
Structural Insights into the Human Astrovirus Capsid

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
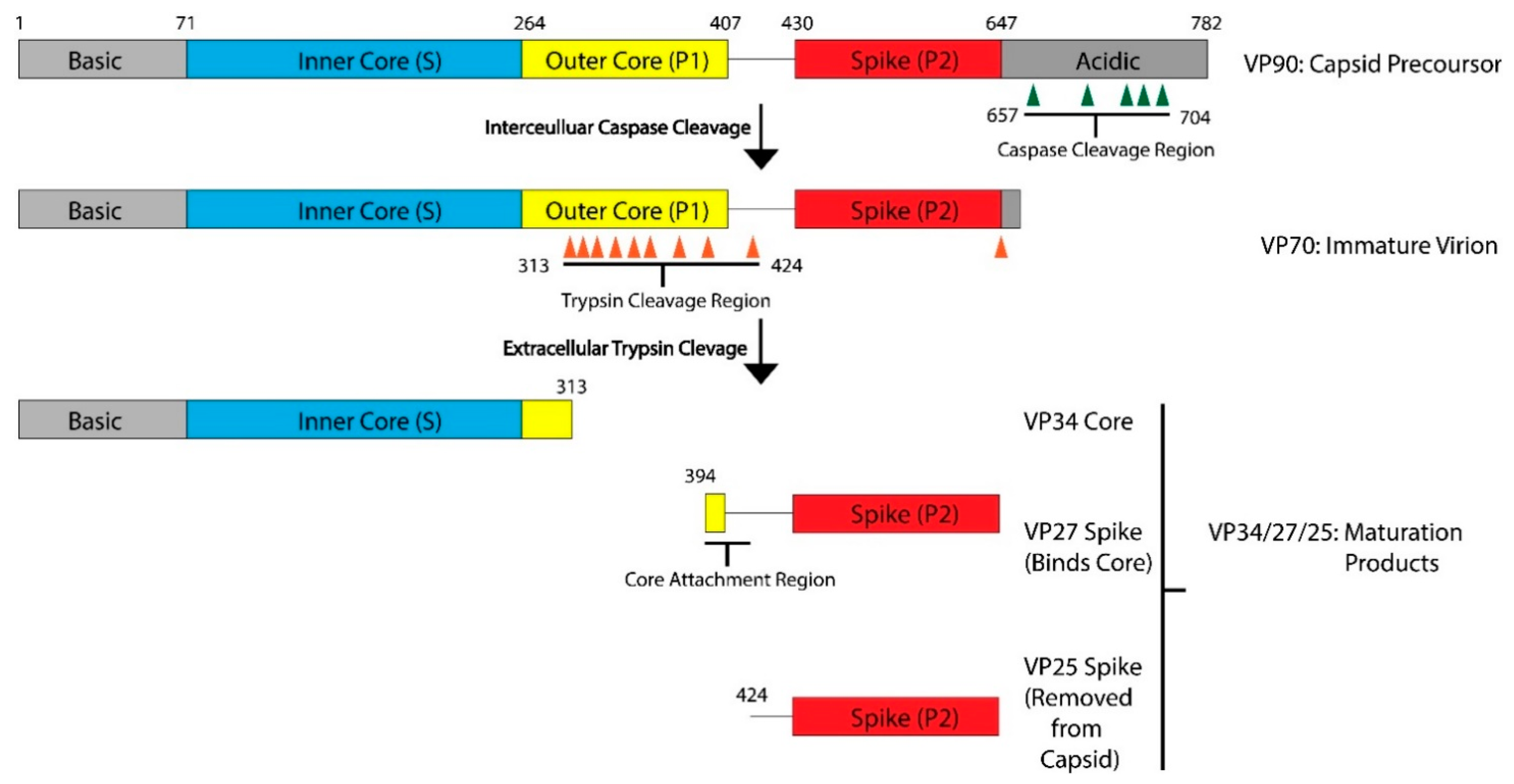
2. Astrovirus Capsid Maturation Is a Host-Driven Process
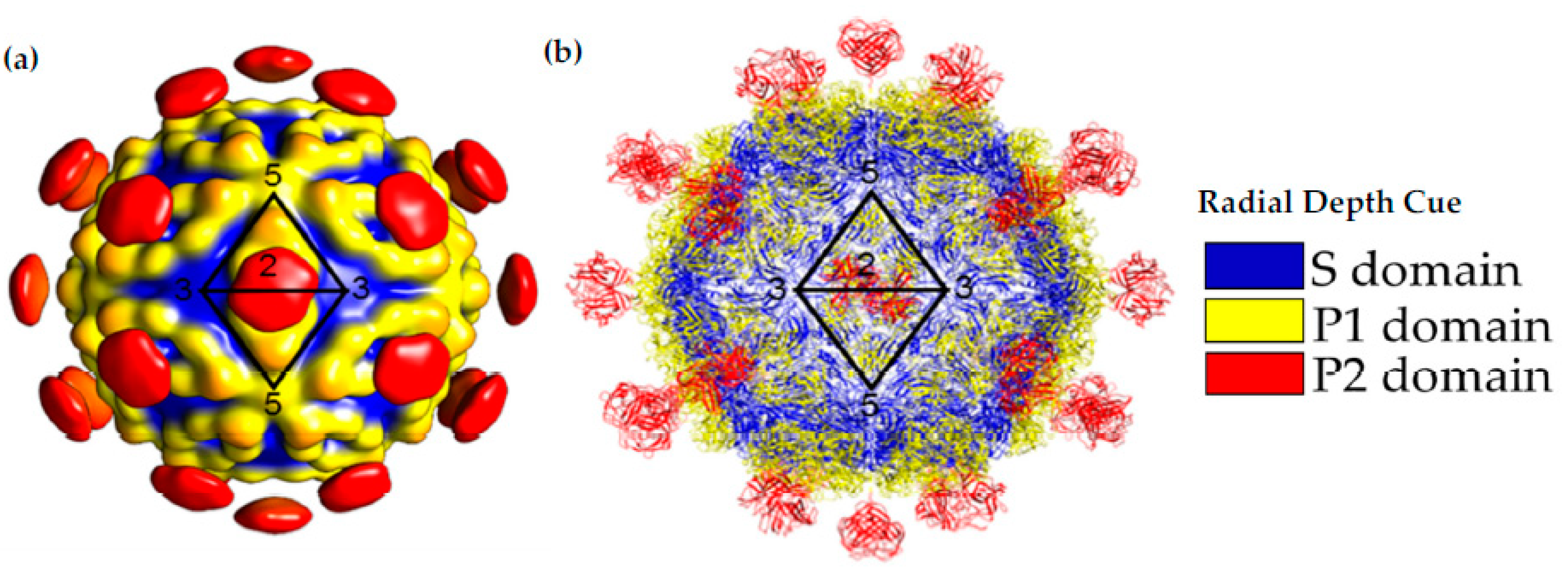
3. Capsid Structures and Modelling Provide Insights into Serotype Variability
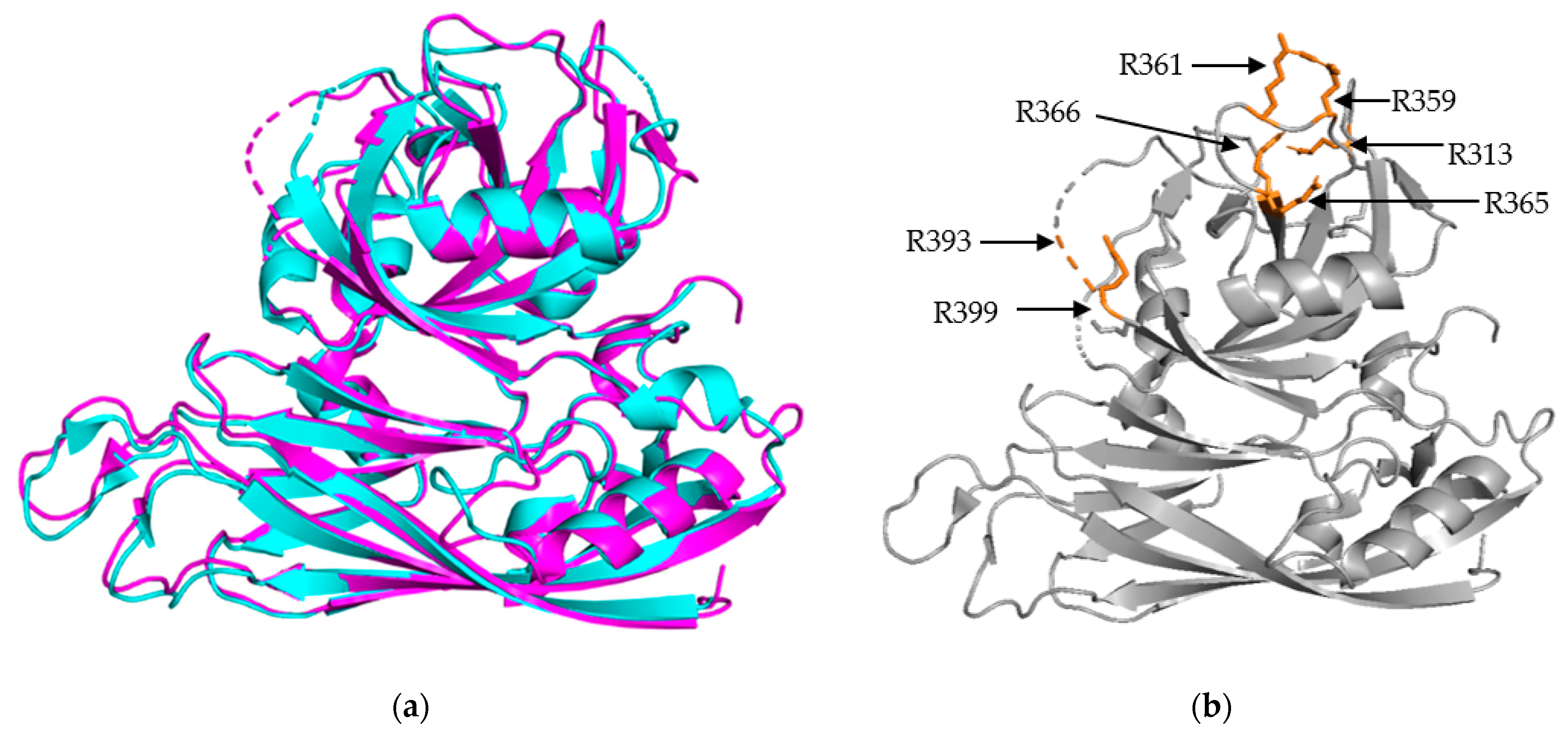
3.1. S/P1 Core Domain
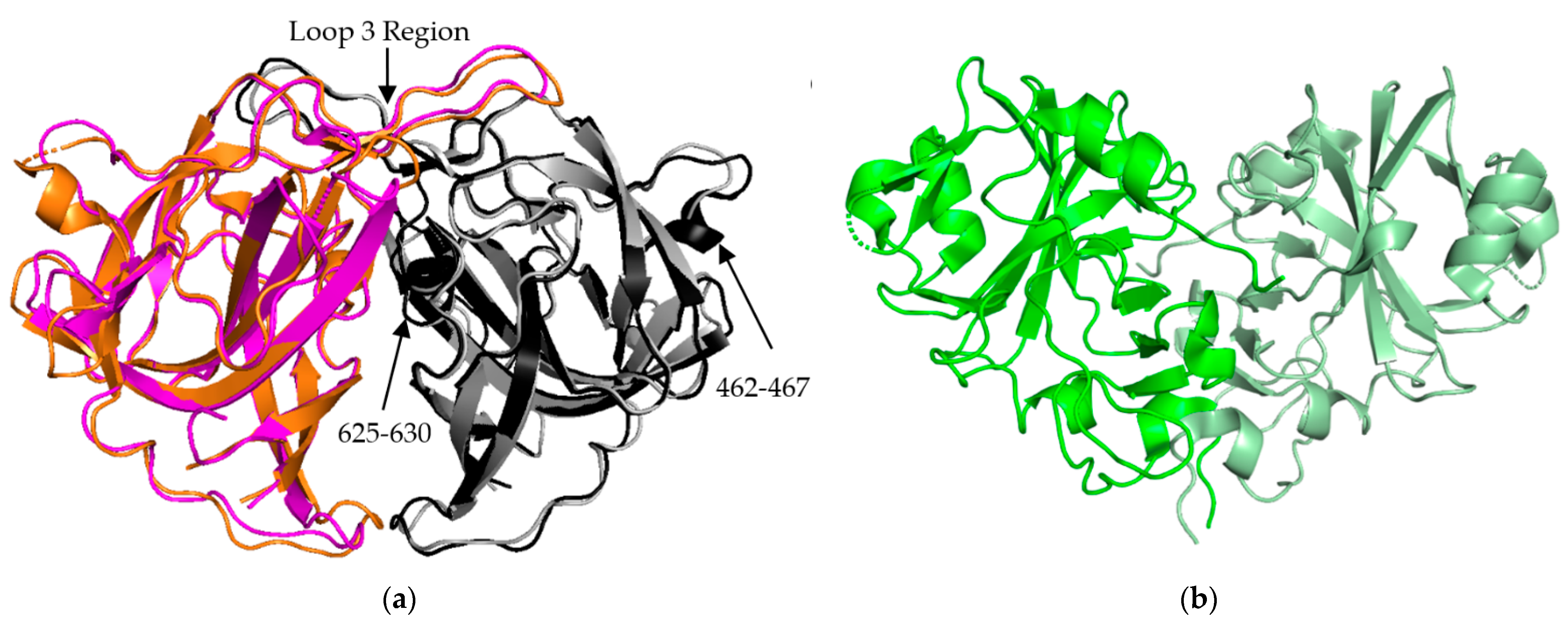
3.2. P2 Spike Domain
4. Capsid Architecture Changes during the Virus Life Cycle
5. Capsid Receptor Binding and Entry
6. Capsid/Membrane Interactions in Viral Assembly and Entry
7. Capsid-Antibody Interactions
8. Engineering and Practical Applications of AstV Structural Proteins and VLPs
9. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Donato, C.; Vijaykrishna, D. The Broad Host Range and Genetic Diversity of Mammalian and Avian Astroviruses. Viruses 2017, 9, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.; Hargest, V.; Cortez, V.; Meliopoulos, V.A.; Schultz-Cherry, S. Astrovirus Pathogenesis. Viruses 2017, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, S.; Guix, S.; El-Senousy, W.M.; Calicó, I.; Pintó, R.M.; Bosch, A. Persistent Gastroenteritis in Children Infected with Astrovirus: Association with Serotype-3 Strains. J. Med. Virol. 2003, 71, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Cordey, S.; Brito, F.; Vu, D.-L.; Turin, L.; Kilowoko, M.; Kyungu, E.; Genton, B.; Zdobnov, E.M.; D’Acremont, V.; Kaiser, L. Astrovirus VA1 Identified by Next-Generation Sequencing in a Nasopharyngeal Specimen of a Febrile Tanzanian Child with Acute Respiratory Disease of Unknown Etiology. Emerg. Microbes Infect. 2016, 5, e67. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, S.R.; Le, B.-M.; Holtz, L.R.; Storch, G.A.; Wang, D. Detection of Newly Described Astrovirus MLB1 in Stool Samples from Children. Emerg. Infect. Dis. 2009, 15, 441–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumthip, K.; Khamrin, P.; Ushijima, H.; Maneekarn, N. Molecular Epidemiology of Classic, MLB and VA Astroviruses Isolated from <5 Year-Old Children with Gastroenteritis in Thailand, 2011–2016. Infect. Genet. Evol. 2018, 65, 373–379. [Google Scholar] [CrossRef]
