
Genome-Wide Analysis of the RAV Transcription Factor Genes in Rice Reveals Their Response Patterns to Hormones and Virus Infection

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Searches for RAV Genes in the Rice, Arabidopsis, and Maize Genome Databases
2.2. Multiple Sequence Alignment and Phylogenetic Analysis
2.3. Gene Structure and Conserved Motif Analysis
2.4. Chromosomal Distribution of RAV Genes
2.5. Plant Materials and Growth Conditions
2.6. Hormone Treatments
2.7. RSV and SRBSDV Infection
2.8. RNA Extraction and qRT-PCR
2.9. Statistical Analysis
3. Results
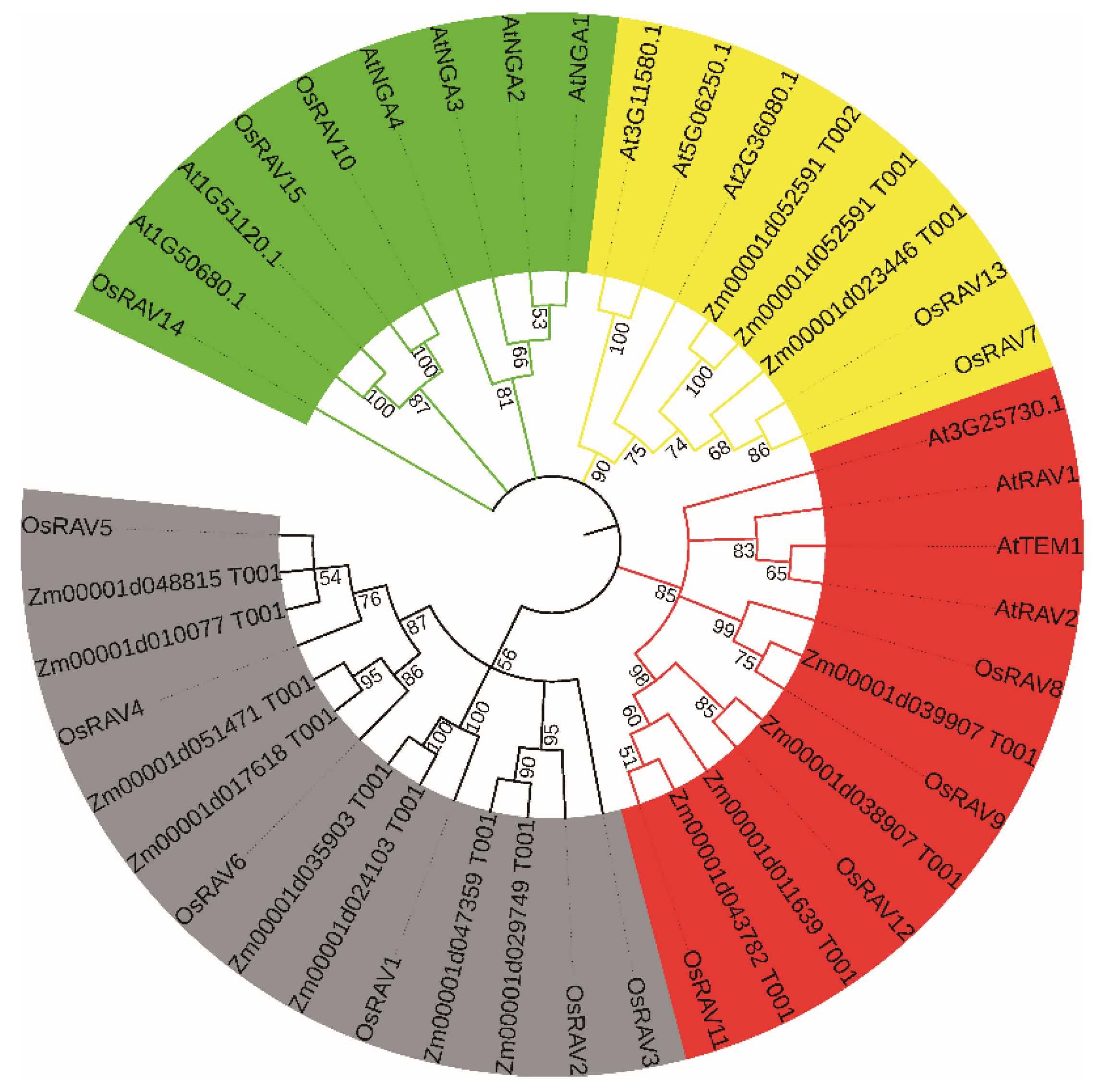
3.1. Identification and Phylogenetic Analysis of RAV Transcription Factors in Rice
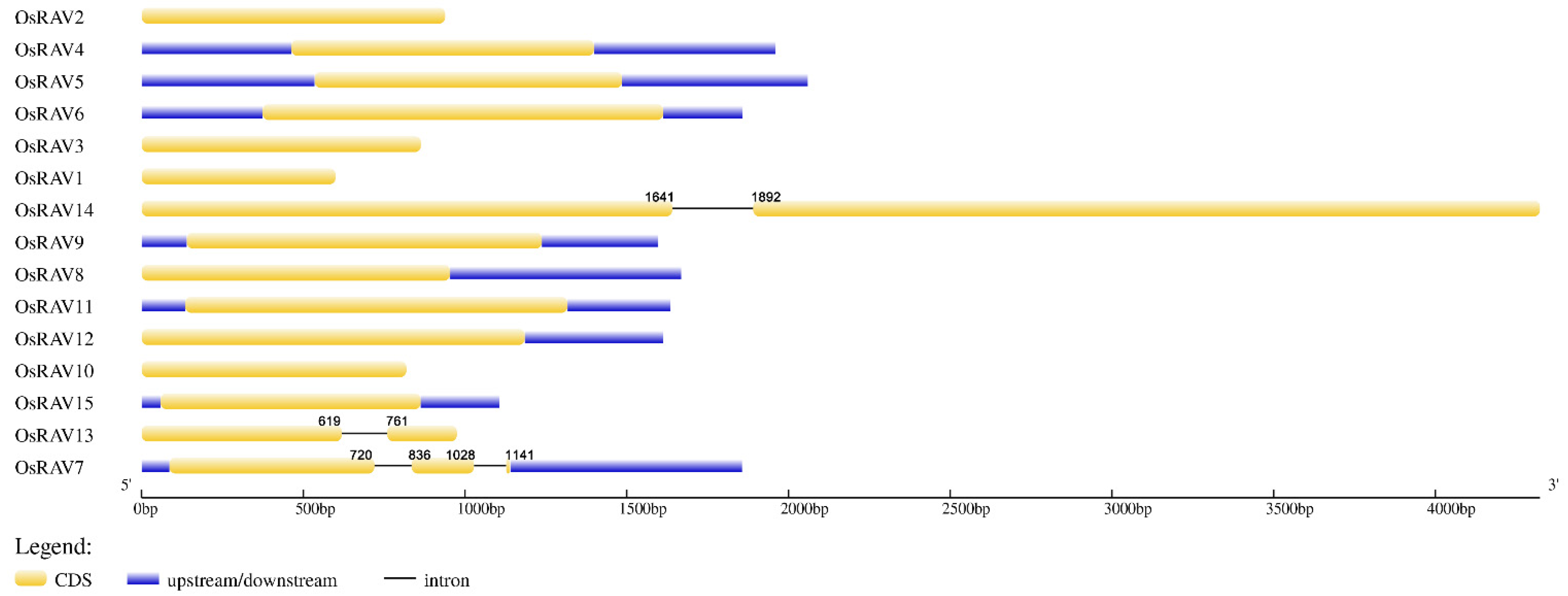
3.2. Gene Structure and Domain Analysis of Rice RAV Genes
3.3. Chromosomal Distribution Analysis of Rice RAV Genes
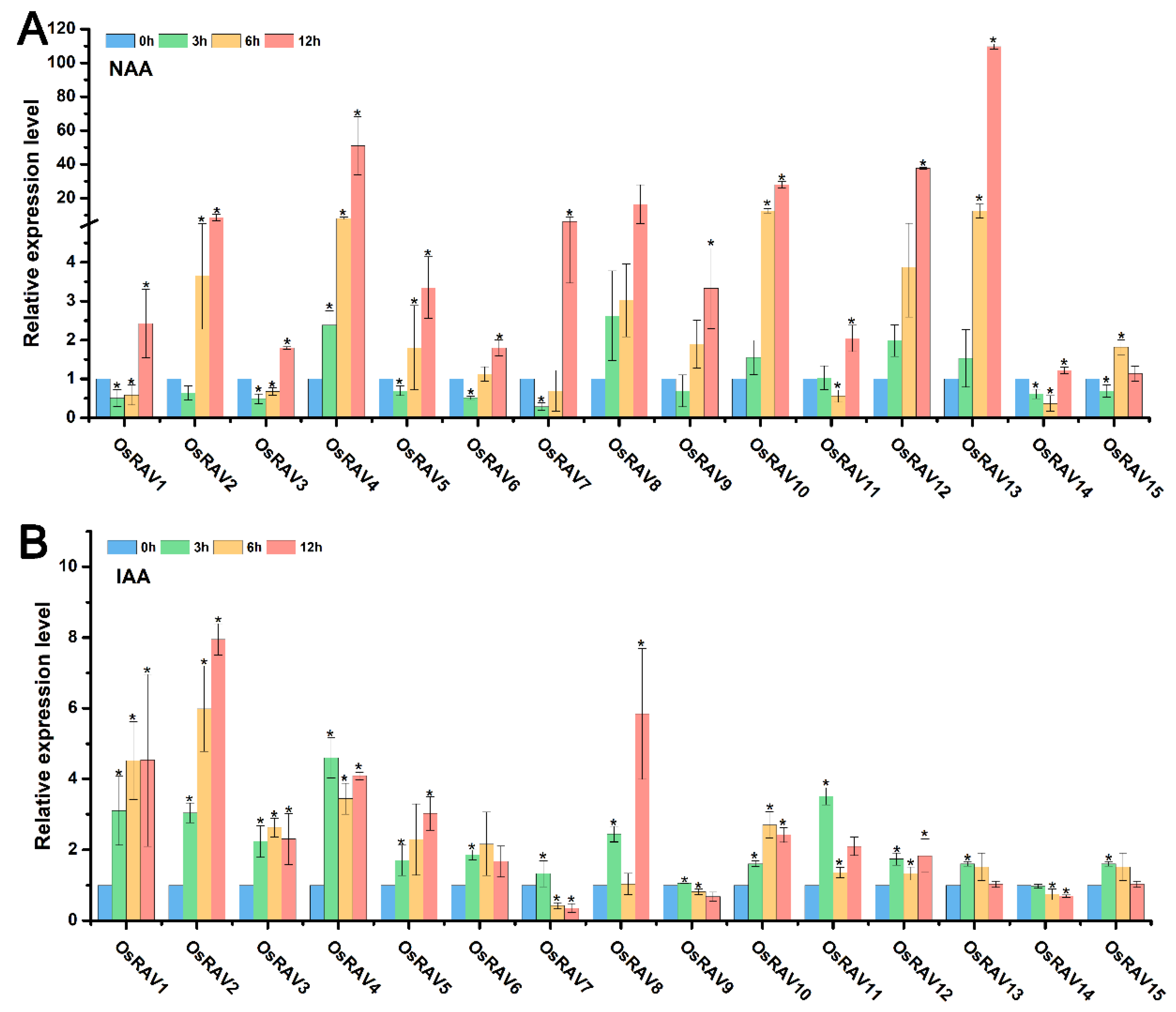
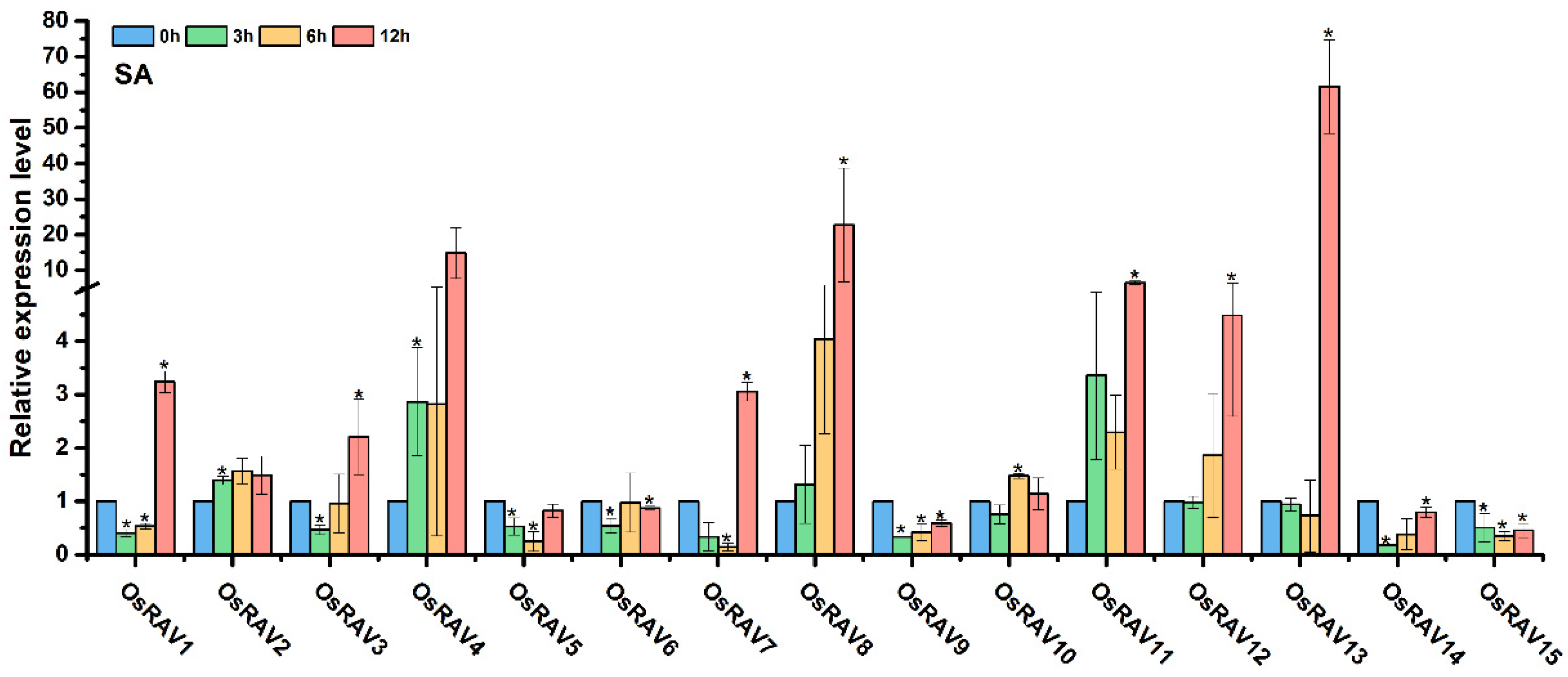
3.4. Expression Patterns of Rice RAV Genes under Hormone Treatments
3.5. Expression Profiling of Rice RAV Genes in Response to Rice Stripe Virus (RSV) and Southern Rice Black-Streaked Dwarf Virus (SRBSDV) Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| qRT-PCR | Quantitative real-time PCR |
