
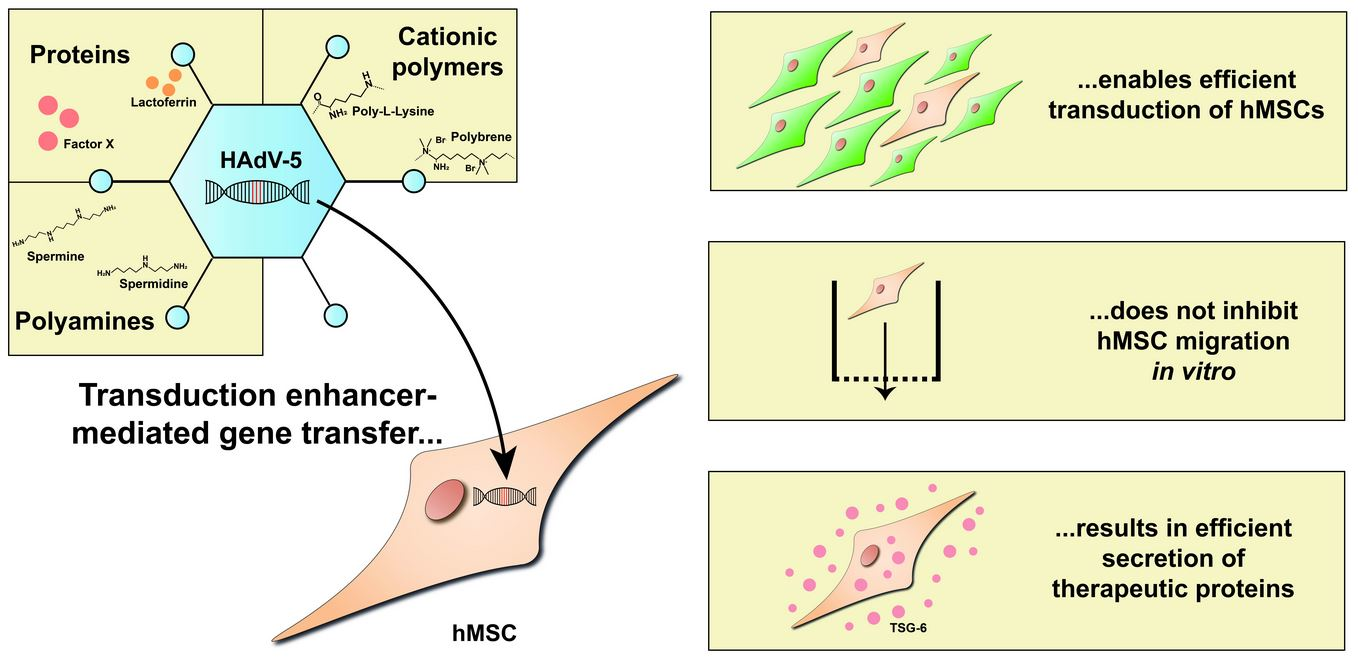
Transduction Enhancers Enable Efficient Human Adenovirus Type 5-Mediated Gene Transfer into Human Multipotent Mesenchymal Stromal Cells

 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Human MSC Isolation and Cell Culture
2.2. Adenoviral Vectors
2.3. Adenoviral Vector Production and Purification
2.4. Transduction of hMSCs by HAdV-5-eGFP with or without Transduction Enhancers
2.5. Boyden Chamber Assays Using BM-hMSCs
2.6. Isolation of Macrophages (MΦ)
2.7. Co-Cultivation of BM-hMSCs with Activated Macrophages
2.8. Transduction of BM-hMSCs with HAdV-5-TSG-6
2.9. Western Blot Analysis for Detection of TSG-6 in Cell Culture Supernatants
2.10. Statistical Analysis
3. Results
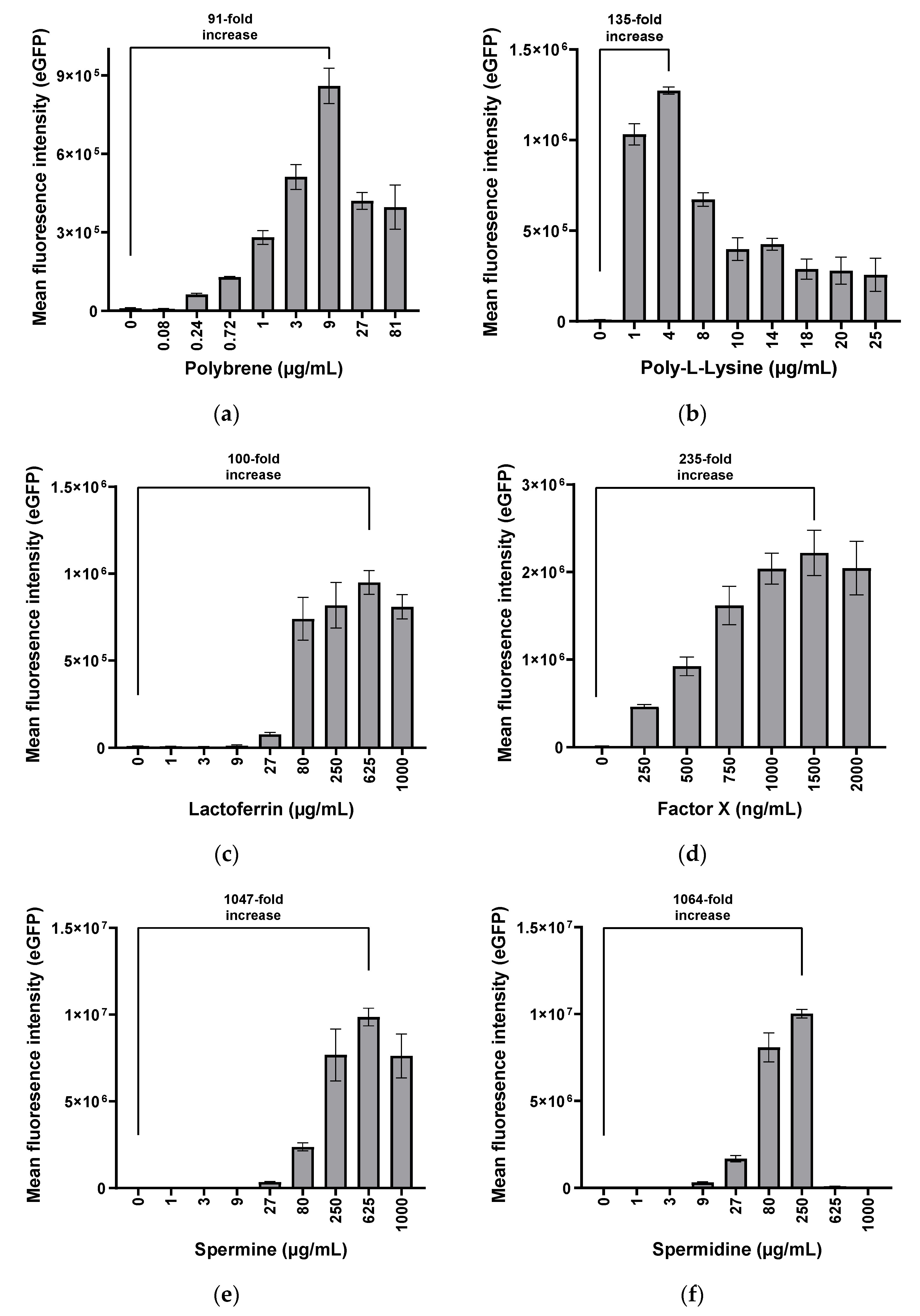
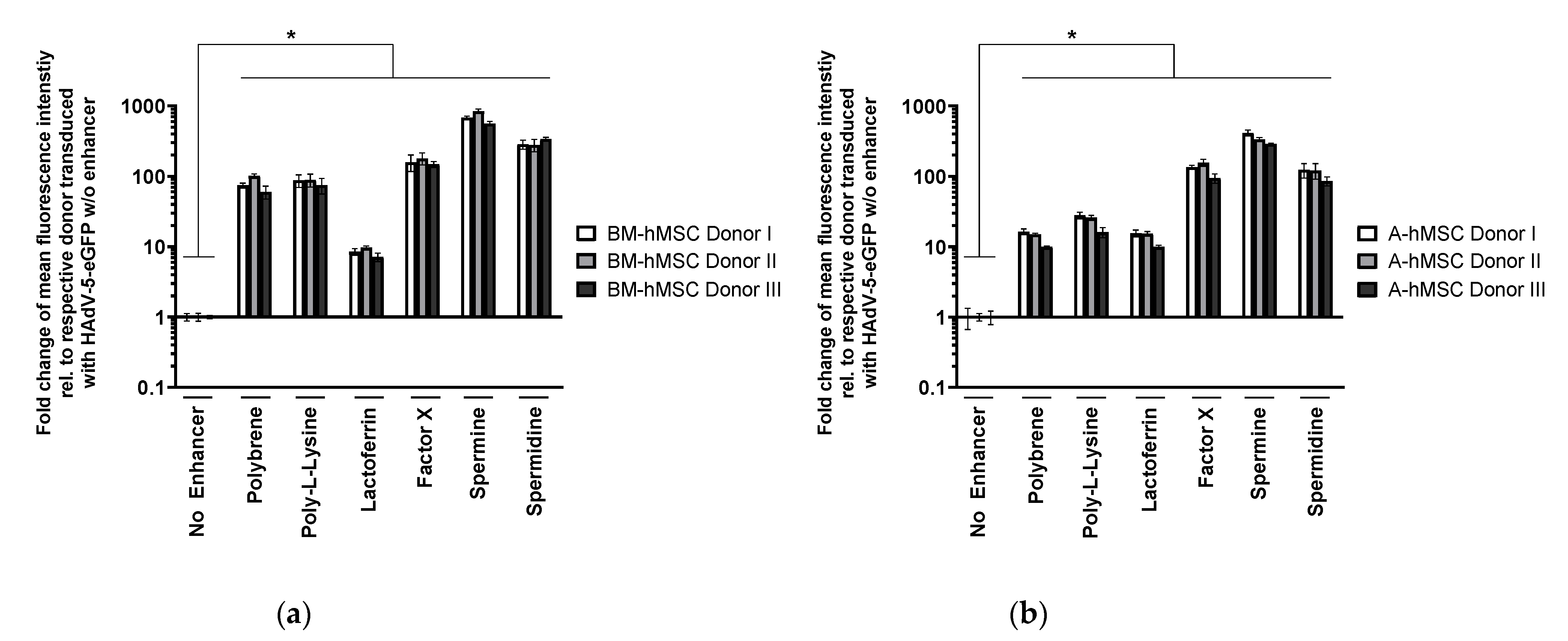
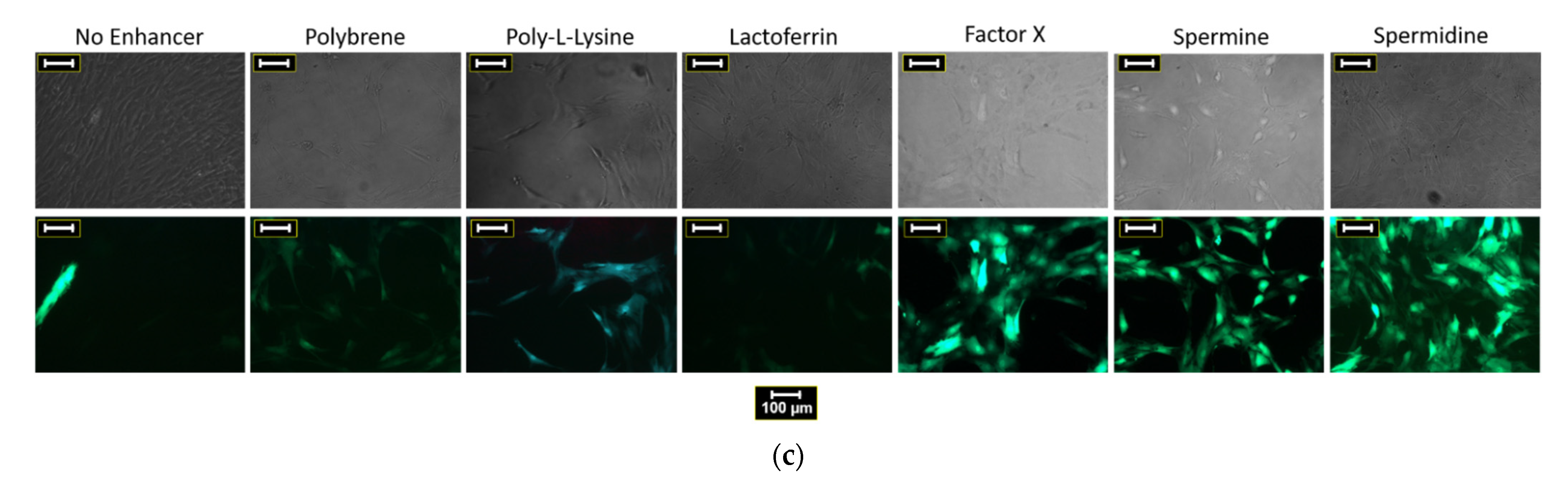
3.1. Polybrene, Poly-l-Lysine, Lactoferrin, Factor X, Spermine, and Spermidine Are Potent Enhancers of hMSC Transduction with HAdV-5 Vectors
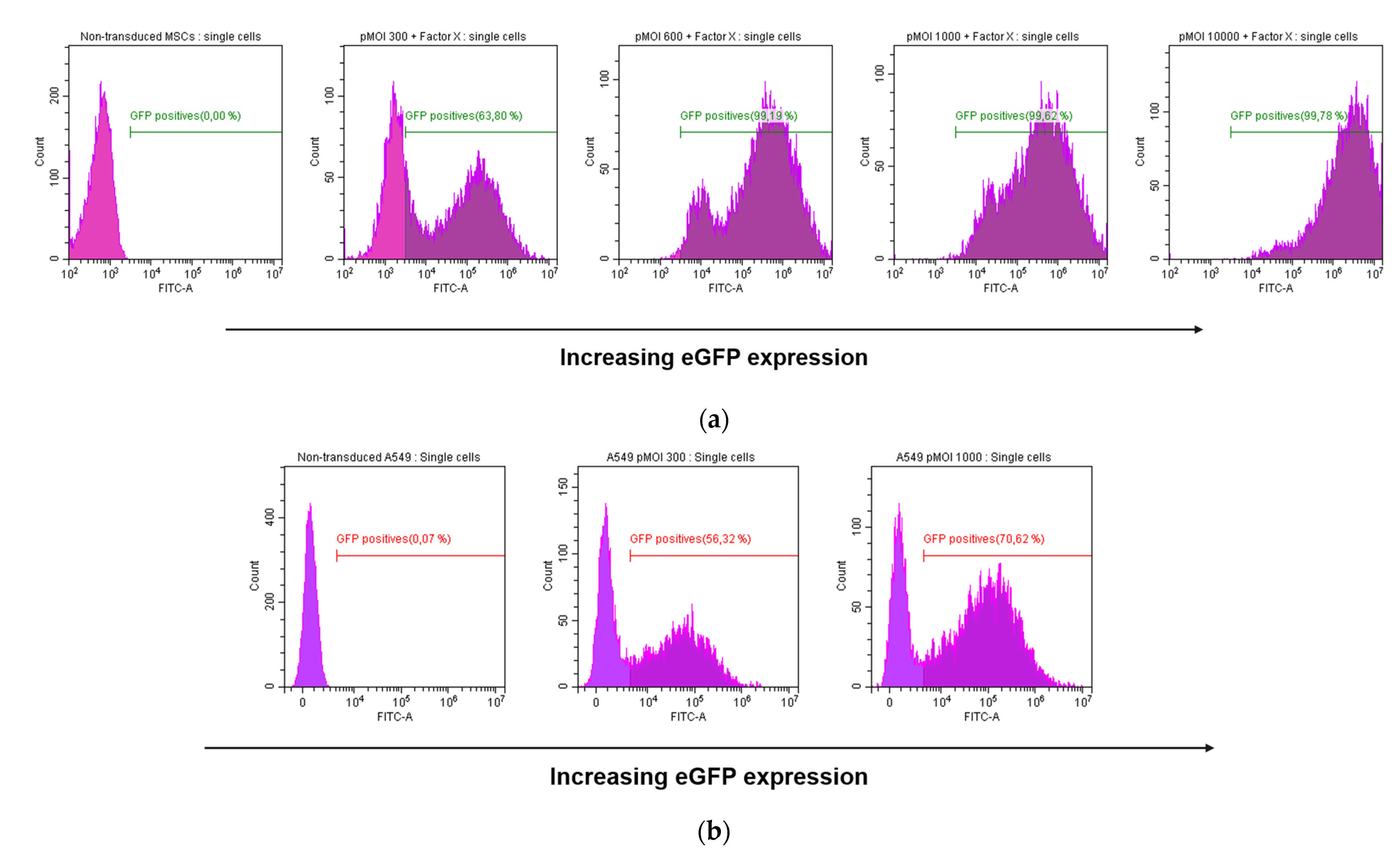
3.1.1. Determination of Optimal Transduction Enhancer Amounts
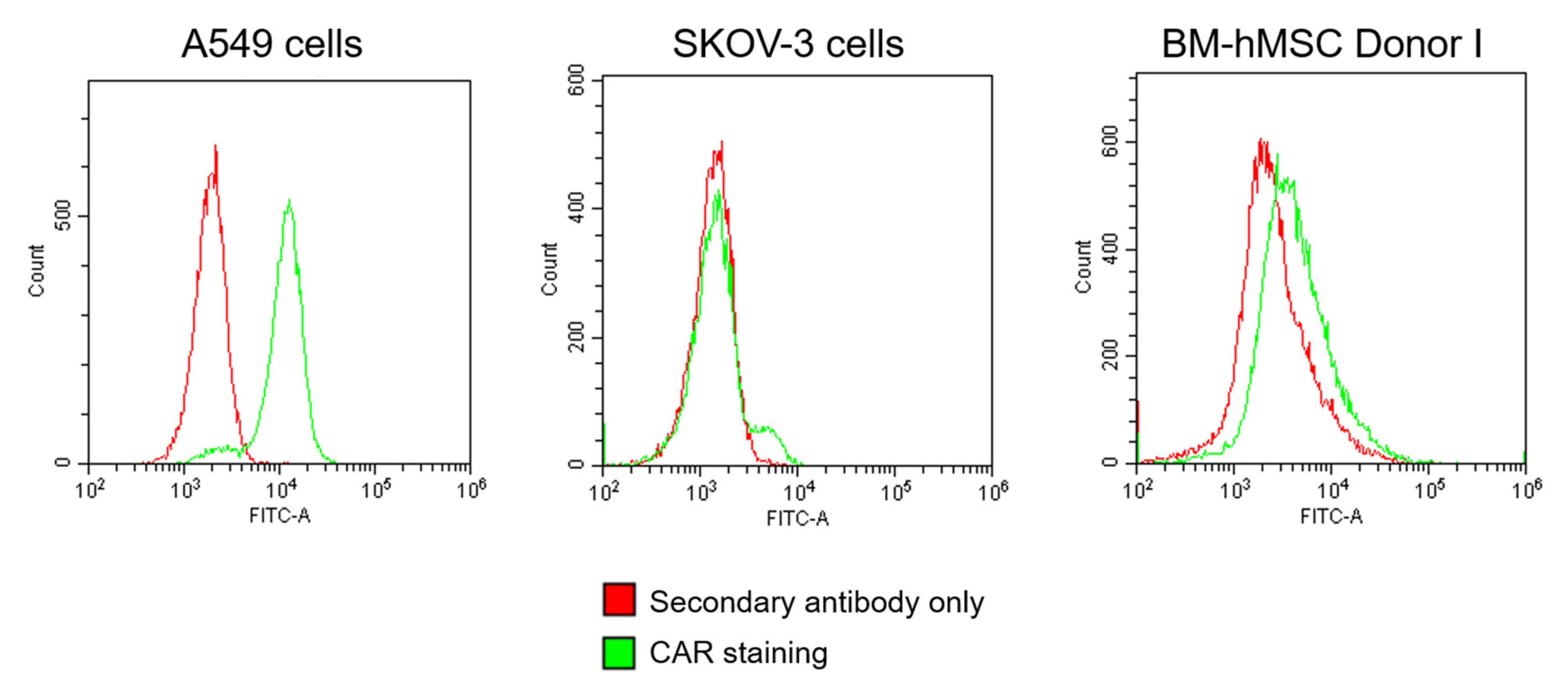
3.1.2. Validation of Transduction Enhancer Activity in BM-hMSCs and A-hMSCs of Different Donors
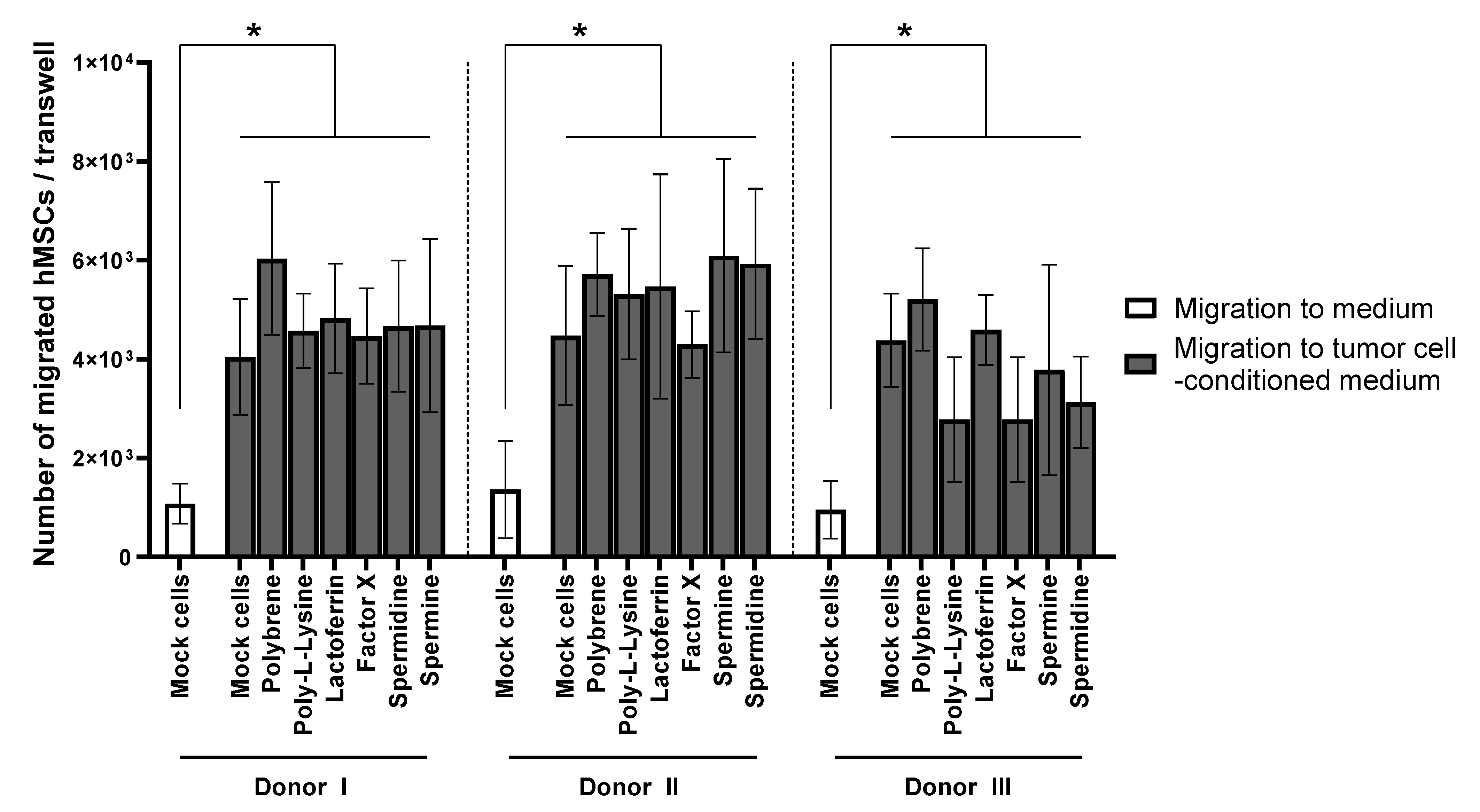
3.2. BM-hMSC Migration Is Not Inhibited by HAdV-5 Transduction or the Transduction Enhancers
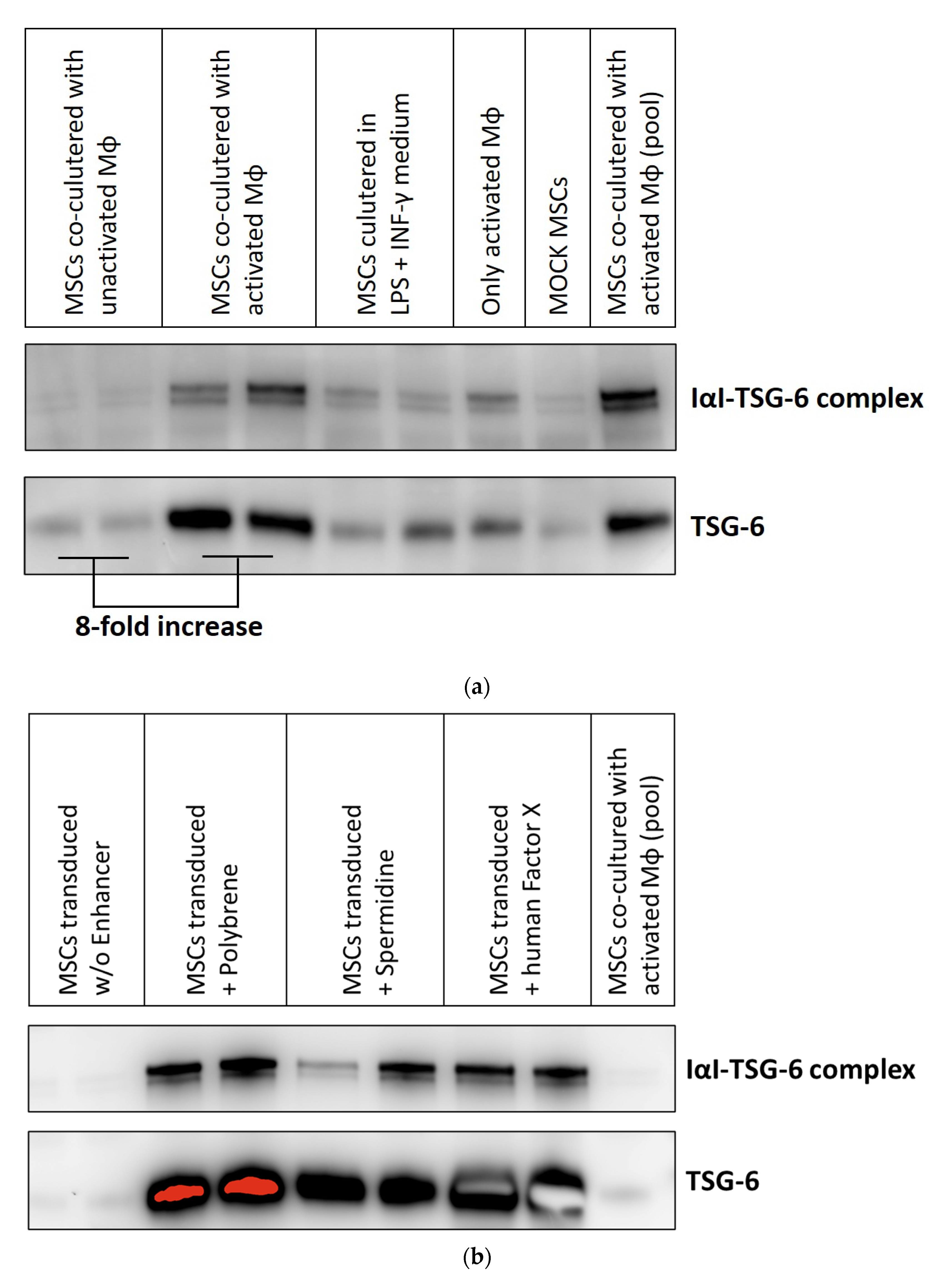
3.3. Transduction Enhancers Facilitate High-Level Expression of a Secreted Therapeutic Protein in BM-hMSC
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Dominici, M.; le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Giordano, A.; Galderisi, U.; Marino, I.R. From the Laboratory Bench to the Patient’s Bedside: An Update on Clinical Trials with Mesenchymal Stem Cells. J. Cell. Physiol. 2007, 211, 27–35. [Google Scholar] [CrossRef]
- Squillaro, T.; Peluso, G.; Galderisi, U. Clinical Trials with Mesenchymal Stem Cells: An Update. Cell Transplant. 2016, 25, 829–848. [Google Scholar] [CrossRef] [Green Version]
- Melen, G.J.; Franco-Luzón, L.; Ruano, D.; González-Murillo, Á.; Alfranca, A.; Casco, F.; Lassaletta, Á.; Alonso, M.; Madero, L.; Alemany, R.; et al. Influence of Carrier Cells on the Clinical Outcome of Children with Neuroblastoma Treated with High Dose of Oncolytic Adenovirus Delivered in Mesenchymal Stem Cells. Cancer Lett. 2016, 371, 161–170. [Google Scholar] [CrossRef]