- Blomström, A.-L.; Widén, F.; Hammer, A.-S.; Belák, S.; Berg, M. Detection of a Novel Astrovirus in Brain Tissue of Mink Suffering from Shaking Mink Syndrome by Use of Viral Metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef] [Green Version]
- Bulbule, N.R.; Mandakhalikar, K.D.; Kapgate, S.S.; Deshmukh, V.V.; Schat, K.A.; Chawak, M.M. Role of Chicken Astrovirus as a Causative Agent of Gout in Commercial Broilers in India. Avian Pathol. 2013, 42, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Schlottau, K.; Schulze, C.; Bilk, S.; Hanke, D.; Höper, D.; Beer, M.; Hoffmann, B. Detection of a Novel Bovine Astrovirus in a Cow with Encephalitis. Transbound. Emerg. Dis. 2016, 63, 253–259. [Google Scholar] [CrossRef]
- Monroe, S.S.; Jiang, B.; Stine, S.E.; Koopmans, M.; Glass, R.I. Subgenomic RNA Sequence of Human Astrovirus Supports Classification of Astroviridae as a New Family of RNA Viruses. J. Virol. 1993, 67, 3611–3614. [Google Scholar] [CrossRef] [Green Version]
- Méndez, E.; Aguirre-Crespo, G.; Zavala, G.; Arias, C.F. Association of the Astrovirus Structural Protein VP90 with Membranes Plays a Role in Virus Morphogenesis. J. Virol. 2007, 81, 10649–10658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guix, S.; Bosch, A.; Ribes, E.; Dora Martínez, L.; Pintó, R.M. Apoptosis in Astrovirus-Infected CaCo-2 Cells. Virology 2004, 319, 249–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez, E.; Salas-Ocampo, E.; Arias, C.F. Caspases Mediate Processing of the Capsid Precursor and Cell Release of Human Astroviruses. J. Virol. 2004, 78, 8601–8608. [Google Scholar] [CrossRef] [Green Version]
