
Changes in Cell-Mediated Immunity (IFN-γ and Granzyme B) Following Influenza Vaccination

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Vaccine
2.2. Antigens
2.3. IFN-γ and Granzyme B Assays
2.4. Antibody Titration
2.5. Statistical Analyses
3. Results
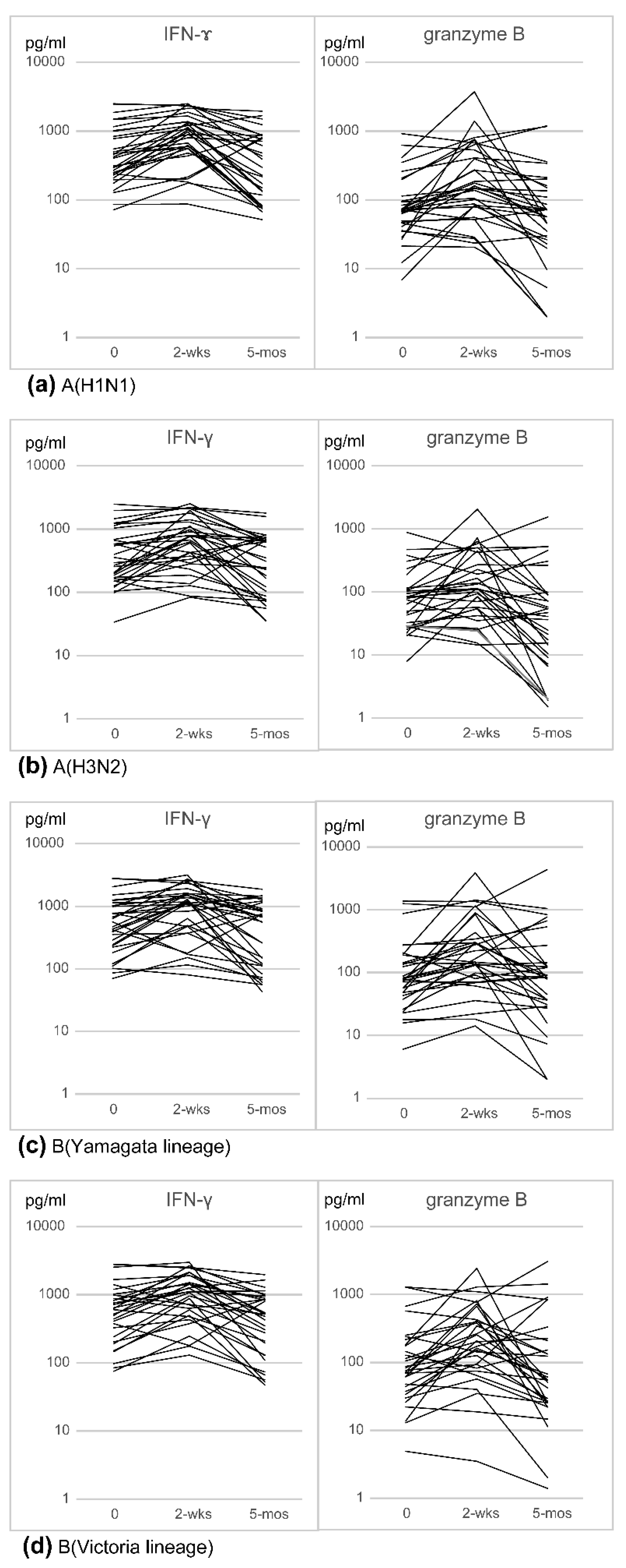
3.1. Changes in Cell-Mediated Immunity (IFN-γ, Granzyme B) after Influenza Vaccination
3.2. Correlation between IFN-γ and Granzyme B
3.3. Changes in Antibody Titers after Vaccination
- (a)
- Percentage of subjects with an HAI titer of ≥1:40.
- (b)
- Percentage of subjects with a pre-vaccine HAI titer of <1:10 and post-vaccine HAI titer of ≥1:40, or over 4-fold increase in HAI titer.
- (c)
- GMT ratio between the post-vaccination and pre-vaccination HAI GMT titers.
