3.6. Analysis of the Culicoides Trapped
3.7. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mertens, P.P.; Diprose, J.; Maan, S.; Singh, K.P.; Attoui, H.; Samuel, A.R. Bluetongue virus replication, molecular and structural biology. Vet. Ital. 2004, 40, 426–437. [Google Scholar]
- Barrat-Boyes, S.M.; MacLachlan, N.J. Dynamics of viral spread in bluetongue virus infected calves. Vet. Microbiol. 1994, 40, 361–371. [Google Scholar] [CrossRef]
- Orange, J.P.; Dinh, E.T.N.; Goodfriend, O.; Citino, S.B.; Wisely, S.M.; Blackburn, J.K. Evidence of epizootic hemorrhagic disease virus and bluetongue virus exposure in nonnative ruminant species in northern Florida. J. Zoo Wildl. Med. 2021, 51, 745–751. [Google Scholar] [CrossRef]
- Bonneau, K.R.; DeMaula, C.; Mullens, B.; MacLachlan, N.J. Duration of viraemia infectious to Culicoides sonorensis in bluetongue virus-infected cattle and sheep. Vet. Microbiol. 2002, 88, 115–125. [Google Scholar] [CrossRef]
- Linden, A.; Gregoire, F.; Nahayo, A.; Hanrez, D.; Mousset, B.; Massart, A.L.; De Leeuw, I.; Vandemeulebroucke, E.; Vandenbussche, F.; De Clercq, K. Bluetongue virus in wild deer, Belgium, 2005–2008. Emerg. Infect. Dis. 2010, 16, 833–836. [Google Scholar] [CrossRef] [PubMed]
- López-Olvera, J.R.; Falconi, C.; Férnandez-Pacheco, P.; Fernández-Pinero, J.; Sánchez, M.A.; Palma, A.; Herruzo, I.; Vicente, J.; Jiménez-Clavero, M.A.; Arias, M.; et al. Experimental infection of European red deer (Cervus elaphus) with bluetongue virus serotype 1 and 8. Vet. Microbiol. 2010, 145, 148–152. [Google Scholar] [CrossRef] [Green Version]
- Gethmann, J.; Probst, C.; Conraths, F.J. Economic Impact of a Bluetongue Serotype 8 Epidemic in Germany. Front. Vet. Sci. 2020, 14, 65. [Google Scholar] [CrossRef]
- World Organization for Animal Health (OIE). Terrestrial Animal Health Code, Chapter 8.3: Infection with Bluetongue Virus. 2020. Available online: https://www.oie.int/index.php?id=169&L=0&htmfile=chapitre_bluetongue.htm (accessed on 22 January 2021).
- Toussaint, J.F.; Sailleau, C.; Mast, J.; Houdart, P.; Czaplicki, G.; Demeestere, L.; Vandenbussche, F.; Van Dessel, W.; Goris, N.; Breard, E.; et al. Bluetongue in Belgium, 2006. Emerg. Infect. Dis. 2007, 13, 614–616. [Google Scholar] [CrossRef] [PubMed]
- Saegerman, C.; Berkvens, D.; Mellor, P.S. Bluetongue epidemiology in the European Union. Emerg. Infect. Dis. 2008, 14, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Zientara, S.; MacLachlan, N.J.; Calistri, P.; Sanchez-Vizcaino, J.M.; Savini, G. Bluetongue vaccination in Europe. Expert Rev. Vaccines 2010, 9, 989–991. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.; Carpenter, S.; Gloster, J.; Mellor, P. Re-emergence of bluetongue in northern Europe in 2007. Vet. Rec. 2007, 161, 487–489. [Google Scholar] [CrossRef]
- Backx, A.; Heutink, R.; van Rooij, E.; van Rijn, P. Transplacental and oral transmission of Wild-type bluetongue virus serotype 8 in cattle after experimental infection. Vet. Microbiol. 2009, 138, 235–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méroc, E.; Herr, C.; Verheyden, B.; Hooyberghs, J.; Houdart, P.; Raemaekers, M.; Vandenbussche, F.; De Clercq, K.; Mintiens, K. Bluetongue in Belgium: Episode II. Transbound. Emerg. Dis. 2009, 56, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Vangeel, I.; De Leeuw, I.; Méroc, E.; Vandenbussche, F.; Riocreux, F.; Hooyberghs, J.; Raemaekers, M.; Houdart, P.; Van der Stede, Y.; De Clercq, K. Bluetongue sentinel surveillance program and cross-sectional serological survey in cattle in Belgium in 2010–2011. Prev. Vet. Med. 2012, 106, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Sailleau, C.; Bréard, E.; Viarouge, C.; Vitour, D.; Romey, A.; Garnier, A.; Fablet, A.; Lowenski, S.; Gorna, K.; Caignard, G.; et al. Re-Emergence of Bluetongue Virus Serotype 8 in France, 2015. Transbound. Emerg. Dis. 2017, 64, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- European Commission, DG Health and Consumers, Animal Health and Welfare. Bluetongue, Control Measures. Available online: https://ec.europa.eu/food/sites/food/files/animals/docs/ad_control-measures_bt_restrictedzones.pdf (accessed on 2 February 2021).