- Volpers, C.; Kochanek, S. Adenoviral Vectors for Gene Transfer and Therapy. J. Gene Med. 2004, 6, S164–S171. [Google Scholar] [CrossRef] [PubMed]
- Breyer, B.; Jiang, W.; Cheng, H.; Zhou, L.; Paul, R.; Feng, T.; He, T.-C. Adenoviral Vector-Mediated Gene Transfer for Human Gene Therapy. Curr. Gene Ther. 2006, 1, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Lusky, M. Good Manufacturing Practice Production of Adenoviral Vectors for Clinical Trials. Hum. Gene Ther. 2005, 16, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Mathias, P.; Wickham, T.; Moore, M.; Nemerow, G. Multiple Adenovirus Serotypes Use Alpha v Integrins for Infection. J. Virol. 1994, 68, 6811–6814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, T.J.; Mathias, P.; Cheresh, D.A.; Nemerow, G.R. Integrins Avβ3 and Avβ5 Promote Adenovirus Internalization but Not Virus Attachment. Cell 1993, 73, 309–319. [Google Scholar] [CrossRef]
- Moreno, R.; Rojas, L.A.; Villellas, F.V.; Soriano, V.C.; García-Castro, J.; Fajardo, C.A.; Alemany, R. Human Menstrual Blood-Derived Mesenchymal Stem Cells as Potential Cell Carriers for Oncolytic Adenovirus. Stem Cells Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef]
- Semon, J.A.; Nagy, L.H.; Llamas, C.B.; Tucker, H.A.; Lee, R.H.; Prockop, D.J. Integrin Expression and Integrin-Mediated Adhesion in Vitro of Human Multipotent Stromal Cells (MSCs) to Endothelial Cells from Various Blood Vessels. Cell Tissue Res. 2010, 341, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Conget, P.A.; Minguell, J.J. Adenoviral-Mediated Gene Transfer into Ex Vivo Expanded Human Bone Marrow Mesenchymal Progenitor Cells. Exp. Hematol. 2000, 28, 382–390. [Google Scholar] [CrossRef]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a Common Receptor for Coxsackie B Viruses and Adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef]
- Sonabend, A.M.; Ulasov, I.V.; Tyler, M.A.; Rivera, A.A.; Mathis, J.M.; Lesniak, M.S. Mesenchymal Stem Cells Effectively Deliver an Oncolytic Adenovirus to Intracranial Glioma. Stem Cells 2008, 26, 831–841. [Google Scholar] [CrossRef] [Green Version]