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Institute of Infectious Diseases (NIID). Influenza Vaccine Strain; NIID: Tokyo, Japan, (In Japanese). Available online: https://www.niid.go.jp/niid/ja/flu-m/flutoppage/2066-idsc/related/584-atpcs002.html (accessed on 31 May 2021).
- Available online: https://www.niid.go.jp/niid/images/iasr/2018/11/465tef01.gif (accessed on 31 May 2021).
- National Institute of Infectious Diseases (NIID). Influenza 2017/18 Season; NIID: Tokyo, Japan, 2018; Volume 39, pp. 181–183.
- National Institute of Infectious Diseases (NIID). Age Distribution of Cases with Isolation/Detection of Influenza Viruses; NIID: Tokyo, Japan, 2017.
- Garten, R.; Blanton, L.; Elal, A.I.A.; Alabi, N.; Barnes, J.; Biggerstaff, M.; Brammer, L.; Budd, A.P.; Burns, E.; Cummings, C.N.; et al. Update: Influenza Activity in the United States During the 2017–18 Season and Composition of the 2018–19 Influenza Vaccine. Morb. Mortal. Wkly. Rep. 2018, 67, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Centers for Diseases Control and Prevention (CDC). Seasonal Influenza Vaccine Effectiveness; CDC: Atlanta, GA, USA, 2017.
- Otani, N.; Shima, M.; Ueda, T.; Ichiki, K.; Nakajima, K.; Takesue, Y.; Okuno, T. Evaluation of Influenza Vaccine-Immunogenicity in Cell-Mediated Immunity. Cell. Immunol. 2016, 310, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Otani, N.; Shima, M.; Ueda, T.; Ichiki, K.; Nakajima, K.; Takesue, Y.; Honjo, K.; Yoshida, N.; Kawata, S.; Okuno, T. Relationship between the Frequency of Influenza Vaccination and Cell-Mediated Immunity. J. Immunol. Methods 2018, 458, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Otani, N.; Shima, M.; Ueda, T.; Ichiki, K.; Nakajima, K.; Takesue, Y.; Okuno, T. Evaluation of Influenza Vaccine-Induced Cell-Mediated Immunity: Comparison between Methods Using Peripheral Blood Mononuclear Cells and Whole Blood. Microbiol. Immunol. 2019, 63, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Abas, A.K.; Lichtman, A.H.; Pillai, S. Chapter 6: Effector Mechanisms of T Cell-Mediated Immunity. In Basic Immunology, 6th ed.; Elsevier: Philadelphia, PA, USA, 2019; pp. 119–136. [Google Scholar]
- McElhaney, J.E.; Ewen, C.; Zhou, X.; Kane, K.P.; Xie, D.; Hager, W.D.; Barry, M.B.; Kleppinger, A.; Wang, Y.; Bleackley, R.C. Granzyme B: Correlates with Protection and Enhanced CTL Response to Influenza Vaccination in Older Adults. Vaccine 2009, 27, 2418–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McElhaney, J.E.; Hooton, J.W.; Hooton, N.; Bleackley, R.C. Comparison of Single versus Booster Dose of Influenza Vaccination on Humoral and Cellular Immune Responses in Older Adults. Vaccine 2005, 23, 3294–3300. [Google Scholar] [CrossRef] [PubMed]
- Bresee, J.S.; Fly, A.M.; Sambhara, S.; Cox, N.J. Inactivated Influenza Vaccines. In Plotkin’s Vaccines, 7th ed.; Plotkin, S.A., Orenstein, W.A., Offit, P.A., Edwards, K.M., Eds.; Elsevier: Philadelphia, PA, USA, 2018; pp. 456–488. [Google Scholar]
