
The VP3 Protein of Bluetongue Virus Associates with the MAVS Complex and Interferes with the RIG-I-Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. DNA Transfection and Plasmids
2.3. Reporter Luciferase Assay
2.4. Gaussia Princeps Complementation Assay (GPCA)
2.5. Co-Affinity Purification
2.6. Western Blot Analysis
2.7. Fluorescence Microscopy
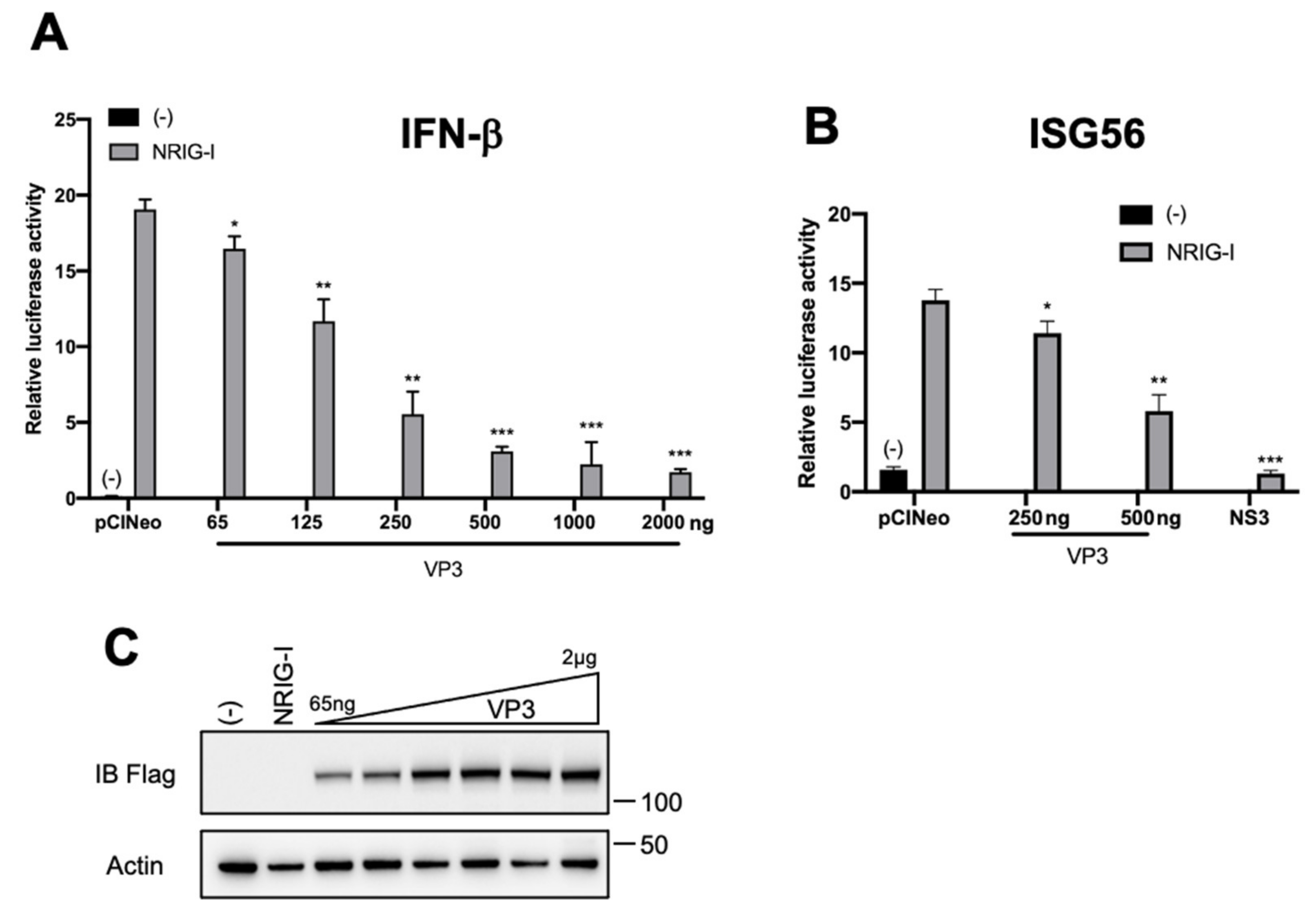
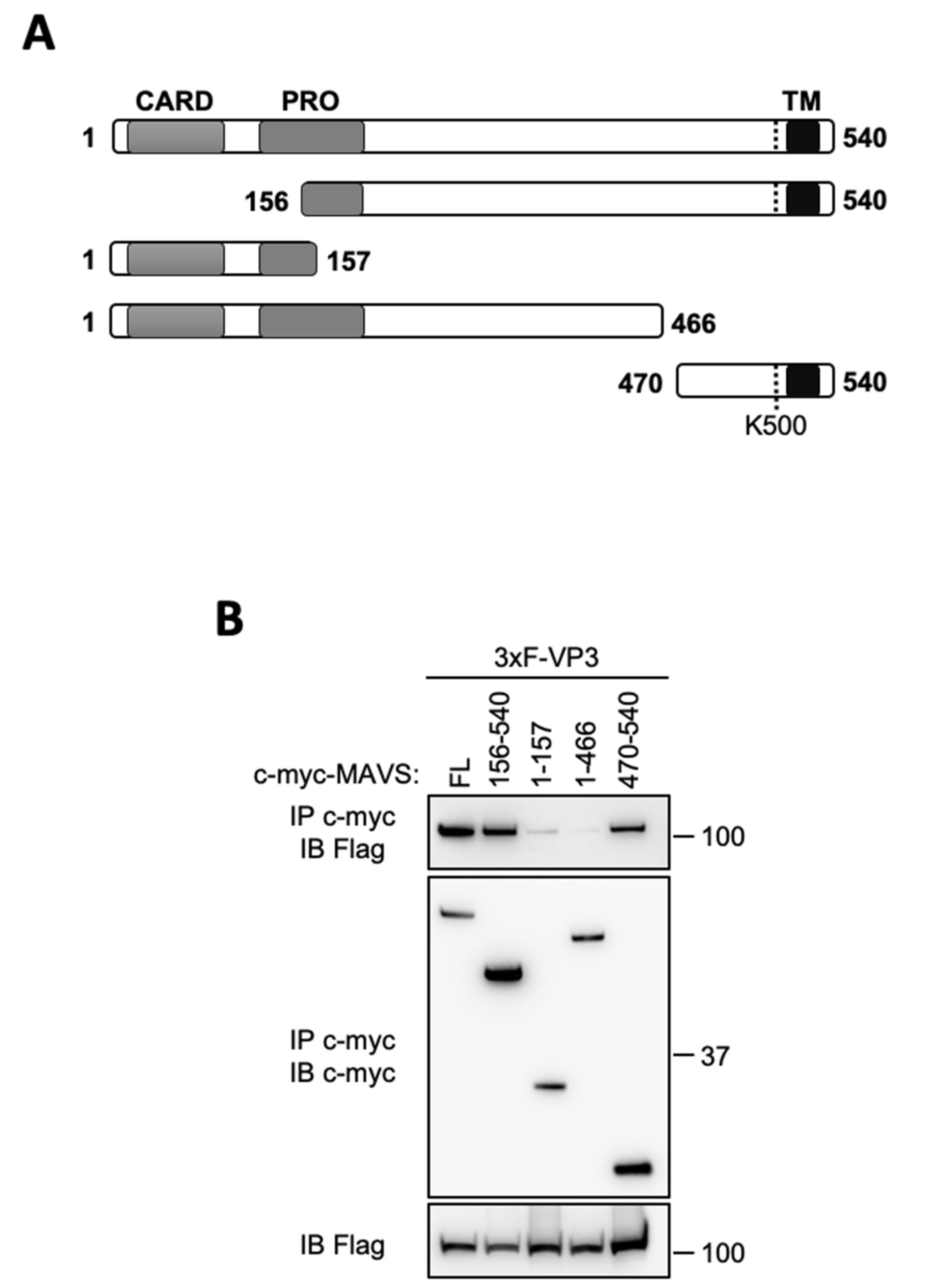
3. Results
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Belhouchet, M.; Mohd Jaafar, F.; Firth, A.E.; Grimes, J.M.; Mertens, P.P.C.; Attoui, H. Detection of a Fourth Orbivirus Non-Structural Protein. PLoS ONE 2011, 6, e25697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratinier, M.; Caporale, M.; Golder, M.; Franzoni, G.; Allan, K.; Nunes, S.F.; Armezzani, A.; Bayoumy, A.; Rixon, F.; Shaw, A.; et al. Identification and Characterization of a Novel Non-Structural Protein of Bluetongue Virus. PLoS Pathog. 2011, 7, e1002477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, M.; Hardy, A.; Barry, G.; Pinto, R.M.; Caporale, M.; Melzi, E.; Hughes, J.; Taggart, A.; Janowicz, A.; Varela, M.; et al. Characterization of a Second Open Reading Frame in Genome Segment 10 of Bluetongue Virus. J. Gen. Virol. 2015, 96, 3280–3293. [Google Scholar] [CrossRef] [PubMed]
- Roy, P. Bluetongue Virus Proteins and Particles and Their Role in Virus Entry, Assembly, and Release. Adv. Virus Res. 2005, 64, 69–123. [Google Scholar] [CrossRef]
- Schwartz-Cornil, I.; Mertens, P.P.; Contreras, V.; Hemati, B.; Pascale, F.; Breard, E.; Mellor, P.S.; MacLachlan, N.J.; Zientara, S. Bluetongue Virus: Virology, Pathogenesis and Immunity. Vet. Res. 2008, 39, 46. [Google Scholar] [CrossRef] [Green Version]