- Courtejoie, N.; Durand, B.; Bournez, L.; Gorlier, A.; Bréard, E.; Sailleau, C.; Vitour, D.; Zientara, S.; Baurier, F.; Gourmelen, C.; et al. Circulation of bluetongue virus 8 in French cattle, before and after the re-emergence in 2015. Transbound. Emerg. Dis. 2018, 65, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Bréard, E.; Sailleau, C.; Quenault, H.; Lucas, P.; Viarouge, C.; Touzain, F.; Fablet, A.; Vitour, D.; Attoui, H.; Zientara, S.; et al. Complete Genome Sequence of Bluetongue Virus Serotype 8, Which Reemerged in France in August 2015. Genome Announc. 2016, 4, e00163-16. [Google Scholar] [CrossRef] [Green Version]
- Pascall, D.J.; Nomikou, K.; Bréard, E.; Zientara, S.; Filipe, A.D.S.; Hoffmann, B.; Jacquot, M.; Singer, J.B.; De Clercq, K.; Bøtner, A.; et al. “Frozen evolution” of an RNA virus suggests accidental release as a potential cause of arbovirus re-emergence. PLoS Biol. 2020, 18, e3000673. [Google Scholar] [CrossRef]
- Thiry, E.; Saegerman, C.; Guyot, H.; Kirten, P.; Losson, B.; Rollin, F.; Bodmer, M.; Czaplicki, G.; Toussaint, J.F.; De Clercq, K.; et al. Bluetongue in northern Europe. Vet. Rec. 2006, 159, 327. [Google Scholar] [CrossRef] [PubMed]
- Elbers, A.R.; Backx, A.; Mintiens, K.; Gerbier, G.; Staubach, C.; Hendrickx, G.; van der Spek, A. Field observations during the Bluetongue serotype 8 epidemic in 2006. II. Morbidity and mortality rate, case fatality and clinical recovery in sheep and cattle in the Netherlands. Prev. Vet. Med. 2008, 87, 31–40. [Google Scholar] [CrossRef]
- Zanella, G.; Martinelle, L.; Guyot, H.; Mauroy, A.; De Clercq, K.; Saegerman, C. Clinical pattern characterization of cattle naturally infected by BTV-8. Transbound. Emerg. Dis. 2013, 60, 231–237. [Google Scholar] [CrossRef]
- De Clercq, K.; De Leeuw, I.; Verheyden, B.; Vandemeulebroucke, E.; Vanbinst, T.; Herr, C.; Meroc, E.; Bertels, G.; Steurbaut, N.; Miry, C.; et al. Transplacental infection and apparently immunotolerance induced by a wild-type bluetongue virus serotype 8 natural infection. Transbound. Emerg. Dis. 2008, 55, 352–359. [Google Scholar] [CrossRef]
- Menzies, F.D.; McCullough, S.J.; McKeown, I.M.; Forster, J.L.; Jess, S.; Batten, C.; Murchie, A.K.; Gloster, J.; Fallows, J.G.; Pelgrim, W.; et al. Evidence for transplacental and contact transmission of bluetongue virus in cattle. Vet. Rec. 2008, 163, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saegerman, C.; Bolkaerts, B.; Baricalla, C.; Raes, M.; Wiggers, L.; de Leeuw, I.; Vandenbussche, F.; Zimmer, J.Y.; Haubruge, E.; Cassart, D.; et al. The impact of naturally-occurring, trans-placental bluetongue virus serotype-8 infection on reproductive performance in sheep. Vet. J. 2011, 187, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Richardson, C.; Taylor, W.P.; Terlecki, S.; Gibbs, E.P.J. Observations on transplacental infection with bluetongue virus in sheep. Am. J. Vet. Res. 1985, 46, 1912–1922. [Google Scholar]