| RSV | Rice stripe virus |
| SRBSDV | Southern rice black-streaked dwarf virus |
| TFs | Transcription factors |
| DBD | DNA binding domain |
| RAV | Related to ABI3/VP1 |
| ARF | AUXIN RESPONSE FACTOR |
| LEC2 | LEAFY COTYLEDON2 |
| ABI3 | ABSCISIC ACID INSENSITIVE3 |
| REM | Reproductive meristem |
| VP1 | VIVIPAROUS1 |
| FT | Flowering locus T |
| ML | Maximum-likelihood |
| NJ | Neighbor-joining |
| BL | Epibrassinolide |
| MeJA | Methyl jasmonate |
| NAA | 1-naphthylacetic acid |
| IAA | Indole-3-acetic acid |
| SA | Salicylic acid |
| Os | Oryza sativa |
| UTRs | Untranslated regions |
| SBPH | The small brown planthopper |
| WBPH | The white-backed planthopper |
| dpi | Days post inoculation |
References
- Ptashne, M. How eukaryotic transcriptional activators work. Nature 1988, 335, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.-Z.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis Transcription Factors: Genome-Wide Comparative Analysis Among Eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Liu, T.; Tian, C.; Sun, S.; Li, J.; Chen, M. Transcription Factors in Rice: A Genome-wide Comparative Analysis between Monocots and Eudicots. Plant Mol. Biol. 2005, 59, 191–203. [Google Scholar] [CrossRef]