- Hammer, K.; Kazcorowski, A.; Liu, L.; Behr, M.; Schemmer, P.; Herr, I.; Nettelbeck, D.M. Engineered Adenoviruses Combine Enhanced Oncolysis with Improved Virus Production by Mesenchymal Stromal Carrier Cells. Int. J. Cancer 2015, 137, 978–990. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wu, N.; Deng, F.; Zhang, H.; Wang, N.; Zhang, W.; Chen, X.; Wen, S.; Zhang, J.; Yin, L.; et al. Adenovirus-Mediated Gene Transfer in Mesenchymal Stem Cells Can Be Significantly Enhanced by the Cationic Polymer Polybrene. PLoS ONE 2014, 9, e92908. [Google Scholar] [CrossRef]
- Arcasoy, S.M.; Latoche, J.D.; Gondor, M.; Pitt, B.R.; Pilewski, J.M. Polycations Increase the Efficiency of Adenovirus-Mediated Gene Transfer to Epithelial and Endothelial Cells in Vitro. Gene Ther. 1997, 4, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Dumitrescu, M.; Vacaru, A.M.; Trusca, V.G.; Fenyo, I.M.; Ionita, R.; Gafencu, A.V. K2 Transfection System Boosts the Adenoviral Transduction of Murine Mesenchymal Stromal Cells. Int. J. Mol. Sci. 2021, 22, 598. [Google Scholar] [CrossRef]
- Fasbender, A.; Zabner, J.; Chillón, M.; Moninger, T.O.; Puga, A.P.; Davidson, B.L.; Welsh, M.J. Complexes of Adenovirus with Polycationic Polymers and Cationic Lipids Increase the Efficiency of Gene Transfer in Vitro and in Vivo. J. Biol. Chem. 1997, 272, 6479–6489. [Google Scholar] [CrossRef] [Green Version]
- Buo, A.M.; Williams, M.S.; Kerr, J.P.; Stains, J.P. A Cost-Effective Method to Enhance Adenoviral Transduction of Primary Murine Osteoblasts and Bone Marrow Stromal Cells. Bone Res. 2016, 4, 16021. [Google Scholar] [CrossRef] [Green Version]
- Johansson, C.; Jonsson, M.; Marttila, M.; Persson, D.; Fan, X.-L.; Skog, J.; Frängsmyr, L.; Wadell, G.; Arnberg, N. Adenoviruses Use Lactoferrin as a Bridge for CAR-Independent Binding to and Infection of Epithelial Cells. J. Virol. 2007, 81, 954–963. [Google Scholar] [CrossRef] [Green Version]
- Adams, W.C.; Bond, E.; Havenga, M.J.E.; Holterman, L.; Goudsmit, J.; Hedestam, G.B.K.; Koup, R.A.; Loré, K. Adenovirus Serotype 5 Infects Human Dendritic Cells via a Coxsackievirus-Adenovirus Receptor-Independent Receptor Pathway Mediated by Lactoferrin and DC-SIGN. J. Gen. Virol. 2009, 90, 1600–1610. [Google Scholar] [CrossRef]
- Zaiss, A.K.; Lawrence, R.; Elashoff, D.; Esko, J.D.; Herschman, H.R. Differential Effects of Murine and Human Factor X on Adenovirus Transduction via Cell-Surface Heparan Sulfate. J. Biol. Chem. 2011, 286, 24535–24543. [Google Scholar] [CrossRef] [Green Version]
