
Human Astrovirus 1–8 Seroprevalence Evaluation in a United States Adult Population


Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Astrovirus Capsid Sequences Used for Sequence Identity Studies
2.2. Reagents and Supplies
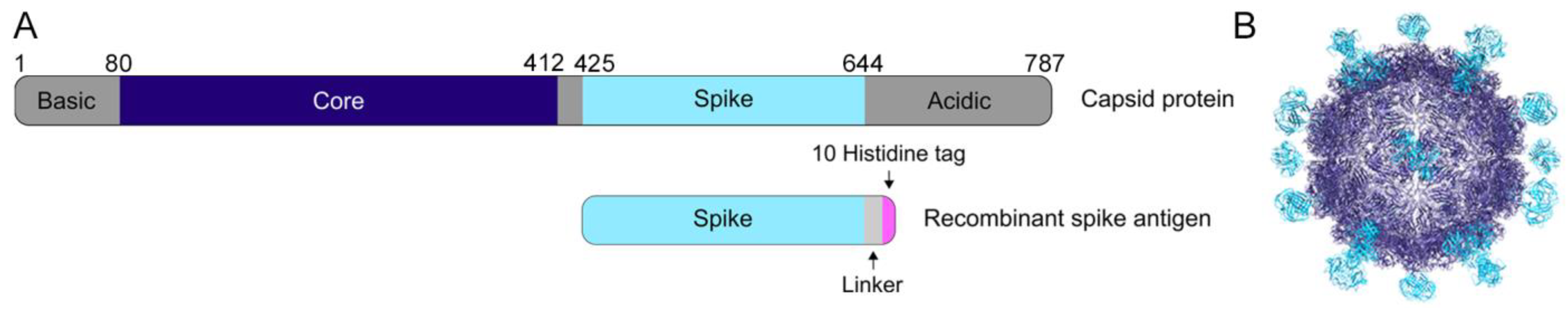
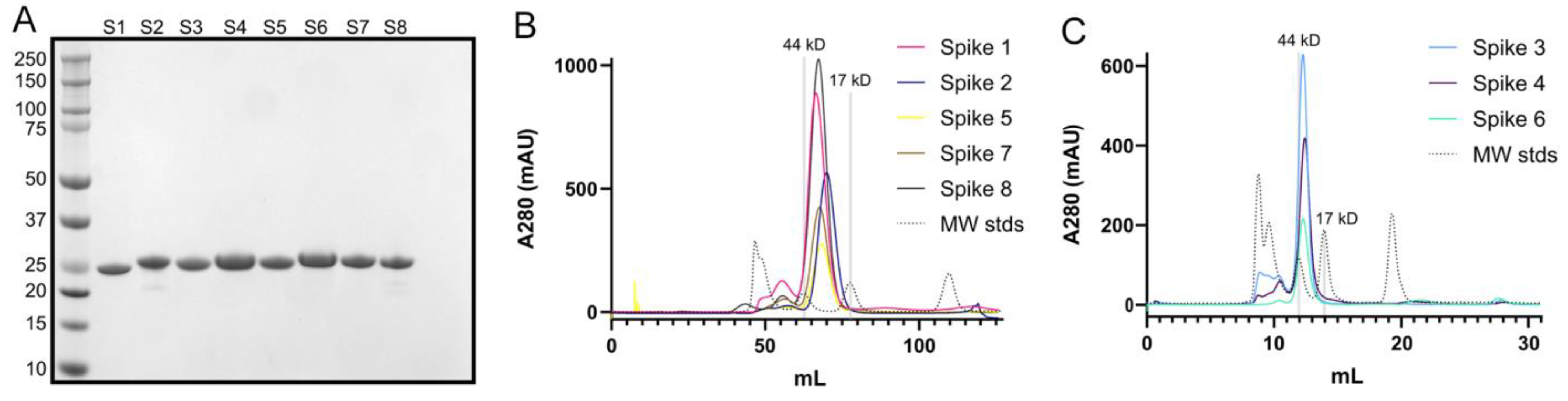
2.3. Expression and Purification of Human Astrovirus Spikes 1–8
2.4. Expression and Purification of the SARS-CoV-2 RBD
2.5. Expression and Purification of Recombinant mAb 3B4
2.6. Generation of Rabbit Polyclonal Serum to Human Astrovirus 1
2.7. Human Plasma Samples
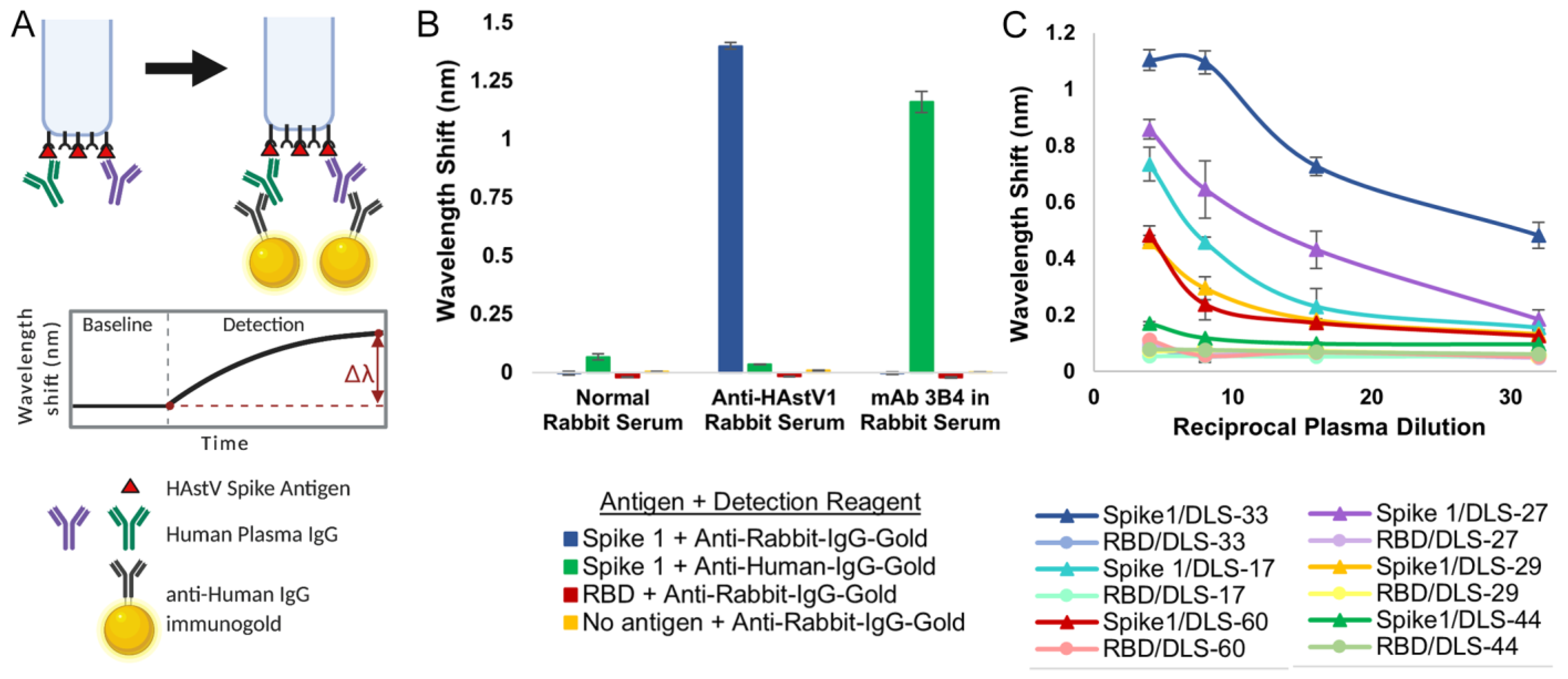
2.8. Biolayer Interferometry Immunosorbent Assay
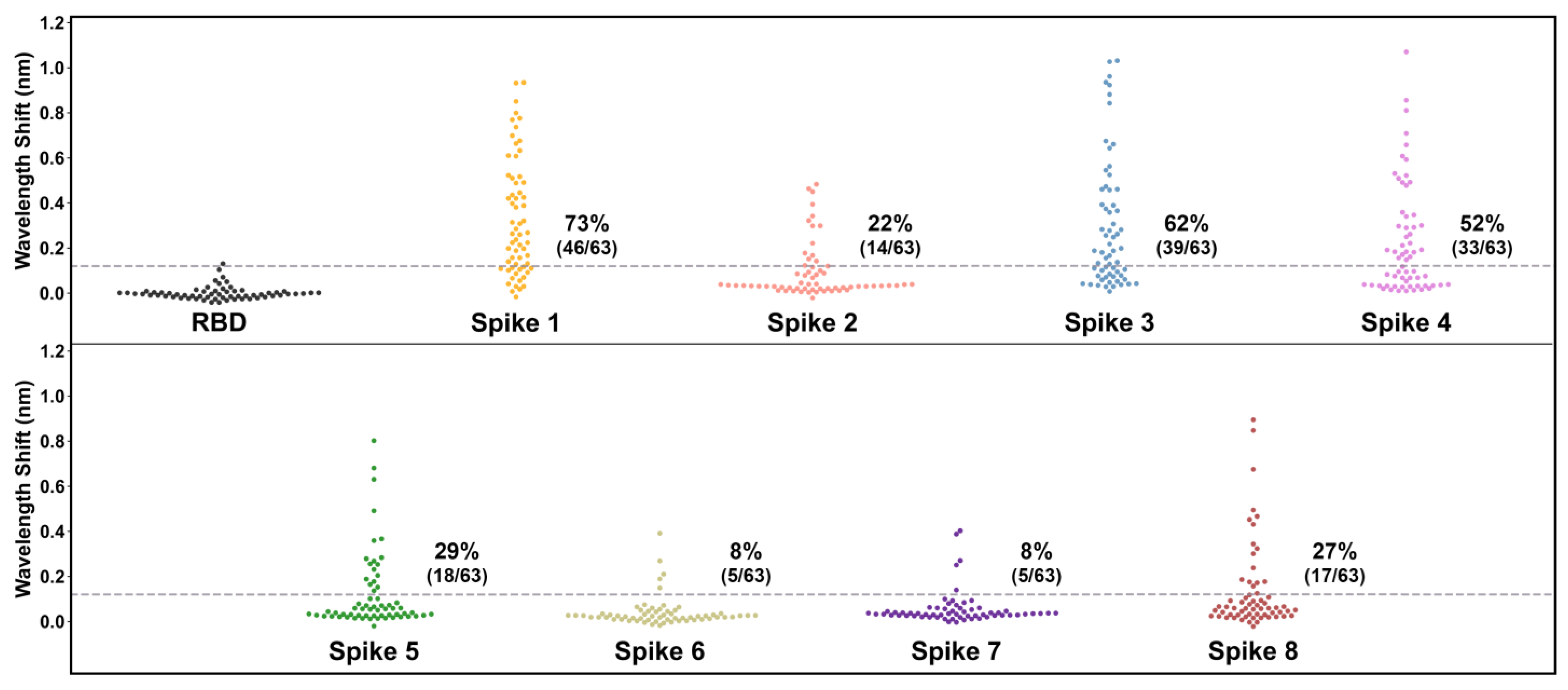
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mendez, E.; Arias, C.F. Astroviruses. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Cohen, J.I., Griffin, D.E., Lamb, R.A., Martin, M.A., Racaniello, V.R., Bernard, R., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 1, pp. 609–628. [Google Scholar]
- Koci, M.D.; Schultz-Cherry, S. Avian astroviruses. Avian Pathol. 2002, 31, 213–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woode, G.N.; Pohlenz, J.F.; Gourley, N.E.; Fagerland, J.A. Astrovirus and breda virus infections of dome cell epithelium of bovine ileum. J. Clin. Microbiol. 1984, 19, 623–630. [Google Scholar] [CrossRef] [Green Version]
- Cortez, V.; Sharp, B.; Yao, J.; Livingston, B.; Vogel, P.; Schultz-Cherry, S. Characterizing a murine model for astrovirus using viral isolates from persistently infected immunocompromised mice. J. Virol. 2019, 93, e00223-19. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.R.; Morfopoulou, S.; Hubb, J.; Emmett, W.A.; Ip, W.; Shah, D.; Brooks, T.; Paine, S.M.; Anderson, G.; Virasami, A.; et al. Astrovirus va1/hmo-c: An increasingly recognized neurotropic pathogen in immunocompromised patients. Clin. Infect. Dis. 2015, 60, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Naccache, S.N.; Peggs, K.S.; Mattes, F.M.; Phadke, R.; Garson, J.A.; Grant, P.; Samayoa, E.; Federman, S.; Miller, S.; Lunn, M.P.; et al. Diagnosis of neuroinvasive astrovirus infection in an immunocompromised adult with encephalitis by unbiased next-generation sequencing. Clin. Infect. Dis. 2015, 60, 919–923. [Google Scholar] [CrossRef] [Green Version]
- Quan, P.L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D.; et al. Astrovirus encephalitis in boy with x-linked agammaglobulinemia. Emergy Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef]