- Richards, W.P.; Crenshaw, G.L.; Bushnell, R.B. Hydranencephaly of calves associated with natural bluetongue virus infection. Cornell Vet. 1971, 61, 336–348. [Google Scholar] [PubMed]
- Vercauteren, G.; Miry, C.; Vandenbussche, F.; Ducatelle, R.; Van der Heyden, S.; Vandemeulebroucke, E.; De Leeuw, I.; Deprez, P.; Chiers, K.; De Clercq, K. Bluetongue virus serotype 8-associated congenital hydranencephaly in calves. Transbound. Emerg. Dis. 2008, 55, 293–298. [Google Scholar] [CrossRef]
- Desmecht, D.; Bergh, R.V.; Sartelet, A.; Leclerc, M.; Mignot, C.; Misse, F.; Sudraud, C.; Berthemin, S.; Jolly, S.; Mousset, B.; et al. Evidence for transplacental transmission of the current wild-type strain of bluetongue virus serotype 8 in cattle. Vet. Rec. 2008, 163, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Darpel, K.E.; Batten, C.A.; Veronesi, E.; Williamson, S.; Anderson, P.; Dennison, M.; Clifford, S.; Smith, C.; Philips, L.; Bidewell, C.; et al. Transplacental transmission of bluetongue virus 8 in cattle, UK. Emerg. Infect. Dis. 2009, 15, 2025–2028. [Google Scholar] [CrossRef] [PubMed]
- Vinomack, C.; Rivière, J.; Bréard, E.; Viarouge, C.; Postic, L.; Zientara, S.; Vitour, D.; Belbis, G.; Spony, V.; Pagneux, C.; et al. Clinical cases of Bluetongue serotype 8 in calves in France in the 2018–2019 winter. Transbound. Emerg. Dis. 2020, 67, 1401–1405. [Google Scholar] [CrossRef]
- Vanbinst, T.; Vandenbussche, F.; Dernelle, E.; De Clercq, K. A duplex real-time RT-PCR for the detection of bluetongue virus in bovine semen. J. Virol. Methods 2010, 169, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Leemans, J.; Raes, M.; Vanbinst, T.; De Clercq, K.; Saegerman, C.; Kirschvink, N. Viral RNA load in semen from bluetongue serotype 8-infected rams: Relationship with sperm quality. Vet. J. 2012, 192, 304–310. [Google Scholar] [CrossRef]
- Kirschvink, N.; Raes, M.; Saegerman, C. Impact of a natural bluetongue serotype 8 infection on semen quality of Belgian rams in 2007. Vet. J. 2009, 182, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Müller, U.; Kemmerling, K.; Straet, D.; Janowitz, U.; Sauerwein, H. Effects of bluetongue virus infection on sperm quality in bulls: A preliminary report. Vet. J. 2009, 186, 402–403. [Google Scholar] [CrossRef]
- Bowen, R.; Howard, T.; Pickett, B. Seminal shedding of bluetongue virus in experimentally infected bulls. Prog. Clin. Biol. Res. 1985, 178, 91–96. [Google Scholar]
- Thomas, F.C.; Singh, E.; Hare, W. Embryo transfer as a means of controlling viral infections. VI. Bluetongue-virus free calves from infectious semen. Theriogenology 1985, 24, 345–350. [Google Scholar] [CrossRef]
- Kirkland, P.D.; Hawkes, R.A. A comparison of laboratory and ‘wild’ strains of bluetongue virus-is there any difference and does it matter? Vet. Ital. 2004, 40, 448–454. [Google Scholar]
- Kirkland, P.D.; Melville, L.; Hunt, N.; Williams, C.; Davis, R. Excretion of bluetongue virus in cattle semen: A feature of laboratory-adapted virus. Vet. Ital. 2004, 40, 497–501. [Google Scholar]