- Geigenmüller, U.; Ginzton, N.H.; Matsui, S.M. Studies on Intracellular Processing of the Capsid Protein of Human Astrovirus Serotype 1 in Infected Cells. J. Gen. Virol. 2002, 83, 1691–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, A.; Tulasne, D. Caspase Cleavage of Viral Proteins, Another Way for Viruses to Make the Best of Apoptosis. Cell Death Dis. 2012, 3, e277. [Google Scholar] [CrossRef]
- Toh, Y.; Harper, J.; Dryden, K.A.; Yeager, M.; Arias, C.; Mendez, E.; Tao, Y.J. Crystal Structure of the Human Astrovirus Capsid Protein. J. Virol. 2016, 90, 9008–9017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Hernández, N.; López, S.; Arias, C.F. Minimal Capsid Composition of Infectious Human Astrovirus. Virology 2018, 521, 58–61. [Google Scholar] [CrossRef]
- del Banos-Lara, M.R.; Méndez, E. Role of Individual Caspases Induced by Astrovirus on the Processing of Its Structural Protein and Its Release from the Cell through a Non-Lytic Mechanism. Virology 2010, 401, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Moser, L.A.; Carter, M.; Schultz-Cherry, S. Astrovirus Increases Epithelial Barrier Permeability Independently of Viral Replication. J. Virol. 2007, 81, 11937–11945. [Google Scholar] [CrossRef] [Green Version]
- Méndez, E.; Fernández-Luna, T.; López, S.; Méndez-Toss, M.; Arias, C.F. Proteolytic Processing of a Serotype 8 Human Astrovirus ORF2 Polyprotein. J. Virol. 2002, 76, 7996–8002. [Google Scholar] [CrossRef] [Green Version]
- Bass, D.M.; Qiu, S. Proteolytic Processing of the Astrovirus Capsid. J. Virol. 2000, 74, 1810–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drozdetskiy, A.; Cole, C.; Procter, J.; Barton, G.J. JPred4: A Protein Secondary Structure Prediction Server. Nucleic Acids Res. 2015, 43, W389–W394. [Google Scholar] [CrossRef] [PubMed]