- Cordey, S.; Vu, D.L.; Schibler, M.; L’Huillier, A.G.; Brito, F.; Docquier, M.; Posfay-Barbe, K.M.; Petty, T.J.; Turin, L.; Zdobnov, E.M.; et al. Astrovirus mlb2, a new gastroenteric virus associated with meningitis and disseminated infection. Emergy Infect. Dis. 2016, 22, 846–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A.M.Q.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Virus taxonomy: Classification and nomenclature of viruses. In Ninth Report of the International Committee on Taxonomy of Viruses, 9th ed.; Elsevier: Philadelphia, PA, USA, 2011. [Google Scholar]
- Koopmans, M.P.; Bijen, M.H.; Monroe, S.S.; Vinje, J. Age-stratified seroprevalence of neutralizing antibodies to astrovirus types 1 to 7 in humans in the netherlands. Clin. Diagn. Lab. Immunol. 1998, 5, 33–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtz, J.B.; Lee, T.W. Human astrovirus serotypes. Lancet 1984, 2, 1405. [Google Scholar] [CrossRef]
- Cortez, V.; Meliopoulos, V.A.; Karlsson, E.A.; Hargest, V.; Johnson, C.; Schultz-Cherry, S. Astrovirus biology and pathogenesis. Annu. Rev. Virol. 2017, 4, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Dennehy, P.H.; Nelson, S.M.; Spangenberger, S.; Noel, J.S.; Monroe, S.S.; Glass, R.I. A prospective case-control study of the role of astrovirus in acute diarrhea among hospitalized young children. J. Infect. Dis. 2001, 184, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Gallimore, C.I.; Taylor, C.; Gennery, A.R.; Cant, A.J.; Galloway, A.; Xerry, J.; Adigwe, J.; Gray, J.J. Contamination of the hospital environment with gastroenteric viruses: Comparison of two pediatric wards over a winter season. J. Clin. Microbiol. 2008, 46, 3112–3115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodgame, R.W. Viral causes of diarrhea. Gastroenterol. Clin. N. Am. 2001, 30, 779–795. [Google Scholar] [CrossRef]
- Gray, J.J.; Wreghitt, T.G.; Cubitt, W.D.; Elliot, P.R. An outbreak of gastroenteritis in a home for the elderly associated with astrovirus type 1 and human calicivirus. J. Med. Virol. 1987, 23, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.C.; Lightfoot, N.F.; Cubitt, W.D.; Wilson, S.A. Outbreaks of astrovirus type 1 and rotavirus gastroenteritis in a geriatric in-patient population. J. Hosp. Infect. 1989, 14, 9–14. [Google Scholar] [CrossRef]