- Swaminathan, K.; Peterson, K.; Jack, T. The plant B3 superfamily. Trends Plant Sci. 2008, 13, 647–655. [Google Scholar] [CrossRef]
- Matías-Hernández, L.; Aguilar-Jaramillo, A.E.; Marín-González, E.; Suárez-López, P.; Pelaz, S. RAV genes: Regulation of floral induction and beyond. Ann. Bot. 2014, 114, 1459–1470. [Google Scholar] [CrossRef] [PubMed]
- Kagaya, Y.; Ohmiya, K.; Hattori, T. RAV1, a novel DNA-binding protein, binds to bipartite recognition sequence through two distinct DNA-binding domains uniquely found in higher plants. Nucleic Acids Res. 1999, 27, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Jofuku, K.D.; den Boer, B.G.; Van Montagu, M.; Okamuro, J.K. Control of Arabidopsis flower and seed development by the homeotic gene APETALA2. Plant Cell 1994, 6, 1211–1225. [Google Scholar]
- Hu, Y.X.; Wang, Y.H.; Liu, X.F.; Li, J.Y. Arabidopsis RAV1 is down-regulated by brassinosteroid and may act as a negative regulator during plant development. Cell Res. 2004, 14, 8–15. [Google Scholar] [CrossRef]
- Zhao, L.; Luo, Q.; Yang, C.; Han, Y.; Li, W. A RAV-like transcription factor controls photosynthesis and senescence in soybean. Planta 2008, 227, 1389–1399. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.P.; Pekker, I.; Goldshmidt, A.; Blum, E.; Amsellem, Z.; Eshed, Y. Endogenous and Synthetic MicroRNAs Stimulate Simultaneous, Efficient, and Localized Regulation of Multiple Targets in Diverse Species. Plant Cell 2006, 18, 1134–1151. [Google Scholar] [CrossRef] [Green Version]
- Castillejo, C.; Pelaz, S. The Balance between CONSTANS and TEMPRANILLO Activities Determines FT Expression to Trigger Flowering. Curr. Biol. 2008, 18, 1338–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osnato, M.; Matias-Hernandez, L.; Aguilar-Jaramillo, A.E.; Kater, M.M.; Pelaz, S. Genes of the RAV Family Control Heading Date and Carpel Development in Rice. Plant Physiol. 2020, 183, 1663–1680. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Chang, Y.; Zeng, H.; Liu, G.; He, C.; Shi, H. RAV transcription factors are essential for disease resistance against cassava bacterial blight via activation of melatonin biosynthesis genes. J. Pineal Res. 2017, 64, e12454. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.K.; Lee, S.C.; Hwang, B.K. Activation of pepper basic PR-1 gene promoter during defense signaling to pathogen, abiotic and environmental stresses. Gene 2005, 356, 169–180. [Google Scholar] [CrossRef]