- Alba, R.; Bradshaw, A.C.; Mestre-Francés, N.; Verdier, J.M.; Henaff, D.; Baker, A.H. Coagulation Factor X Mediates Adenovirus Type 5 Liver Gene Transfer in Non-Human Primates (Microcebus Murinus). Gene Ther. 2012, 19, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Fekete, N.; Rojewski, M.T.; Fürst, D.; Kreja, L.; Ignatius, A.; Dausend, J.; Schrezenmeier, H. GMP-Compliant Isolation and Large-Scale Expansion of Bone Marrow-Derived MSC. PLoS ONE 2012, 7, e43255. [Google Scholar] [CrossRef] [PubMed]
- Rojewski, M.T.; Lotfi, R.; Gjerde, C.; Mustafa, K.; Veronesi, E.; Ahmed, A.B.; Wiesneth, M.; Körper, S.; Sensebé, L.U.C.; Layrolle, P.; et al. Translation of a Standardized Manufacturing Protocol for Mesenchymal Stromal Cells: A Systematic Comparison of Validation and Manufacturing Data. Cytotherapy 2019, 21, 468–482. [Google Scholar] [CrossRef]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal Cells from the Adipose Tissue-Derived Stromal Vascular Fraction and Culture Expanded Adipose Tissue-Derived Stromal/Stem Cells: A Joint Statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy 2013, 15, 641–648. [Google Scholar] [CrossRef] [Green Version]
- Fekete, N.; Gadelorge, M.; Frst, D.; Maurer, C.; Dausend, J.; Fleury-Cappellesso, S.; Mailnder, V.; Lotfi, R.; Ignatius, A.; Sensebé, L.; et al. Platelet Lysate from Whole Blood-Derived Pooled Platelet Concentrates and Apheresis-Derived Platelet Concentrates for the Isolation and Expansion of Human Bone Marrow Mesenchymal Stromal Cells: Production Process, Content and Identification of Active Components. Cytotherapy 2012, 14, 540–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiedner, G.; Hertel, S.; Kochanek, S. Efficient Transformation of Primary Human Amniocytes by E1 Functions of Ad5: Generation of New Cell Lines for Adenoviral Vector Production. Hum. Gene Ther. 2000, 11, 2105–2116. [Google Scholar] [CrossRef]
- Mittereder, N.; March, K.L.; Trapnell, B.C. Evaluation of the Concentration and Bioactivity of Adenovirus Vectors for Gene Therapy. J. Virol. 1996, 70, 7498–7509. [Google Scholar] [CrossRef] [Green Version]