- Marshall, J.A.; Bruggink, L.D.; Sturge, K.; Subasinghe, N.; Tan, A.; Hogg, G.G. Molecular features of astrovirus associated with a gastroenteritis outbreak in an aged-care centre. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Olortegui, M.P.; Rouhani, S.; Yori, P.P.; Salas, M.S.; Trigoso, D.R.; Mondal, D.; Bodhidatta, L.; Platts-Mills, J.; Samie, A.; Kabir, F.; et al. Astrovirus infection and diarrhea in 8 countries. Pediatrics 2017, 141, e20171326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, J.E.; Mitchell, D.K. Astrovirus infection in children. Curr. Opin. Infect. Dis. 2003, 16, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Pinto, R.M.; Guix, S. Human astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [Green Version]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the united states. Emergy Infect. Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef]
- Kurtz, J.; Lee, T. Astrovirus gastroenteritis age distribution of antibody. Med. Microbiol. Immunol. 1978, 166, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J.B.; Lee, T.W.; Craig, J.W.; Reed, S.E. Astrovirus infection in volunteers. J. Med. Virol. 1979, 3, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.K. Astrovirus gastroenteritis. Pediatric Infect. Dis. J. 2002, 21, 1067–1069. [Google Scholar] [CrossRef] [PubMed]
- Bjorkholm, M.; Celsing, F.; Runarsson, G.; Waldenstrom, J. Successful intravenous immunoglobulin therapy for severe and persistent astrovirus gastroenteritis after fludarabine treatment in a patient with waldenstrom’s macroglobulinemia. Int. J. Hematol. 1995, 62, 117–120. [Google Scholar] [CrossRef]
- Dong, J.; Dong, L.; Mendez, E.; Tao, Y. Crystal structure of the human astrovirus capsid spike. Proc. Natl. Acad. Sci. USA 2011, 108, 12681–12686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- York, R.L.; Yousefi, P.A.; Bogdanoff, W.; Haile, S.; Tripathi, S.; DuBois, R.M. Structural, mechanistic, and antigenic characterization of the human astrovirus capsid. J. Virol. 2015, 90, 2254–2263. [Google Scholar] [CrossRef] [Green Version]
- Toh, Y.; Harper, J.; Dryden, K.A.; Yeager, M.; Arias, C.F.; Mendez, E.; Tao, Y.J. Crystal structure of the human astrovirus capsid protein. J. Virol. 2016, 90, 9008–9017. [Google Scholar] [CrossRef] [Green Version]
- Bogdanoff, W.A.; Perez, E.I.; Lopez, T.; Arias, C.F.; DuBois, R.M. Structural basis for escape of human astrovirus from antibody neutralization: Broad implications for rational vaccine design. J. Virol. 2018, 92, e01546-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, D.K.; Matson, D.O.; Cubitt, W.D.; Jackson, L.J.; Willcocks, M.M.; Pickering, L.K.; Carter, M.J. Prevalence of antibodies to astrovirus types 1 and 3 in children and adolescents in norfolk, virginia. Pediatric Infect. Dis. J. 1999, 18, 249–254. [Google Scholar] [CrossRef]