- Savini, G.; Lorusso, A.; Paladini, C.; Migliaccio, P.; Di Gennaro, A.; Di Provvido, A.; Scacchia, A.M.; Monaco, F. Bluetongue serotype 2 and 9 modified live vaccine viruses as causative agents of abortion in livestock: A retrospective analysis in Italy. Transbound. Emerg. Dis. 2014, 61, 69–74. [Google Scholar] [CrossRef]
- Napp, S.; Allepuz, A.; Garcia-Bocanegra, I.; Alba, A.; Vilar, M.J.; Casal, J. Quantitative assessment of the probability of bluetongue virus transmission by bovine semen and effectiveness of preventive measures. Theriogenology 2011, 75, 920–932. [Google Scholar] [CrossRef]
- Hoflack, G.; Opsomer, G.; Rijsselaere, T.; Van Soom, A.; Maes, D.; de Kruif, A.; Duchateau, L. Comparison of computer-assisted sperm motility analysis parameters in semen from belgian blue and Holstein-Friesian bulls. Reprod. Dom. Anim. 2007, 42, 153–161. [Google Scholar] [CrossRef]
- Beckers, J.-F.; Ballman, P.; Ectors, F.; Derivaux, J.; Herlant, M.M. Le dosage radio-immunologique de la progesterone plasmatique chez la vache. C. R. Acad. Hebd. Seances Acad. Sci. D 1975, 280, 335–338. [Google Scholar]
- Perényi, Z.S.; Szenci, O.; Sulon, J.; Drion, P.V.; Beckers, J.-F. Comparison of the ability of three radioimmunoassay to detect pregnancy-associated glycoproteins in bovine plasma. Reprod. Dom. Anim. 2002, 37, 100–104. [Google Scholar] [CrossRef]
- Dal Pozzo, F.; De Clercq, K.; Guyot, H.; Vandemeulebroucke, E.; Sarradin, P.; Vandenbussche, F.; Thiry, E.; Saegerman, C. Experimental reproduction of bluetongue virus serotype 8 clinical disease in calves. Vet. Microbiol. 2009, 136, 352–358. [Google Scholar] [CrossRef]
- Delécolle, J.C. Nouvelle Contribution à l’Etude Systématique et Iconographique des Espèces du Genre Culicoides (Diptera: Ceratopogonidae) du Nord-Est de la France. Ph.D. Thesis, University of Strasbourg, Strasbourg, France, 1985. (In French). [Google Scholar]
- Dyce, A.L. The recognition of nulliparous and parous Culicoides (Diptera: Ceratopogonidae) without dissection. J. Aust. Entomol. Soc. 1969, 8, 11–15. [Google Scholar] [CrossRef]
- Mullens, B.A.; Gerry, A.; Monteys, V.; Pinna, M.; González, A. Field studies on Culicoides (Diptera: Ceratopogonidae) activity and response to deltamethrin applications to sheep in northeastern Spain. J. Med. Entomol. 2010, 47, 106–110. [Google Scholar] [CrossRef]
- Vandenbussche, F.; Vandemeulebroucke, E.; De Clercq, K. Simultaneous detection of bluetongue virus RNA, internal control GAPDH mRNA, and external control synthetic RNA by multiplex real-time PCR. Methods Mol. Biol. 2010, 630, 97–108. [Google Scholar] [CrossRef]
- Vandemeulebroucke, E.; De Clercq, K.; Van der Stede, Y.; Vandenbussche, F. Proposal of a validation method for automated nucleic acid extraction and RT-qPCR analysis: An example with Bluetongue virus. J. Virol. Methods 2010, 165, 76–82. [Google Scholar] [CrossRef]