- Saegerman, C.; Mellor, P.; Uyttenhoef, A.; Hanon, J.B.; Kirschvink, N.; Haubruge, E.; Delcroix, P.; Houtain, J.Y.; Pourquier, P.; Vandenbussche, F.; et al. The Most Likely Time and Place of Introduction of BTV8 into Belgian Ruminants. PLoS ONE 2010, 5, e9405. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and Innate Immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Takahashi, K.; Sato, S.; Coban, C.; Kumar, H.; Kato, H.; Ishii, K.J.; Takeuchi, O.; Akira, S. IPS-1, an Adaptor Triggering RIG-I- and Mda5-Mediated Type I Interferon Induction. Nat. Immunol. 2005, 6, 981–988. [Google Scholar] [CrossRef]
- Seth, R.B.; Sun, L.; Ea, C.-K.; Chen, Z.J. Identification and Characterization of MAVS, a Mitochondrial Antiviral Signaling Protein That Activates NF-KappaB and IRF 3. Cell 2005, 122, 669–682. [Google Scholar] [CrossRef] [Green Version]
- Meylan, E.; Curran, J.; Hofmann, K.; Moradpour, D.; Binder, M.; Bartenschlager, R.; Tschopp, J. Cardif Is an Adaptor Protein in the RIG-I Antiviral Pathway and Is Targeted by Hepatitis C Virus. Nature 2005, 437, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.-G.; Wang, Y.-Y.; Han, K.-J.; Li, L.-Y.; Zhai, Z.; Shu, H.-B. VISA Is an Adapter Protein Required for Virus-Triggered IFN-Beta Signaling. Mol. Cell 2005, 19, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.A.; McWhirter, S.M.; Faia, K.L.; Rowe, D.C.; Latz, E.; Golenbock, D.T.; Coyle, A.J.; Liao, S.-M.; Maniatis, T. IKKepsilon and TBK1 Are Essential Components of the IRF3 Signaling Pathway. Nat. Immunol. 2003, 4, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Hemmi, H.; Takeuchi, O.; Sato, S.; Yamamoto, M.; Kaisho, T.; Sanjo, H.; Kawai, T.; Hoshino, K.; Takeda, K.; Akira, S. The Roles of Two IkappaB Kinase-Related Kinases in Lipopolysaccharide and Double Stranded RNA Signaling and Viral Infection. J. Exp. Med. 2004, 199, 1641–1650. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; tenOever, B.R.; Grandvaux, N.; Zhou, G.-P.; Lin, R.; Hiscott, J. Triggering the Interferon Antiviral Response through an IKK-Related Pathway. Science 2003, 300, 1148–1151. [Google Scholar] [CrossRef]