- Kriston, S.; Willcocks, M.M.; Carter, M.J.; Cubitt, W.D. Seroprevalence of astrovirus types 1 and 6 in london, determined using recombinant virus antigen. Epidemiol. Infect. 1996, 117, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Kobayashi, M.; Araki, K.; Shinozaki, T.; Yanagawa, Y. Antibody prevalence against astrovirus according to age groups. Kansenshogaku Zasshi 1999, 73, 578–583. [Google Scholar] [CrossRef] [Green Version]
- Holtz, L.R.; Bauer, I.K.; Jiang, H.; Belshe, R.; Freiden, P.; Schultz-Cherry, S.L.; Wang, D. Seroepidemiology of astrovirus mlb1. Clin. Vaccine Immunol. 2014, 21, 908–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burbelo, P.D.; Ching, K.H.; Esper, F.; Iadarola, M.J.; Delwart, E.; Lipkin, W.I.; Kapoor, A. Serological studies confirm the novel astrovirus hmoastv-c as a highly prevalent human infectious agent. PLoS ONE 2011, 6, e22576. [Google Scholar] [CrossRef] [PubMed]
- Meliopoulos, V.A.; Kayali, G.; Burnham, A.; Oshansky, C.M.; Thomas, P.G.; Gray, G.C.; Beck, M.A.; Schultz-Cherry, S. Detection of antibodies against turkey astrovirus in humans. PLoS ONE 2014, 9, e96934. [Google Scholar] [CrossRef] [PubMed]
- Dzimianski, J.V.; Lorig-Roach, N.; O’Rourke, S.M.; Alexander, D.L.; Kimmey, J.M.; DuBois, R.M. Rapid and sensitive detection of sars-cov-2 antibodies by biolayer interferometry. Sci. Rep. 2020, 10, 21738. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, R.; López, T.; Bogdanoff, W.A.; Espinoza, M.A.; López, S.; DuBois, R.M.; Arias, C.F. Isolation of neutralizing monoclonal antibodies to human astrovirus and characterization of virus variants that escape neutralization. J. Virol. 2018, 93, e01465-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, L.; López, T.; Espinosa, R.; Arias, C.F.; Vollmers, C.; DuBois, R.M. A simplified workflow for monoclonal antibody sequencing. PLoS ONE 2019, 14, e0218717. [Google Scholar] [CrossRef]
- Wu, X.; Yang, Z.Y.; Li, Y.; Hogerkorp, C.M.; Schief, W.R.; Seaman, M.S.; Zhou, T.; Schmidt, S.D.; Wu, L.; Xu, L.; et al. Rational design of envelope identifies broadly neutralizing human monoclonal antibodies to hiv-1. Science 2010, 329, 856–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bass, D.M.; Upadhyayula, U. Characterization of human serotype 1 astrovirus-neutralizing epitopes. J. Virol. 1997, 71, 8666–8671. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Fauquier, A.; Carrascosa, A.L.; Carrascosa, J.L.; Otero, A.; Glass, R.I.; Lopez, J.A.; San Martin, C.; Melero, J.A. Characterization of a human astrovirus serotype 2 structural protein (vp26) that contains an epitope involved in virus neutralization. Virology 1994, 201, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Bogdanoff, W.A.; Campos, J.; Perez, E.I.; Yin, L.; Alexander, D.L.; DuBois, R.M. Structure of a human astrovirus capsid-antibody complex and mechanistic insights into virus neutralization. J. Virol. 2016, 91, e01859-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Core 1 | Core 2 | Core 3 | Core 4 | Core 5 | Core 6 | Core 7 | Core 8 | |
|---|---|---|---|---|---|---|---|---|
| Core 1 | 100% | 86.7% | 91.5% | 86.1% | 90.0% | 90.9% | 87.9% | 86.7% |
| Core 2 | - | 100% | 86.7% | 87.9% | 83.3% | 83.9% | 85.2% | 87.9% |
| Core 3 | - | - | 100% | 87.3% | 90.6% | 91.2% | 92.8% | 88.2% |
| Core 4 | - | - | - | 100% | 84.6% | 86.1% | 84.9% | 97.0% |
| Core 5 | - | - | - | - | 100% | 90.3% | 88.8% | 85.8% |
| Core 6 | - | - | - | - | - | 100% | 87.9% | 87.6% |
| Core 7 | - | - | - | - | - | - | 100% | 85.8% |
| Core 8 | - | - | - | - | - | - | - | 100% |
| Spike 1 | Spike 2 | Spike 3 | Spike 4 | Spike 5 | Spike 6 | Spike 7 | Spike 8 | |
|---|---|---|---|---|---|---|---|---|
| Spike 1 | 100% | 50.2% | 61.2% | 41.4% | 47.0% | 53.1% | 58.9% | 53.0% |
| Spike 2 | - | 100% | 46.7% | 47.9% | 41.8% | 47.0% | 44.9% | 49.8% |
| Spike 3 | - | - | 100% | 46.3% | 48.6% | 54.7% | 75.7% | 55.1% |
| Spike 4 | - | - | - | 100% | 46.0% | 47.0% | 43.5% | 46.1% |
| Spike 5 | - | - | - | - | 100% | 63.34% | 52.8% | 52.1% |
| Spike 6 | - | - | - | - | - | 100% | 56.6% | 57.3% |
| Spike 7 | - | - | - | - | - | - | 100% | 53.3% |
| Spike 8 | - | - | - | - | - | - | - | 100% |
| Study Year Published | Seroprevalence | Size | Antigen |
|---|---|---|---|
| Koopmans et al. [10] 1998 | Ages 0–79: Human astrovirus 1: 91% Human astrovirus 2: 31% Human astrovirus 3: 69% Human astrovirus 4: 56% Human astrovirus 5: 36% Human astrovirus 6: 16% Human astrovirus 7: 10% Human astrovirus 8: --- | 242 people ages 0–79 | Human astrovirus particles cultured in Caco-2 cells |
| Mitchell et al. [31] 1999 | By age 9: Human astrovirus 1: 94% Human astrovirus 3: 42% | 393 children | Baculovirus-expressed recombinant capsid proteins |
| Kriston et al. [32] 1996 | By age 5: Human astrovirus 1: 90% Human astrovirus 6: 10–30% | 273 children and hospital staff | Baculovirus-expressed recombinant capsid proteins |