- York, R.L.; Yousefi, P.A.; Bogdanoff, W.; Haile, S.; Tripathi, S.; DuBois, R.M. Structural, Mechanistic, and Antigenic Characterization of the Human Astrovirus Capsid. J. Virol. 2016, 90, 2254–2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Dong, L.; Méndez, E.; Tao, Y. Crystal Structure of the Human Astrovirus Capsid Spike. Proc. Natl. Acad. Sci. USA 2011, 108, 12681–12686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuBois, R.M.; Freiden, P.; Marvin, S.; Reddivari, M.; Heath, R.J.; White, S.W.; Schultz-Cherry, S. Crystal Structure of the Avian Astrovirus Capsid Spike. J. Virol. 2013, 87, 7853–7863. [Google Scholar] [CrossRef] [Green Version]
- Khayat, R.; Johnson, J.E. Pass the Jelly Rolls. Structure 2011, 19, 904–906. [Google Scholar] [CrossRef] [Green Version]
- Dryden, K.A.; Tihova, M.; Nowotny, N.; Matsui, S.M.; Mendez, E.; Yeager, M. Immature and Mature Human Astrovirus: Structure, Conformational Changes, and Similarities to Hepatitis E Virus. J. Mol. Biol. 2012, 422, 650–658. [Google Scholar] [CrossRef]
- Bogdanoff, W.A.; Campos, J.; Perez, E.I.; Yin, L.; Alexander, D.L.; DuBois, R.M. Structure of a Human Astrovirus Capsid-Antibody Complex and Mechanistic Insights into Virus Neutralization. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Bosch, A.; Pintó, R.M.; Guix, S. Human Astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [Green Version]
- Koci, M.D.; Schultz-Cherry, S. Avian Astroviruses. Avian Pathol. 2002, 31, 213–227. [Google Scholar] [CrossRef] [Green Version]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus Infections in Humans and Animals—Molecular Biology, Genetic Diversity, and Interspecies Transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Smyth, V.J.; Todd, D.; Trudgett, J.; Lee, A.; Welsh, M.D. Capsid Protein Sequence Diversity of Chicken Astrovirus. Avian Pathol. 2012, 41, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lum, S.H.; Turner, A.; Guiver, M.; Bonney, D.; Martland, T.; Davies, E.; Newbould, M.; Brown, J.; Morfopoulou, S.; Breuer, J.; et al. An Emerging Opportunistic Infection: Fatal Astrovirus (VA1/HMO-C) Encephalitis in a Pediatric Stem Cell Transplant Recipient. Transpl. Infect. Dis. 2016, 18, 960–964. [Google Scholar] [CrossRef]