- Sohn, K.H.; Lee, S.C.; Jung, H.W.; Hong, J.K.; Hwang, B.K. Expression and functional roles of the pepper pathogen-induced transcription factor RAV1 in bacterial disease resistance, and drought and salt stress tolerance. Plant Mol. Biol. 2006, 61, 897. [Google Scholar] [CrossRef]
- Alonso, J.M.; Stepanova, A.N.; Leisse, T.J.; Kim, C.J.; Chen, H.; Shinn, P.; Stevenson, D.K.; Zimmerman, J.; Barajas, P.; Cheuk, R.; et al. Genome-Wide Insertional Mutagenesis of Arabidopsis thaliana. Science 2003, 301, 653–657. [Google Scholar] [CrossRef] [Green Version]
- Fu, M.; Kang, H.K.; Son, S.-H.; Kim, S.-K.; Nam, K.H. A Subset of Arabidopsis RAV Transcription Factors Modulates Drought and Salt Stress Responses Independent of ABA. Plant Cell Physiol. 2014, 55, 1892–1904. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.-J.; Liu, Q.-Q.; Wilson, J.D.; Gu, M.-H.; Shi, Y.-C. Digestibility and physicochemical properties of rice (Oryza sativa L.) flours and starches differing in amylose content. Carbohydr. Polym. 2011, 86, 1751–1759. [Google Scholar] [CrossRef]
- Phytozome, the Plant Comparative Genomics Portal of the Department of Energy’s Joint Genome Institute. Available online: https://phytozome.jgi.doe.gov/pz/portal.html (accessed on 15 April 2021).