- Vanbinst, T.; Vandenbussche, F.; Vandemeulebroucke, E.; De Leeuw, I.; Deblauwe, I.; De Deken, G.; Madder, M.; Haubruge, E.; Losson, B.; De Clercq, K. Bluetongue Virus Detection by Real-Time RT-PCR in Culicoides Captured During the 2006 Epizootic in Belgium and Development of an Internal Control. Transbound. Emerg. Dis. 2009, 56, 170–177. [Google Scholar] [CrossRef]
- Vandenbussche, F.; Vanbinst, T.; Verheyden, B.; Van Dessel, W.; Demeestere, L.; Houdart, P.; Bertels, G.; Praet, N.; Berkvens, D.; Mintiens, K.; et al. Evaluation of antibody-ELISA and real-time RT-PCR for the diagnosis and profiling of bluetongue virus serotype 8 during the epidemic in Belgium in 2006. Vet. Microbiol. 2008, 129, 15–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertens, P.P.; Burroughs, J.N.; Walton, A.; Wellby, M.P.; Fu, H.; O’Hara, R.S.; Brookes, S.M.; Mellor, P.S. Enhanced infectivity of modified bluetongue virus particles for two insect cell lines and for two Culicoides vector species. Virology 1996, 217, 582–593. [Google Scholar] [CrossRef] [Green Version]
- Kärber, G. Calculation of the LD50 titer by Kärber method. In Diagnostic Procedures for Viral Rickettsial and Chlamydial Infections, 5th ed.; Lennette, E.H., Schmidt, N.J., Eds.; American Public Health Association: Washington, DC, USA, 1979; pp. 34–35. [Google Scholar]
- Delhalle, L.; De Sadeleer, L.; Bollaerts, K.; Farnir, F.; Saegerman, C.; Korsak, N.; Dewulf, J.; De Zutter, L.; Daube, G. Risk factors for Salmonella and hygiene indicators in the 10 largest Belgian pig slaughterhouses. J. Food Prot. 2008, 71, 1320–1329. [Google Scholar] [CrossRef]
- Petrie, A.; Watson, P. Statistics for Veterinary and Animal Science; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; 391p. [Google Scholar]
- Luedke, A.J.; Walton, T.E. Effect of natural breeding of heifers to a bluetongue virus carrier bull. Bov. Pract. 1981, 16, 96–100. [Google Scholar]
- Schultz, R.D.; Rhodes, P.; Panangala, V.S.; Kaproth, D. Unusual observations on a serologically seronegative bluetongue virus infected bull. Prog. Clin. Biol. Res. 1985, 178, 97–102. [Google Scholar]
- Parsonson, I.M.; Thompson, L.H.; Walton, T.E. Experimentally induced infection with bluetongue virus serotype 11 in cows. Am. J. Vet. Res. 1994, 55, 1529–1534. [Google Scholar]
- MacLachlan, N.J.; Jagels, G.; Rossitto, P.V.; Moore, P.F.; Heidner, H.W. The pathogenesis of experimental bluetongue virus infection of calves. Vet. Pathol. 1990, 27, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Osburn, B.I. The impact of bluetongue virus on reproduction. Comp. Immunol. Microbiol. Infect. Dis. 1994, 17, 189–196. [Google Scholar] [CrossRef]
- Phillips, R.M.; Carnahan, D.L.; Rademacher, D.J. Virus isolation from semen of bulls serologically positive for bluetongue virus. Am. J. Vet. Res. 1986, 47, 84–85. [Google Scholar] [PubMed]
- Gard, G.P.; Melville, L.F.; Shorthose, J.E. Investigations of bluetongue and other arboviruses in the blood and semen of naturally infected bulls. Vet. Microbiol. 1989, 20, 315–322. [Google Scholar] [CrossRef]