| Kobayashi et al. [33] 1999 | By age 3: Human astrovirus 1: ~100% Human astrovirus 2: ~100% | 170 children | Unclear: Article in Japanese |
| Kurtz et al. [23] 1978 | Presence of “astrovirus antibody” By 6 months: 45% By age 1: 7% By age 2: 25% By age 4: 71% By age 10: 75% Young adult cohort: 77% | 87 children ages 0–10 70 young adults ages 17–30 | Human astrovirus particles cultured in HEK cells |
| Holtz et al. [34] 2014 | By age 17: MLB1: 100% | 395 people ages 0–95 | Baculovirus-expressed recombinant capsid protein |
| Burbelo et al. [35] 2011 | HMOAstV-C (VA1): 36% by age 10 65% in adults | 103 children and 106 adults | Crude protein extracts containing N- and C-terminal capsid fragments |
| Meliopoulos et al. [36] 2014 | By adulthood: Human astrovirus 1: 81% Turkey astrovirus 2: 26% | 160 turkey growers, turkey meat processing plant workers, and unexposed workers | Baculovirus-expressed recombinant capsid proteins |
| Controls | Human Astrovirus Spike Antigens (Background-Corrected) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ID | No Antigen | RBD | Spike 1 | Spike 2 | Spike 3 | Spike 4 | Spike 5 | Spike 6 | Spike 7 | Spike 8 | |
| 1 | DLS-18 | 0.13 | 0.06 | 0.11 | 0.04 | 1.03 | 0.03 | 0.02 | 0.00 | 0.05 | 0.03 |
| 2 | DLS-14 | 0.10 | 0.05 | 0.20 | 0.03 | 0.88 | 0.15 | 0.03 | 0.03 | 0.09 | 0.07 |
| 3 | DLS-55 | 0.07 | 0.06 | 0.32 | 0.15 | 0.92 | 0.04 | 0.04 | 0.06 | 0.10 | 0.18 |
| 4 | DLS-44 | 0.05 | 0.04 | 0.02 | 0.03 | 0.46 | 0.15 | 0.02 | 0.05 | 0.04 | 0.17 |
| 5 | DLS-36 | 0.14 | 0.07 | 0.01 | 0.09 | 0.54 | 0.03 | 0.01 | 0.01 | 0.03 | 0.06 |
| 6 | DLS-51 | 0.08 | 0.07 | 0.16 | 0.02 | 0.66 | 0.01 | 0.02 | 0.01 | 0.04 | 0.02 |
| 7 | DLS-06 | 0.11 | 0.06 | 0.11 | 0.04 | 0.67 | 0.53 | 0.06 | 0.03 | 0.05 | 0.09 |
| 8 | DLS-01 | 0.14 | 0.08 | 0.03 | 0.11 | 0.96 | 0.59 | 0.03 | 0.04 | 0.06 | 0.32 |
| 9 | DLS-30 | 0.13 | 0.09 | 0.43 | 0.02 | 0.31 | 0.09 | 0.26 | 0.03 | 0.07 | 0.07 |
| 10 | DLS-02 | 0.20 | 0.10 | 0.52 | 0.03 | 0.47 | 0.09 | 0.27 | 0.05 | 0.05 | 0.10 |
| 11 | DLS-41 | 0.10 | 0.07 | 0.68 | 0.03 | 0.52 | 0.22 | 0.28 | 0.19 | 0.08 | 0.08 |
| 12 | DLS-20 | 0.09 | 0.05 | 0.44 | 0.03 | 0.46 | 0.03 | 0.03 | 0.02 | 0.04 | 0.03 |
| 13 | DLS-05 | 0.37 | 0.20 | 0.42 | −0.02 | 0.56 | 0.07 | 0.16 | 0.00 | 0.01 | −0.02 |
| 14 | DLS-54 | 0.07 | 0.07 | 0.31 | 0.01 | 0.37 | 0.03 | 0.03 | 0.03 | 0.09 | 0.02 |
| 15 | DLS-21 | 0.22 | 0.17 | 0.27 | 0.02 | 0.46 | 0.03 | −0.02 | 0.00 | 0.04 | 0.12 |
| 16 | DLS-49 | 0.09 | 0.06 | 0.78 | 0.02 | 0.84 | 0.02 | 0.06 | 0.02 | 0.25 | 0.06 |