- Sindrilaru, A.; Peters, T.; Wieschalka, S.; Baican, C.; Baican, A.; Peter, H.; Hainzl, A.; Schatz, S.; Qi, Y.; Schlecht, A.; et al. An Unrestrained Proinflammatory M1 Macrophage Population Induced by Iron Impairs Wound Healing in Humans and Mice. J. Clin. Investig. 2011, 121, 985–997. [Google Scholar] [CrossRef] [Green Version]
- Wisniewski, H.G.; Vilček, J. TSG-6: An IL-1/TNF-Inducible Protein with Anti-Inflammatory Activity. Cytokine Growth Factor Rev. 1997, 8, 143–156. [Google Scholar] [CrossRef]
- Milner, C.M.; Day, A.J. TSG-6: A Multifunctional Protein Associated with Inflammation. J. Cell Sci. 2003, 116, 1863–1873. [Google Scholar] [CrossRef] [Green Version]
- Day, A.J.; Milner, C.M. TSG-6: A Multifunctional Protein with Anti-Inflammatory and Tissue-Protective Properties. Matrix Biol. 2019, 78–79, 60–83. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Jiang, D.; Sindrilaru, A.; Stegemann, A.; Schatz, S.; Treiber, N.; Rojewski, M.; Schrezenmeier, H.; vander Beken, S.; Wlaschek, M.; et al. TSG-6 Released from Intradermally Injected Mesenchymal Stem Cells Accelerates Wound Healing and Reduces Tissue Fibrosis in Murine Full-Thickness Skin Wounds. J. Investig. Dermatol. 2014, 134, 526–537. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.H.; Wisniewski, H.G.; Vilcek, J. A Novel Secretory Tumor Necrosis Factor-Inducible Protein (TSG-6) Is a Member of the Family of Hyaluronate Binding Proteins, Closely Related to the Adhesion Receptor CD44. J. Cell Biol. 1992, 116, 545–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisniewski, H.G.; Oppenheim, J.D.; Vilček, J.; Burgess, W.H. TSG-6, an Arthritis-Associated Hyaluronan Binding Protein, Forms a Stable Complex with the Serum Protein Inter-α-Inhibitor. Biochemistry 1994, 33, 7423–7429. [Google Scholar] [CrossRef]
- Rugg, M.S.; Willis, A.C.; Mukhopadhyay, D.; Hascall, V.C.; Fries, E.; Fülöp, C.; Milner, C.M.; Day, A.J. Characterization of Complexes Formed between TSG-6 and Inter-α- Inhibitor That Act as Intermediates in the Covalent Transfer of Heavy Chains onto Hyaluronan. J. Biol. Chem. 2005, 280, 25674–25686. [Google Scholar] [CrossRef] [Green Version]