- Brodie, S.J.; Wilson, W.C.; O’Hearn, P.M.; Muthui, D.; Diem, K.; Pearson, L.D. The effects of pharmacological and Lentivirus-induced immune suppression on Orbivirus pathogenesis: Assessment of virus burden in blood monocytes and tissues by reverse-transcription-In situ PCR. J. Virol. 1998, 72, 5599–5609. [Google Scholar] [CrossRef] [Green Version]
- Hemati, B.; Contreras, V.; Urien, C.; Bonneau, M.; Takamatsu, H.H.; Mertens, P.P.; Bréard, E.; Sailleau, C.; Zientara, S.; Schwartz-Cornil, L. Bluetongue targets conventional dendritic cells in skin lymph. J. Virol. 2009, 83, 8789–8799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maclachlan, N.J.; Mayo, C.E.; Daniels, P.W.; Savini, G.; Zientara, S.; Gibbs, E.P. Bluetongue. Rev. Sci. Tech. 2015, 34, 329–340. [Google Scholar] [CrossRef]
- Darpel, K.; Batten, C.; Veronesi, E.; Shaw, A.; Anthony, S.; Bachenek-Bankowska, K.; Kgosana, L.; Bin-Tarif, A.; Carpenter, S.; Müller-Doblies, U.; et al. Clinical signs and pathology shown by British sheep and cattle infected with bluetongue virus serotype 8 derived from the 2006 outbreak in northern Europe. Vet. Rec. 2007, 161, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Souza, A.H.; Silva, E.P.; Cunha, A.P.; Gümen, A.; Ayres, H.; Brusveen, D.J.; Guenther, J.N.; Wiltbank, M.C. Ultrasonographic evaluation of endometrial thickness near timed AI as a predictor of fertility in high-producing dairy cows. Theriogenology 2011, 75, 722–733. [Google Scholar] [CrossRef] [PubMed]
- De Smit, A.J.; Bouma, A.; Terpstra, C.; van Oirschot, J.T. Transmission of classical swine fever virus by artificial insemination. Vet. Microbiol. 1999, 67, 239–249. [Google Scholar] [CrossRef]
- World Organisation for Animal Health (OIE). Chapter 4.7: Collection and Processing of Bovine, Small Ruminant and Porcine Semen. 2020. Available online: https://www.oie.int/index.php?id=169&L=0&htmfile=chapitre_coll_semen.htm (accessed on 31 March 2021).
- European Commission, DG Health and Consumers, Animal Health and Welfare. Council Directive 2000/75/EC of 20 November 2000 Laying Down Specific Provisions for the Control and Eradication of Bluetongue. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32000L0075&from=EN (accessed on 31 March 2021).
- European Union. Regulation (EU) 2016/429 of the European Parliament and of the Council of 9 March 2016 on Transmissible Animal Diseases and Amending and Repealing Certain Acts in the Area of Animal Health (‘Animal Health Law’). Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32016R0429&from=NL (accessed on 31 March 2021).
- European Commission, DG Health and Consumers, Animal Health and Welfare. Bluetongue, Overview Seasonally Vector Free Periods 2009–2010. Available online: https://ec.europa.eu/food/sites/food/files/animals/docs/ad_control-measures_bt_overview_seasonally_vfp_2009-2010.pdf (accessed on 24 January 2021).