- De Grazia, S.; Lanave, G.; Bonura, F.; Urone, N.; Cappa, V.; Li Muli, S.; Pepe, A.; Gellért, A.; Banyai, K.; Martella, V.; et al. Molecular Evolutionary Analysis of Type-1 Human Astroviruses Identifies Putative Sites under Selection Pressure on the Capsid Protein. Infect. Genet. Evol. 2018, 58, 199–208. [Google Scholar] [CrossRef]
- De Nova-Ocampo, M.; Soliman, M.C.; Espinosa-Hernández, W.; Velez-del Valle, C.; Salas-Benito, J.; Valdés-Flores, J.; García-Morales, L. Human Astroviruses: In Silico Analysis of the Untranslated Region and Putative Binding Sites of Cellular Proteins. Mol. Biol. Rep. 2018. [Google Scholar] [CrossRef] [PubMed]
- Dalton, R.M.; Pastrana, E.P.; Sánchez-Fauquier, A. Vaccinia Virus Recombinant Expressing an 87-Kilodalton Polyprotein That Is Sufficient To Form Astrovirus-Like Particles. J. Virol. 2003, 77, 9094–9098. [Google Scholar] [CrossRef] [Green Version]
- Caballero, S.; Guix, S.; Ribes, E.; Bosch, A.; Pintó, R.M.; Pinto, R.M. Structural Requirements of Astrovirus Virus-Like Particles Assembled in Insect Cells Structural Requirements of Astrovirus Virus-Like Particles Assembled in Insect Cells. J. Virol. 2004, 78, 13285–13292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishna, N.K. Identification of Structural Domains Involved in Astrovirus Capsid Biology. Viral Immunol. 2005, 18, 17–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez, E.; Muñoz-Yañez, C.; Martín, C.S.-S.; Aguirre-Crespo, G.; del Baños-Lara, M.R.; Gutierrez, M.; Espinosa, R.; Acevedo, Y.; Arias, C.F.; López, S. Characterization of Human Astrovirus Cell Entry. J. Virol. 2014, 88, 2452–2460. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Hernández, N.; Meyer, L.; López, S.; DuBois, R.M.; Arias, C.F. Protein Disulfide Isomerase A4 Is Involved in Genome Uncoating during Human Astrovirus Cell Entry. Viruses 2021, 13, 53. [Google Scholar] [CrossRef]