- Merani, S.; Kuchel, G.A.; Kleppinger, A.; McElhaney, J.E. Influenza Vaccine-Mediated Protection in Older Adults: Impact of Influenza Infection, Cytomegalovirus Serostatus and Vaccine Dosage. Exp. Gerontol. 2018, 107, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Mbawuike, I.; Zang, Y.; Couch, R.B. Humoral and Cell-Mediated Immune Responses of Humans to Inactivated Influenza Vaccine with or without QS21 Adjuvant. Vaccine 2007, 25, 3263–3269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; McElhaney, J.E.; Walrond, L.; Cyr, T.D.; Merani, S.; Kollmann, T.R.; Halperin, S.A.; Scheifele, D.W. Cellular Immune Responses of Older Adults to Four Influenza Vaccines: Results of a Randomized, Controlled Comparison. Hum. Vaccin. Immunother. 2017, 13, 2048–2057. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, M.; Nakamura, A.; Arai, A.; Yamasaki, D.; Hirano, K.; Kobayashi, T.; Taguchi, O.; Kaneko, T.; Ito, M. The Direct Comparison of Two Interferon-gamma Release Assays in the Tuberculosis Screening of Japanese Healthcare Workers. Intern. Med. 2017, 56, 773–779. [Google Scholar] [CrossRef] [Green Version]
- Arvin, A. Aging, immunity, and the varicella-zoster virus. N. Engl. J. Med. 2005, 352, 2266–2267. [Google Scholar] [CrossRef] [PubMed]
- Sadaoka, K.; Okamoto, S.; Gomi, Y.; Tanimoto, T.; Ishikawa, T.; Yoshikawa, T.; Asano, Y.; Yamanishi, K.; Mori, Y. Measurement of varicella-zoster virus (VZV)-specific cell-mediated immunity: Comparison between VZV skin test and interferon-gamma enzyme-linked immunospot assay. J. Infect. Dis. 2008, 198, 1327–1333. [Google Scholar] [CrossRef]
- Otani, N.; Yamanishi, K.; Sakaguchi, Y.; Imai, Y.; Shima, M.; Okuno, T. Varicella-zoster virus-specific cell-mediated immunity in subjects with herpes zoster. J. Immunol. Methods. 2012, 377, 53–55. [Google Scholar] [CrossRef]
- Peters, W.; Brandl, J.R.; Lindbloom, J.D.; Martinez, C.J.; Scallan, C.D.; Trager, G.R.; Tingley, D.W.; Kabongo, M.L.; Tucker, S.N. Oral administration of an adenovirus vector encoding both an avian influenza A hemagglutinin and a TLR3 ligand induces antigen specific granzyme B and IFN-γ T cell responses in humans. Vaccine 2013, 31, 1752–1758. [Google Scholar] [CrossRef]
- European Agency for the Evaluation of Medicinal Products (EMEA). Committee for Proprietary Medicinal Products. Note for Guidance on Harmonization of Requirements for Influenza Vaccines. CPMP/BWP/214/96. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/note-guidance-harmonisation-requirements-influenza-vaccines_en.pdf (accessed on 31 May 2021).


{kind=link}
| (a) | ||||||||||
| n = 32 | IFN-γ: GMC (GMCFR) | Granzyme B: GMC (GMCFR) | ||||||||
| Pre-vaccine | 2 wks | 5 mos | Pre-vaccine | 2 wks | 5 mos | |||||
| H1N1 | 429.9 | 801.1 (1.9) | 307.5 (0.72) | 76.3 | 173.2 (2.3) | 53.9 (0.71) | ||||
| H3N2 | 344.8 | 597.4 (1.7) | 218.5 (0.6) | 68.9 | 122.7 (1.8) | 34.4 (0.5) | ||||
| Yamagata lineage | 490 | 835.1 (1.7) | 349 (0.7) | 87 | 196.8 (2.3) | 79.3 (0.9) | ||||
| Victoria lineage | 531.5 | 909.7 (1.7) | 350 (0.66) | 87.7 | 187 (2.1) | 66 (0.75) | ||||