- Saeed, H.; Ahsan, M.; Saleem, Z.; Iqtedar, M.; Islam, M.; Danish, Z.; Khan, A.M. Mesenchymal Stem Cells (MSCs) as Skeletal Therapeutics-an Update. J. Biomed. Sci. 2016, 23, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Hare, J.M.; Traverse, J.H.; Henry, T.D.; Dib, N.; Strumpf, R.K.; Schulman, S.P.; Gerstenblith, G.; DeMaria, A.N.; Denktas, A.E.; Gammon, R.S.; et al. A Randomized, Double-Blind, Placebo-Controlled, Dose-Escalation Study of Intravenous Adult Human Mesenchymal Stem Cells (Prochymal) After Acute Myocardial Infarction. J. Am. Coll. Cardiol. 2009, 54, 2277–2286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- le Blanc, K.; Frassoni, F.; Ball, L.; Locatelli, F.; Roelofs, H.; Lewis, I.; Lanino, E.; Sundberg, B.; Bernardo, M.E.; Remberger, M.; et al. Mesenchymal Stem Cells for Treatment of Steroid-Resistant, Severe, Acute Graft-versus-Host Disease: A Phase II Study. Lancet 2008, 371, 1579–1586. [Google Scholar] [CrossRef]
- Cheng, Z.; Ou, L.; Zhou, X.; Li, F.; Jia, X.; Zhang, Y.; Liu, X.; Li, Y.; Ward, C.A.; Melo, L.G.; et al. Targeted Migration of Mesenchymal Stem Cells Modified with CXCR4 Gene to Infarcted Myocardium Improves Cardiac Performance. Mol. Ther. 2008, 16, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Moon, H.H.; Kim, H.A.; Hwang, K.C.; Lee, M.; Choi, D. Hypoxia-Inducible Vascular Endothelial Growth Factor-Engineered Mesenchymal Stem Cells Prevent Myocardial Ischemic Injury. Mol. Ther. 2011, 19, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Vural, A.C.; Odabas, S.; Korkusuz, P.; Yar Sağlam, A.S.; Bilgiç, E.; Çavuşoğlu, T.; Piskin, E.; Vargel, İ. Cranial Bone Regeneration via BMP-2 Encoding Mesenchymal Stem Cells. Artif. Cells Nanomed. Biotechnol. 2017, 45, 544–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, M.; García-Castro, J.; Melen, G.J.; González-Murillo, Á.; Franco-Luzón, L. Patient-Derived Mesenchymal Stem Cells as Delivery Vehicles for Oncolytic Virotherapy: Novel State-of-the-Art Technology. Oncolytic Virother. 2015, 4, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.; Zhou, N.; Wan, L.; Su, X.; Sun, Z.; Mizuguchi, H.; Yoshioka, Y.; Nakagawa, S.; Zhao, R.C.; Gao, J.Q. Polyethyleneimine-Coating Enhances Adenoviral Transduction of Mesenchymal Stem Cells. Biochem. Biophys. Res. Commun. 2014, 447, 383–387. [Google Scholar] [CrossRef]