- Du, H.; Huang, M.; Zhang, Z.; Cheng, S. Genome-wide analysis of the AP2/ERF gene family in maize waterlogging stress response. Euphytica 2014, 198, 115–126. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; I Hurwitz, D.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batch Web CD-Search Tool. Available online: https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi (accessed on 15 April 2021).
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.-C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein Identification and Analysis Tools in the ExPASy Server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar]
- Bioinformatics Resource Portal of the SIB Swiss Institute of Bioinformatics Home. Available online: http://www.expasy.org/ (accessed on 15 April 2021).
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clustal: Multiple Sequence Alignment Home Page. Available online: http://www.clustal.org/ (accessed on 15 April 2021).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Molecular Evolutionary Genetics Analysis Home Page. Available online: http://www.megasoftware.net/ (accessed on 15 April 2021).
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- GSDS 2.0: Gene Structure Display Server Home Page. Available online: http://gsds.gao-lab.org/ (accessed on 15 April 2021).
- Ouyang, S.; Zhu, W.; Hamilton, J.; Lin, H.; Campbell, M.; Childs, K.; Thibaud-Nissen, F.; Malek, R.L.; Lee, Y.; Zheng, L.; et al. The TIGR Rice Genome Annotation Resource: Improvements and new features. Nucleic Acids Res. 2006, 35, D883–D887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Li, L.; He, Y.; Qin, Q.; Chen, C.; Wei, Z.; Tan, X.; Xie, K.; Zhang, R.; Hong, G.; et al. Distinct modes of manipulation of rice auxin response factor OsARF17 by different plant RNA viruses for infection. Proc. Natl. Acad. Sci. USA 2020, 117, 9112–9121. [Google Scholar] [CrossRef]
- Li, L.; Zhang, H.; Chen, C.; Huang, H.; Tan, X.; Wei, Z.; Li, J.; Yan, F.; Zhang, C.; Chen, J.; et al. A class of independently evolved transcriptional repressors in plant RNA viruses facilitates viral infection and vector feeding. Proc. Natl. Acad. Sci. USA 2021, 118, 2016673118. [Google Scholar] [CrossRef]
- Sun, Z.; He, Y.; Li, J.; Wang, X.; Chen, J. Genome-Wide Characterization of Rice Black Streaked Dwarf Virus-Responsive MicroRNAs in Rice Leaves and Roots by Small RNA and Degradome Sequencing. Plant Cell Physiol. 2014, 56, 688–699. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Seifert, E. OriginPro 9.1: Scientific Data Analysis and Graphing Software—Software Review. J. Chem. Inf. Model. 2014, 54, 1552. [Google Scholar] [CrossRef]
- Toriyama, S.; Takahashi, M.; Sano, Y.; Shimizu, T.; Ishihama, A. Nucleotide sequence of RNA 1, the largest genomic segment of rice stripe virus, the prototype of the tenuiviruses. J. Gen. Virol. 1994, 75, 3569–3579. [Google Scholar] [CrossRef]
- Du, Z.; Xiao, D.; Wu, J.; Jia, D.; Yuan, Z.; Liu, Y.; Hu, L.; Han, Z.; Wei, T.; Lin, Q.; et al. p2 of Rice stripe virus (RSV) interacts with OsSGS3 and is a silencing suppressor. Mol. Plant Pathol. 2011, 12, 808–814. [Google Scholar] [CrossRef]
- Wei, T.-Y.; Yang, J.-G.; Liao, F.-L.; Gao, F.-L.; Lu, L.-M.; Zhang, X.-T.; Li, F.; Wu, Z.-J.; Lin, Q.-Y.; Xie, L.-H.; et al. Genetic diversity and population structure of rice stripe virus in China. J. Gen. Virol. 2009, 90, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Li, W.; Wei, T.; Wu, Z.; Xie, L. Advances in the studies of Rice stripe virus. Front. Agric. China 2010, 4, 287–292. [Google Scholar] [CrossRef]