- Randall, R.E.; Goodbourn, S. Interferons and Viruses: An Interplay between Induction, Signalling, Antiviral Responses and Virus Countermeasures. J. Gen. Virol. 2008, 89, 1–47. [Google Scholar] [CrossRef]
- Chauveau, E.; Doceul, V.; Lara, E.; Adam, M.; Breard, E.; Sailleau, C.; Viarouge, C.; Desprat, A.; Meyer, G.; Schwartz-Cornil, I.; et al. Sensing and Control of Bluetongue Virus Infection in Epithelial Cells via RIG-I and MDA5 Helicases. J. Virol. 2012, 86, 11789–11799. [Google Scholar] [CrossRef] [Green Version]
- Ruscanu, S.; Pascale, F.; Bourge, M.; Hemati, B.; Elhmouzi-Younes, J.; Urien, C.; Bonneau, M.; Takamatsu, H.; Hope, J.; Mertens, P.; et al. The Double-Stranded RNA Bluetongue Virus Induces Type I Interferon in Plasmacytoid Dendritic Cells via a MYD88-Dependent TLR7/8-Independent Signaling Pathway. J. Virol. 2012, 86, 5817–5828. [Google Scholar] [CrossRef] [Green Version]
- Doceul, V.; Chauveau, E.; Lara, E.; Bréard, E.; Sailleau, C.; Zientara, S.; Vitour, D. Dual Modulation of Type I Interferon Response by Bluetongue Virus. J. Virol. 2014, 88, 10792–10802. [Google Scholar] [CrossRef] [Green Version]
- Chauveau, E.; Doceul, V.; Lara, E.; Breard, E.; Sailleau, C.; Vidalain, P.-O.; Meurs, E.F.; Dabo, S.; Schwartz-Cornil, I.; Zientara, S.; et al. NS3 of Bluetongue Virus Interferes with the Induction of Type I Interferon. J. Virol. 2013, 87, 8241–8246. [Google Scholar] [CrossRef] [Green Version]
- Pourcelot, M.; Zemirli, N.; Silva Da Costa, L.; Loyant, R.; Garcin, D.; Vitour, D.; Munitic, I.; Vazquez, A.; Arnoult, D. The Golgi Apparatus Acts as a Platform for TBK1 Activation after Viral RNA Sensing. BMC Biol. 2016, 14, 69. [Google Scholar] [CrossRef] [PubMed]
- Avia, M.; Rojas, J.M.; Miorin, L.; Pascual, E.; Van Rijn, P.A.; Martín, V.; García-Sastre, A.; Sevilla, N. Virus-Induced Autophagic Degradation of STAT2 as a Mechanism for Interferon Signaling Blockade. EMBO Rep. 2019, 20, e48766. [Google Scholar] [CrossRef] [PubMed]