- Galligan, J.J.; Petersen, D.R. The Human Protein Disulfide Isomerase Gene Family. Hum. Genomics 2012, 6, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, C.S.; Dey, D.; Ghosh, S.; Banerjee, M. Breach: Host Membrane Penetration and Entry by Nonenveloped Viruses. Trends Microbiol. 2018, 26, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Marvin, S.A.; Huerta, C.T.; Sharp, B.; Freiden, P.; Cline, T.D.; Schultz-Cherry, S. Type I Interferon Response Limits Astrovirus Replication and Protects against Increased Barrier Permeability In Vitro and In Vivo. J. Virol. 2016, 90, 1988–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marczinke, B.; Bloys, A.J.; Brown, T.D.; Willcocks, M.M.; Carter, M.J.; Brierley, I. The Human Astrovirus RNA-Dependent RNA Polymerase Coding Region Is Expressed by Ribosomal Frameshifting. J. Virol. 1994, 68, 5588–5595. [Google Scholar] [CrossRef] [Green Version]
- Lulla, V.; Firth, A.E. A Hidden Gene in Astroviruses Encodes a Viroporin. Nat. Commun. 2020, 11, 4070. [Google Scholar] [CrossRef]
- Burbelo, P.D.; Ching, K.H.; Esper, F.; Iadarola, M.J.; Delwart, E.; Lipkin, W.I.; Kapoor, A. Serological Studies Confirm the Novel Astrovirus HMOAstV-C as a Highly Prevalent Human Infectious Agent. PLoS ONE 2011, 6, e22576. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.K.; Matson, D.O.; Cubitt, W.D.; Jackson, L.J.; Willcocks, M.M.; Pickering, L.K.; Carter, M.J. Prevalence of Antibodies to Astrovirus Types 1 and 3 in Children and Adolescents in Norfolk, Virginia. Pediatr. Infect. Dis. J. 1999, 18, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Bogdanoff, W.A.; Morgenstern, D.; Bern, M.; Ueberheide, B.M.; Sanchez-Fauquier, A.; DuBois, R.M. De Novo Sequencing and Resurrection of a Human Astrovirus-Neutralizing Antibody. ACS Infect. Dis. 