- Clark, P.R.; Stopeck, A.T.; Brailey, J.L.; Wang, Q.; Mcarthur, J.; Finer, M.H.; Hersh, E.M. Polycations and Cationic Lipids Enhance Adenovirus Transduction and Transgene Expression in Tumor Cells. Cancer Gene Ther. 1999, 6, 437–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- di Nicola, M.; Milanesi, M.; Magni, M.; Bregni, M.; Carlo-Stella, C.; Longoni, P.; Tomanin, R.; Ravagnani, F.; Scarpa, M.; Jordan, C.; et al. Recombinant Adenoviral Vector-LipofectAMINE Complex for Gene Transduction into Human T Lymphocytes. Hum. Gene Ther. 1999, 10, 1875–1884. [Google Scholar] [CrossRef]
- Byk, T.; Haddada, H.; Vainchenker, W.; Louache, F. Lipofectamine and Related Cationic Lipids Strongly Improve Adenoviral Infection Efficiency of Primitive Human Hematopoietic Cells. Hum. Gene Ther. 1998, 9, 2493–2502. [Google Scholar] [CrossRef]
- Handa, A.K.; Fatima, T.; Mattoo, A.K. Polyamines: Bio-Molecules with Diverse Functions in Plant and Human Health and Disease. Front. Chem. 2018, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firpo, M.R.; Mounce, B.C. Diverse Functions of Polyamines in Virus Infection. Biomolecules 2020, 10, 628. [Google Scholar] [CrossRef] [Green Version]
- Gibson, W.; Roizman, B. Compartmentalization of Spermine and Spermidine in the Herpes Simplex Virion. Proc. Natl. Acad. Sci. USA 1971, 68, 2818–2821. [Google Scholar] [CrossRef] [Green Version]
- Olsen, M.E.; Filone, C.M.; Rozelle, D.; Mire, C.E.; Agans, K.N.; Hensley, L.; Connor, J.H. Polyamines and Hypusination Are Required for Ebolavirus Gene Expression and Replication. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kicmal, T.M.; Tate, P.M.; Dial, C.N.; Esin, J.J.; Mounce, B.C. Polyamine Depletion Abrogates Enterovirus Cellular Attachment. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Hegre, O.D.; Marshall, S.; Hickey, G.E. Spermidine cytotoxicity in vitro: Effect of serum and oxygen tension. In Vitro 1984, 20, 198–204. [Google Scholar] [CrossRef]
- Sharmin, S.; Sakata, K.; Kashiwagi, K.; Ueda, S.; Iwasaki, S.; Shirahata, A.; Igarashi, K. Polyamine Cytotoxicity in the Presence of Bovine Serum Amine Oxidase. Biochem. Biophys. Res. Commun. 2001, 282, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Averill-Bates, D.A.; Ke, Q.; Tanel, A.; Roy, J.; Fortier, G.; Agostinelli, E. Mechanism of Cell Death Induced by Spermine and Amine Oxidase in Mouse Melanoma Cells. Int. J. Oncol. 2008, 32, 79–88. [Google Scholar] [CrossRef]