- Kar, A.K.; Iwatani, N.; Roy, P. Assembly and Intracellular Localization of the Bluetongue Virus Core Protein VP3. J. Virol. 2005, 79, 11487–11495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundlacz, C.; Pourcelot, M.; Fablet, A.; Amaral Da Silva Moraes, R.; Léger, T.; Morlet, B.; Viarouge, C.; Sailleau, C.; Turpaud, M.; Gorlier, A.; et al. Novel Function of Bluetongue Virus NS3 Protein in Regulation of the MAPK/ERK Signaling Pathway. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Vitour, D.; Dabo, S.; Ahmadi Pour, M.; Vilasco, M.; Vidalain, P.O.; Jacob, Y.; Mezel-Lemoine, M.; Paz, S.; Arguello, M.; Lin, R.; et al. Polo-like Kinase 1 (PLK1) Regulates Interferon (IFN) Induction by MAVS. J. Biol. Chem. 2009, 284, 21797–21809. [Google Scholar] [CrossRef] [Green Version]
- Poirson, J.; Biquand, E.; Straub, M.-L.; Cassonnet, P.; Nominé, Y.; Jones, L.; van der Werf, S.; Travé, G.; Zanier, K.; Jacob, Y.; et al. Mapping the Interactome of HPV E6 and E7 Oncoproteins with the Ubiquitin-Proteasome System. FEBS J. 2017, 284, 3171–3201. [Google Scholar] [CrossRef] [Green Version]
- Cassonnet, P.; Rolloy, C.; Neveu, G.; Vidalain, P.O.; Chantier, T.; Pellet, J.; Jones, L.; Muller, M.; Demeret, C.; Gaud, G.; et al. Benchmarking a Luciferase Complementation Assay for Detecting Protein Complexes. Nat. Methods 2011, 8, 990–992. [Google Scholar] [CrossRef]
- Versteeg, G.A.; Garcia-Sastre, A. Viral Tricks to Grid-Lock the Type I Interferon System. Curr. Opin. Microbiol. 2010, 13, 508–516. [Google Scholar] [CrossRef]
- Paz, S.; Vilasco, M.; Werden, S.J.; Arguello, M.; Joseph-Pillai, D.; Zhao, T.; Nguyen, T.L.; Sun, Q.; Meurs, E.F.; Lin, R.; et al. A Functional C-Terminal TRAF3-Binding Site in MAVS Participates in Positive and Negative Regulation of the IFN Antiviral Response. Cell Res. 2011, 21, 895–910. [Google Scholar] [CrossRef]