| 17 | DLS-35 | 0.09 | 0.07 | 0.63 | 0.14 | 0.64 | 0.19 | 0.01 | 0.01 | 0.02 | 0.01 |
| 18 | DLS-24 | 0.12 | 0.06 | 0.11 | 0.03 | 1.03 | 0.07 | 0.19 | 0.06 | 0.27 | 0.89 |
| 19 | DLS-19 | 0.08 | 0.08 | 0.12 | 0.00 | 0.18 | 0.01 | 0.14 | 0.00 | 0.39 | 0.47 |
| 20 | DLS-28 | 0.06 | 0.03 | 0.19 | 0.03 | 0.28 | 0.04 | 0.02 | 0.01 | 0.02 | 0.67 |
| 21 | DLS-42 | 0.07 | 0.06 | 0.26 | 0.03 | 0.13 | 0.01 | 0.02 | 0.01 | 0.04 | 0.43 |
| 22 | DLS-07 | 0.09 | 0.05 | 0.26 | 0.01 | 0.08 | 0.10 | 0.68 | 0.05 | 0.03 | 0.05 |
| 23 | DLS-25 | 0.10 | 0.06 | 0.85 | 0.46 | 0.05 | 0.12 | 0.63 | 0.03 | 0.03 | 0.09 |
| 24 | DLS-27 | 0.11 | 0.08 | 0.77 | 0.01 | 0.06 | 0.08 | 0.02 | −0.01 | 0.01 | 0.34 |
| 25 | DLS-34 | 0.10 | 0.07 | 0.70 | 0.01 | 0.14 | 0.03 | 0.03 | 0.00 | 0.02 | 0.45 |
| 26 | DLS-23 | 0.09 | 0.05 | 0.93 | 0.09 | 0.28 | 0.07 | 0.36 | 0.03 | 0.04 | 0.04 |
| 27 | DLS-33 | 0.10 | 0.10 | 0.93 | 0.03 | 0.08 | 0.01 | 0.03 | −0.02 | 0.00 | 0.00 |
| 28 | DLS-09 | 0.29 | 0.12 | 0.80 | 0.07 | 0.10 | 0.17 | 0.05 | 0.07 | 0.02 | 0.02 |
| 29 | DLS-53 | 0.08 | 0.06 | 0.74 | 0.03 | 0.04 | 0.29 | 0.10 | 0.06 | 0.04 | 0.08 |
| 30 | DLS-61 | 0.08 | 0.07 | 0.49 | 0.01 | 0.03 | 0.02 | 0.06 | 0.01 | 0.03 | 0.01 |
| 31 | DLS-17 | 0.09 | 0.06 | 0.61 | 0.03 | 0.19 | 0.03 | 0.02 | 0.01 | 0.04 | 0.03 |
| 32 | DLS-04 | 0.09 | 0.04 | 0.49 | 0.03 | 0.25 | 0.05 | 0.07 | 0.02 | 0.06 | 0.06 |
| 33 | DLS-46 | 0.08 | 0.06 | 0.39 | 0.02 | 0.17 | 0.03 | 0.03 | 0.02 | 0.03 | 0.02 |
| 34 | DLS-12 | 0.06 | 0.03 | 0.38 | 0.03 | 0.11 | 0.03 | 0.25 | 0.03 | 0.03 | 0.05 |
| 35 | DLS-31 | 0.16 | 0.14 | 0.40 | 0.01 | 0.07 | 0.18 | 0.08 | −0.01 | 0.01 | 0.00 |
| 36 | DLS-40 | 0.09 | 0.05 | 0.31 | 0.02 | 0.06 | 0.21 | 0.07 | 0.03 | 0.03 | 0.03 |
| 37 | DLS-26 | 0.12 | 0.08 | 0.61 | 0.04 | 0.20 | 0.26 | 0.06 | 0.00 | 0.02 | 0.17 |
| 38 | DLS-45 | 0.10 | 0.07 | 0.51 | 0.12 | 0.28 | 0.25 | 0.07 | 0.02 | 0.06 | 0.30 |
| 39 | DLS-39 | 0.13 | 0.07 | 0.45 | 0.17 | 0.10 | 0.36 | 0.02 | 0.02 | 0.06 | 0.06 |
| 40 | DLS-47 | 0.09 | 0.05 | 0.66 | 0.01 | 0.04 | 0.52 | 0.03 | 0.02 | 0.03 | 0.02 |
| 41 | DLS-59 | 0.08 | 0.07 | 0.21 | 0.02 | 0.09 | 0.49 | 0.03 | 0.03 | 0.40 | 0.02 |
| 42 | DLS-32 | 0.11 | 0.07 | 0.07 | 0.00 | 0.13 | 0.49 | 0.01 | 0.02 | 0.01 | 0.07 |
| 43 | DLS-37 | 0.11 | 0.07 | 0.09 | 0.03 | 0.36 | 0.35 | 0.05 | 0.02 | 0.04 | 0.24 |
| 44 | DLS-43 | 0.10 | 0.06 | 0.09 | 0.03 | 0.36 | 0.30 | 0.03 | 0.02 | 0.03 | 0.04 |
| 45 | DLS-15 | 0.08 | 0.04 | 0.22 | 0.08 | 0.25 | 0.34 | 0.23 | 0.21 | 0.14 | 0.16 |
| 46 | DLS-58 | 0.08 | 0.06 | 0.28 | 0.22 | 0.39 | 0.48 | 0.04 | 0.03 | 0.04 | 0.12 |