- Wei, T.; Li, Y. Rice Reoviruses in Insect Vectors. Annu. Rev. Phytopathol. 2016, 54, 99–120. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.-P.; Xu, Z.-S.; Zheng, W.-J.; Zhao, W.; Wang, Y.-X.; Yu, T.-F.; Chen, M.; Zhou, Y.-B.; Min, D.-H.; Ma, Y.-Z.; et al. Genome-Wide Analysis of the RAV Family in Soybean and Functional Identification of GmRAV-03 Involvement in Salt and Drought Stresses and Exogenous ABA Treatment. Front. Plant Sci. 2017, 8, 905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Zhu, W.; Silva, J.C.; Gu, X.; Buell, C.R. Intron gain and loss in segmentally duplicated genes in rice. Genome Biol. 2006, 7, R41. [Google Scholar] [CrossRef] [Green Version]
- Rose, A.B. Intron-Mediated Regulation of Gene Expression. In Nuclear Pre-mRNA Processing in Plants; Reddy, A.S.N., Golovkin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 277–290. ISBN 978-3-540-76775-6. [Google Scholar]
- Brenchley, R.; Spannagl, M.; Pfeifer, M.; Barker, G.L.A.; D’Amore, R.; Allen, A.M.; McKenzie, N.; Kramer, M.; Kerhornou, A.; Bolser, D.; et al. Analysis of the bread wheat genome using whole-genome shotgun sequencing. Nat. Cell Biol. 2012, 491, 705–710. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Gene Name | Gene ID Number | Amino Acid Residues | B3 Domain | AP2 Domain | pI |
|---|---|---|---|---|---|---|
| 1 | OsRAV1 | >LOC_Os06g01860 | 199 | 37–126 | 9.56 | |
| 2 | OsRAV2 | >LOC_Os10g39190 | 312 | 35–153 | 9.62 | |
| 3 | OsRAV3 | >LOC_Os08g06120 | 287 | 71–159 | 5.03 | |
| 4 | OsRAV4 | >LOC_Os03g02900 | 311 | 37–141 | 9.80 | |
| 5 | OsRAV5 | >LOC_Os04g49230 | 316 | 110–195 | 8.81 | |
| 6 | OsRAV6 | >LOC_Os02g45850 | 412 | 96–201 | 7.33 | |
| 7 | OsRAV7 | >LOC_Os11g05740 | 279 | 38–144 | 6.67 | |
| 8 | OsRAV8 | >LOC_Os01g04750 | 317 | 176–271 | 65–119 | 7.08 |
| 9 | OsRAV9 | >LOC_Os01g04800 | 365 | 182–294 | 68–123 | 9.82 |
| 10 | OsRAV10 | >LOC_Os02g25830 | 272 | 28–132 | 6.85 | |
| 11 | OsRAV11 | >LOC_Os01g49830 | 393 | 216–312 | 81–138 | 9.09 |
| 12 | OsRAV12 | >LOC_Os05g47650 | 394 | 193–308 | 67–123 | 9.51 |
| 13 | OsRAV13 | >LOC_Os12g06080 | 277 | 36–141 | 7.25 | |
| 14 | OsRAV14 | >LOC_Os07g17230 | 1357 | 74–172 | 5.76 | |
| 15 | OsRAV15 | >LOC_Os02g25820 | 267 | 28–128 | 9.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Li, Y.; Zhang, H.; Ma, Q.; Wei, Z.; Chen, J.; Sun, Z. Genome-Wide Analysis of the RAV Transcription Factor Genes in Rice Reveals Their Response Patterns to Hormones and Virus Infection. Viruses 2021, 13, 752. https://doi.org/10.3390/v13050752
Chen C, Li Y, Zhang H, Ma Q, Wei Z, Chen J, Sun Z. Genome-Wide Analysis of the RAV Transcription Factor Genes in Rice Reveals Their Response Patterns to Hormones and Virus Infection. Viruses. 2021; 13(5):752. https://doi.org/10.3390/v13050752
Chicago/Turabian StyleChen, Changhai, Yanjun Li, Hehong Zhang, Qiang Ma, Zhongyan Wei, Jianping Chen, and Zongtao Sun. 2021. "Genome-Wide Analysis of the RAV Transcription Factor Genes in Rice Reveals Their Response Patterns to Hormones and Virus Infection" Viruses 13, no. 5: 752. https://doi.org/10.3390/v13050752
APA StyleChen, C., Li, Y., Zhang, H., Ma, Q., Wei, Z., Chen, J., & Sun, Z. (2021). Genome-Wide Analysis of the RAV Transcription Factor Genes in Rice Reveals Their Response Patterns to Hormones and Virus Infection. Viruses, 13(5), 752. https://doi.org/10.3390/v13050752