- Konno, H.; Yamamoto, T.; Yamazaki, K.; Gohda, J.; Akiyama, T.; Semba, K.; Goto, H.; Kato, A.; Yujiri, T.; Imai, T.; et al. TRAF6 Establishes Innate Immune Responses by Activating NF-KappaB and IRF7 upon Sensing Cytosolic Viral RNA and DNA. PLoS ONE 2009, 4, e5674. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Zhang, Z.; Zhang, Z.; Wang, Y.; Li, C.; Wang, X.; He, F.; Sun, L.; Jiao, S.; Shi, W.; et al. Structural Insights into Mitochondrial Antiviral Signaling Protein (MAVS)-Tumor Necrosis Factor Receptor-Associated Factor 6 (TRAF6) Signaling. J. Biol. Chem. 2015, 290, 26811–26820. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pourcelot, M.; Amaral Moraes, R.; Fablet, A.; Bréard, E.; Sailleau, C.; Viarouge, C.; Postic, L.; Zientara, S.; Caignard, G.; Vitour, D. The VP3 Protein of Bluetongue Virus Associates with the MAVS Complex and Interferes with the RIG-I-Signaling Pathway. Viruses 2021, 13, 230. https://doi.org/10.3390/v13020230
Pourcelot M, Amaral Moraes R, Fablet A, Bréard E, Sailleau C, Viarouge C, Postic L, Zientara S, Caignard G, Vitour D. The VP3 Protein of Bluetongue Virus Associates with the MAVS Complex and Interferes with the RIG-I-Signaling Pathway. Viruses. 2021; 13(2):230. https://doi.org/10.3390/v13020230
Chicago/Turabian StylePourcelot, Marie, Rayane Amaral Moraes, Aurore Fablet, Emmanuel Bréard, Corinne Sailleau, Cyril Viarouge, Lydie Postic, Stéphan Zientara, Grégory Caignard, and Damien Vitour. 2021. "The VP3 Protein of Bluetongue Virus Associates with the MAVS Complex and Interferes with the RIG-I-Signaling Pathway" Viruses 13, no. 2: 230. https://doi.org/10.3390/v13020230
APA StylePourcelot, M., Amaral Moraes, R., Fablet, A., Bréard, E., Sailleau, C., Viarouge, C., Postic, L., Zientara, S., Caignard, G., & Vitour, D. (2021). The VP3 Protein of Bluetongue Virus Associates with the MAVS Complex and Interferes with the RIG-I-Signaling Pathway. Viruses, 13(2), 230. https://doi.org/10.3390/v13020230