2016, 2, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Fauquier, A.; Carrascosa, A.L.; Carrascosa, J.L.; Otero, A.; Glass, R.I.; Lopez, J.A.; San Martin, C.; Melero, J.A. Characterization of a Human Astrovirus Serotype 2 Structural Protein (VP26) That Contains an Epitope Involved in Virus Neutralization. Virology 1994, 201, 312–320. [Google Scholar] [CrossRef]
- Bogdanoff, W.A.; Perez, E.I.; López, T.; Arias, C.F.; DuBois, R.M. Structural Basis for Escape of Human Astrovirus from Antibody Neutralization: Broad Implications for Rational Vaccine Design. J. Virol. 2018, 92, e01546-17. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, R.; López, T.; Bogdanoff, W.A.; Espinoza, M.A.; López, S.; DuBois, R.M.; Arias, C.F. Isolation of Neutralizing Monoclonal Antibodies to Human Astrovirus and Characterization of Virus Variants That Escape Neutralization. J. Virol. 2019, 93, e01465-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, M.; Wei, C.; Wang, L.; Cao, D.; Meng, X.-J.; Jiang, X.; Tan, M. A Trivalent Vaccine Candidate against Hepatitis E Virus, Norovirus, and Astrovirus. Vaccine 2016, 34, 905–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidokhti, M.R.M.; Ullman, K.; Jensen, T.H.; Chriél, M.; Mottahedin, A.; Munir, M.; Andersson, A.M.; Detournay, O.; Hammer, A.S.; Baule, C. Establishment of Stably Transfected Cells Constitutively Expressing the Full-Length and Truncated Antigenic Proteins of Two Genetically Distinct Mink Astroviruses. PLoS ONE 2013, 8, e82978. [Google Scholar] [CrossRef] [PubMed]
- Sellers, H.; Linneman, E.; Icard, A.H.; Mundt, E. A Purified Recombinant Baculovirus Expressed Capsid Protein of a New Astrovirus Provides Partial Protection to Runting–Stunting Syndrome in Chickens. Vaccine 2010, 28, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Hewson, K.A.; Wawegama, N.K.; O’Rourke, D.; Putri, K.; Chamings, A.; Noormohammadi, A.H.; Ignjatovic, J. The C-Terminal End of the Capsid Protein of Avian Nephritis Virus Is Antigenic and Induces Broadly Cross-Reactive Antibodies. J. Virol. Methods 2015, 221, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Jaime, C. Astrovirus Reverse Genetics Systems, a Story of Success. Curr. Opin. Virol. 