- Carpenter, S.; Wilson, A.; Barber, J.; Veronesi, E.; Mellor, P.; Venter, G.; Gubbins, S. Temperature dependence of the extrinsic incubation period of orbiviruses in Culicoides biting midges. PLoS ONE 2011, 6, e27987. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Heifer Group | Semen Batch (Bull ID) | Semen RT-qPCR Cp-Value | Heifer ID | Dpai First BTV Genome Detection | Dpai First BTV Isolation Positive | Dpai First BTV Antibody Detection | Pregnancy Loss (Dpai) | |
|---|---|---|---|---|---|---|---|---|
| Exp 1-AI with 0.25 mL semen | A | 1.1 (094) | 20.5 | A1 | 10 | neg | 14 | 35 |
| A2 | 10 | neg | 14 | 26 | ||||
| A3 | 10 | 10 | 14 | 26 | ||||
| B | 1.2 (094) | 25.1 | B1 | neg | nd | neg | No | |
| B2 | neg | nd | neg | No | ||||
| B3 | neg | nd | neg | No | ||||
| C | 2 (202) | 25.4 | C1 | 24 | 28 | 31 | 40 | |
| C2 | neg | nd | neg | No | ||||
| C3 | neg | nd | neg | No | ||||
| D | 3 (572) | 27.2 | D1 | neg | nd | neg | No | |
| D2 | neg | nd | neg | 56 | ||||
| D3 | 42 | neg | 49 | No | ||||
| Ctrl | na | Ctrl-1 | neg | nd | neg | na | ||
| Exp 2-AI with 1 mL semen | E | 2 (202) | 23.4 | E1 | 21 | 24 | 24 | 49 |
| E2 | 21 | 28 | 29 | No | ||||
| E3 | neg | nd | neg | No | ||||
| F | 3 (572) | 25.2 | F1 | neg | nd | neg | No | |
| F2 | neg | nd | neg | No | ||||
| F3 | 7 | 14 | 17 | * | ||||
| Ctrl | na | Ctrl-2 | neg | nd | neg | na |
| Sperm Quality | ||||||
|---|---|---|---|---|---|---|
| Bull ID | Semen Batch | Progressive Motility (%) | Total Motility (%) | Concentration (× 106 Sperm cells/mL) | Viable (%) | Normal Morphology (%) |
| 094 | 1.1 | 27 | 47 | 124 | 44 | 82 |
| 094 | 1.2 | 29 | 56 | 104 | 44 | 80 |
| 202 | 2 | 16 | 33 | 94 | 25 | 70 |
| 572 | 3 | 7 | 26 | 92 | 42 | 63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Clercq, K.; Vandaele, L.; Vanbinst, T.; Riou, M.; Deblauwe, I.; Wesselingh, W.; Pinard, A.; Van Eetvelde, M.; Boulesteix, O.; Leemans, B.; et al. Transmission of Bluetongue Virus Serotype 8 by Artificial Insemination with Frozen–Thawed Semen from Naturally Infected Bulls. Viruses 2021, 13, 652. https://doi.org/10.3390/v13040652
De Clercq K, Vandaele L, Vanbinst T, Riou M, Deblauwe I, Wesselingh W, Pinard A, Van Eetvelde M, Boulesteix O, Leemans B, et al. Transmission of Bluetongue Virus Serotype 8 by Artificial Insemination with Frozen–Thawed Semen from Naturally Infected Bulls. Viruses. 2021; 13(4):652. https://doi.org/10.3390/v13040652
Chicago/Turabian StyleDe Clercq, Kris, Leen Vandaele, Tine Vanbinst, Mickaël Riou, Isra Deblauwe, Wendy Wesselingh, Anne Pinard, Mieke Van Eetvelde, Olivier Boulesteix, Bart Leemans, and et al. 2021. "Transmission of Bluetongue Virus Serotype 8 by Artificial Insemination with Frozen–Thawed Semen from Naturally Infected Bulls" Viruses 13, no. 4: 652. https://doi.org/10.3390/v13040652
APA StyleDe Clercq, K., Vandaele, L., Vanbinst, T., Riou, M., Deblauwe, I., Wesselingh, W., Pinard, A., Van Eetvelde, M., Boulesteix, O., Leemans, B., Gélineau, R., Vercauteren, G., Van der Heyden, S., Beckers, J. -F., Saegerman, C., Sammin, D., de Kruif, A., & De Leeuw, I. (2021). Transmission of Bluetongue Virus Serotype 8 by Artificial Insemination with Frozen–Thawed Semen from Naturally Infected Bulls. Viruses, 13(4), 652. https://doi.org/10.3390/v13040652