| (b) | ||||||||||
| IFN-γ | Granzyme B | |||||||||
| n = 32 | 2 wks | 5 mos | 2 wks | 5 mos | ||||||
| H1N1 | 50% (16/32) | 25% (8/32) | 56.3% (18/32) | 28% (9/32) | ||||||
| H3N2 | 50% (16/32) | 22% (7/32) | 43.8% (14/32) | 16% (5/32) | ||||||
| Yamagata lineage | 50% (16/32) | 22% (7/32) | 62.5% (20/32) | 28% (9/32) | ||||||
| Victoria lineage | 62.5% (20/32) | 19% (6/32) | 65.6% (21/32) | 28% (9/32) | ||||||
| n = 32 | IFN-γ and Granzyme B | |||
|---|---|---|---|---|
| Pre-Vaccine | 2 wks Post-Vaccine | 5 mos Post-Vaccine | 2 wks Post-Vaccine/Pre-Vaccine | |
| H1N1 | 0.66 ** | 0.74 ** | 0.50 ** | 0.59 ** |
| H3N2 | 0.62 ** | 0.71 ** | 0.60 ** | 0.56 ** |
| Yamagata lineage | 0.74 ** | 0.80 ** | 0.77 ** | 0.47 ** |
| Victoria lineage | 0.75 ** | 0.75 ** | 0.63 ** | 0.61 ** |
| (a) | ||||||||
| Percentage of Subjects with HAI ≥ 1:40 | ||||||||
| n = 32 | Pre-Vaccine | 2 wks | 5 mos | |||||
| H1N1 | 15.6% (5/32) | 56.3% (18/32) | 43.8% (14/32) | |||||
| H3N2 | 68.8% (22/32) | 90.6% (29/32) | 68.8% (22/32) | |||||
| Yamagata lineage | 31.3% (10/32) | 53.1% (17/32) | 40.6% (13/32) | |||||
| Victoria lineage | 18.8% (6/32) | 34.4% (11/32) | 31.3% (10/32) | |||||
| (b) | ||||||||
| HAI | ||||||||
| n = 32 | 2 wks | 5 mos | ||||||
| H1N1 | 37.5% (12/32) | 21.9% (7/32) | ||||||
| H3N2 | 28.1% (9/32) | 12.5% (4/32) | ||||||
| Yamagata lineage | 15.6% (5/32) | 9.4% (3/32) | ||||||
| Victoria lineage | 15.6% (5/32) | 12.5% (4/32) | ||||||
| (c) | ||||||||
| n = 32 | GMT (GMT Ratio) | |||||||
| Pre-Vaccine | 2 wks Post-Vaccine | 5 mos Post-Vaccine | ||||||
| H1N1 | 10.9 | 28.9 (2.7) | 27.1 (2.5) | |||||
| H3N2 | 30.2 | 54.2 (1.8) | 43.6 (1.4) | |||||
| Yamagata lineage | 17.9 | 24.8 (1.4) | 22.8 (1.3) | |||||
| Victoria lineage | 12.2 | 17.2 (1.4) | 17.9 (1.5) | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otani, N.; Nakajima, K.; Ishikawa, K.; Ichiki, K.; Ueda, T.; Takesue, Y.; Yamamoto, T.; Tanimura, S.; Shima, M.; Okuno, T. Changes in Cell-Mediated Immunity (IFN-γ and Granzyme B) Following Influenza Vaccination. Viruses 2021, 13, 1137. https://doi.org/10.3390/v13061137
Otani N, Nakajima K, Ishikawa K, Ichiki K, Ueda T, Takesue Y, Yamamoto T, Tanimura S, Shima M, Okuno T. Changes in Cell-Mediated Immunity (IFN-γ and Granzyme B) Following Influenza Vaccination. Viruses. 2021; 13(6):1137. https://doi.org/10.3390/v13061137
Chicago/Turabian StyleOtani, Naruhito, Kazuhiko Nakajima, Kaori Ishikawa, Kaoru Ichiki, Takashi Ueda, Yoshio Takesue, Takuma Yamamoto, Susumu Tanimura, Masayuki Shima, and Toshiomi Okuno. 2021. "Changes in Cell-Mediated Immunity (IFN-γ and Granzyme B) Following Influenza Vaccination" Viruses 13, no. 6: 1137. https://doi.org/10.3390/v13061137
APA StyleOtani, N., Nakajima, K., Ishikawa, K., Ichiki, K., Ueda, T., Takesue, Y., Yamamoto, T., Tanimura, S., Shima, M., & Okuno, T. (2021). Changes in Cell-Mediated Immunity (IFN-γ and Granzyme B) Following Influenza Vaccination. Viruses, 13(6), 1137. https://doi.org/10.3390/v13061137