2020, 44, 57–65. [Google Scholar] [CrossRef]
- Velázquez-Moctezuma, R.; Baños-Lara, M.R.; Acevedo, Y.; Méndez, E. Alternative Cell Lines to Improve the Rescue of Infectious Human Astrovirus from a CDNA Clone. J. Virol. Methods 2012, 179, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Jiang, X. Norovirus Capsid Protein-Derived Nanoparticles and Polymers as Versatile Platforms for Antigen Presentation and Vaccine Development. Pharmaceutics 2019, 11, 472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guenther, C.M.; Brun, M.J.; Bennett, A.D.; Ho, M.L.; Chen, W.; Zhu, B.; Lam, M.; Yamagami, M.; Kwon, S.; Bhattacharya, N.; et al. Protease-Activatable Adeno-Associated Virus Vector for Gene Delivery to Damaged Heart Tissue. Mol. Ther. 2019, 27, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Judd, J.; Ho, M.L.; Tiwari, A.; Gomez, E.J.; Dempsey, C.; Van Vliet, K.; Igoshin, O.A.; Silberg, J.J.; Agbandje-McKenna, M.; Suh, J. Tunable Protease-Activatable Virus Nanonodes. ACS Nano 2014, 8, 4740–4746. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ykema, M.; Tao, Y.J. Structural Insights into the Human Astrovirus Capsid. Viruses 2021, 13, 821. https://doi.org/10.3390/v13050821
Ykema M, Tao YJ. Structural Insights into the Human Astrovirus Capsid. Viruses. 2021; 13(5):821. https://doi.org/10.3390/v13050821
Chicago/Turabian StyleYkema, Matthew, and Yizhi J. Tao. 2021. "Structural Insights into the Human Astrovirus Capsid" Viruses 13, no. 5: 821. https://doi.org/10.3390/v13050821
APA StyleYkema, M., & Tao, Y. J. (2021). Structural Insights into the Human Astrovirus Capsid. Viruses, 13(5), 821. https://doi.org/10.3390/v13050821