| 47 | DLS-11 | 0.08 | 0.04 | 0.07 | 0.45 | 0.03 | 0.19 | 0.18 | 0.02 | 0.03 | 0.06 |
| 48 | DLS-62 | 0.13 | 0.12 | 0.10 | 0.32 | 0.26 | 0.07 | 0.03 | 0.05 | 0.08 | 0.08 |
| 49 | DLS-57 | 0.07 | 0.06 | 0.04 | 0.04 | 0.04 | 0.16 | 0.04 | 0.02 | 0.03 | 0.19 |
| 50 | DLS-13 | 0.09 | 0.04 | 0.03 | 0.12 | 0.19 | 0.07 | 0.15 | 0.01 | 0.04 | 0.05 |
| 51 | DLS-48 | 0.09 | 0.06 | 0.06 | 0.09 | 0.22 | 0.18 | 0.07 | 0.02 | 0.03 | 0.02 |
| 52 | DLS-56 | 0.07 | 0.08 | 0.14 | 0.34 | 0.93 | 0.71 | 0.80 | 0.27 | 0.01 | 0.11 |
| 53 | DLS-63 | 0.07 | 0.08 | 0.11 | 0.48 | 0.39 | 0.51 | 0.49 | 0.15 | 0.03 | 0.04 |
| 54 | DLS-08 | 0.08 | 0.05 | 0.52 | 0.30 | 0.11 | 1.07 | 0.28 | 0.39 | 0.03 | 0.85 |
| 55 | DLS-52 | 0.22 | 0.11 | −0.02 | 0.39 | 0.01 | 0.81 | 0.01 | 0.00 | 0.00 | 0.49 |
| 56 | DLS-50 | 0.10 | 0.07 | 0.42 | 0.18 | 0.12 | 0.66 | 0.37 | 0.05 | 0.04 | 0.09 |
| 57 | DLS-03 | 0.08 | 0.04 | 0.17 | 0.04 | 0.04 | 0.85 | 0.20 | 0.07 | 0.03 | 0.05 |
| 58 | DLS-22 | 0.11 | 0.06 | 0.16 | 0.30 | 0.07 | 0.61 | 0.08 | 0.03 | 0.06 | 0.04 |
| 59 | DLS-29 | 0.08 | 0.05 | 0.20 | 0.04 | 0.04 | 0.18 | 0.06 | 0.02 | 0.03 | 0.02 |
| 60 | DLS-10 | 0.10 | 0.05 | 0.22 | 0.10 | 0.05 | 0.30 | 0.10 | 0.04 | 0.03 | 0.06 |
| 61 | DLS-16 | 0.09 | 0.04 | 0.13 | 0.08 | 0.08 | 0.29 | 0.03 | 0.02 | 0.03 | 0.03 |
| 62 | DLS-38 | 0.16 | 0.08 | 0.24 | 0.01 | 0.15 | 0.03 | 0.01 | 0.01 | 0.02 | 0.02 |
| 63 | DLS-60 | 0.05 | 0.04 | 0.13 | 0.01 | 0.03 | 0.01 | 0.04 | 0.03 | 0.02 | 0.02 |
| 64 | PBS | 0.06 | 0.05 | 0.01 | 0.02 | 0.02 | 0.02 | 0.02 | 0.03 | 0.02 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meyer, L.; Delgado-Cunningham, K.; Lorig-Roach, N.; Ford, J.; DuBois, R.M. Human Astrovirus 1–8 Seroprevalence Evaluation in a United States Adult Population. Viruses 2021, 13, 979. https://doi.org/10.3390/v13060979
Meyer L, Delgado-Cunningham K, Lorig-Roach N, Ford J, DuBois RM. Human Astrovirus 1–8 Seroprevalence Evaluation in a United States Adult Population. Viruses. 2021; 13(6):979. https://doi.org/10.3390/v13060979
Chicago/Turabian StyleMeyer, Lena, Kevin Delgado-Cunningham, Nicholas Lorig-Roach, Jordan Ford, and Rebecca M. DuBois. 2021. "Human Astrovirus 1–8 Seroprevalence Evaluation in a United States Adult Population" Viruses 13, no. 6: 979. https://doi.org/10.3390/v13060979
APA StyleMeyer, L., Delgado-Cunningham, K., Lorig-Roach, N., Ford, J., & DuBois, R. M. (2021). Human Astrovirus 1–8 Seroprevalence Evaluation in a United States Adult Population. Viruses, 13(6), 979. https://doi.org/10.3390/v13060979