- Alba, R.; Bradshaw, A.C.; Parker, A.L.; Bhella, D.; Waddington, S.N.; Nicklin, S.A.; van Rooijen, N.; Custers, J.; Goudsmit, J.; Barouch, D.H.; et al. Undefined Identification of Coagulation Factor (F) X Binding Sites on the Adenovirus Serotype 5 Hexon: Effect of Mutagenesis on FX Interactions and Gene Transfer. J. Am. Soc. Hematol. 2009, 114, 965–971. [Google Scholar]
- Alba, R.; Bradshaw, A.C.; Coughlan, L.; Denby, L.; McDonald, R.A.; Waddington, S.N.; Buckley, S.M.K.; Greig, J.A.; Parker, A.L.; Miller, A.M.; et al. Biodistribution and Retargeting of FX-Binding Ablated Adenovirus Serotype 5 Vectors. Blood 2010, 116, 2656–2664. [Google Scholar] [CrossRef] [Green Version]
- Shayakhmetov, D.M.; Gaggar, A.; Ni, S.; Li, Z.-Y.; Lieber, A. Adenovirus Binding to Blood Factors Results in Liver Cell Infection and Hepatotoxicity. J. Virol. 2005, 79, 7478–7491. [Google Scholar] [CrossRef] [Green Version]
- Kalyuzhniy, O.; di Paolo, N.C.; Silvestry, M.; Hofherr, S.E.; Barry, M.A.; Stewart, P.L.; Shayakhmetov, D.M. Adenovirus Serotype 5 Hexon Is Critical for Virus Infection of Hepatocytes in Vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 5483–5488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradshaw, A.C.; Parker, A.L.; Duffy, M.R.; Coughlan, L.; van Rooijen, N.; Kähäri, V.M.; Nicklin, S.A.; Baker, A.H. Requirements for Receptor Engagement during Infection by Adenovirus Complexed with Blood Coagulation Factor X. PLoS Pathog. 2010, 6, e1001142. [Google Scholar] [CrossRef] [Green Version]
- Spaeth, E.; Klopp, A.; Dembinski, J.; Andreeff, M.; Marini, F. Inflammation and Tumor Microenvironments: Defining the Migratory Itinerary of Mesenchymal Stem Cells. Gene Ther. 2008. [Google Scholar] [CrossRef] [PubMed]
- Schichor, C.; Birnbaum, T.; Etminan, N.; Schnell, O.; Grau, S.; Miebach, S.; Aboody, K.; Padovan, C.; Straube, A.; Tonn, J.C.; et al. Vascular Endothelial Growth Factor A Contributes to Glioma-Induced Migration of Human Marrow Stromal Cells (HMSC). Exp. Neurol. 2006, 199, 301–310. [Google Scholar] [CrossRef]
- Gao, H.; Priebe, W.; Glod, J.; Banerjee, D. Activation of Signal Transducers and Activators of Transcription 3 and Focal Adhesion Kinase by Stromal Cell-Derived Factor 1 Is Required for Migration of Human Mesenchymal Stem Cells in Response to Tumor Cell-Conditioned Medium. Stem Cells 2009, 27, 857–865. [Google Scholar] [CrossRef]
- Zhu, H.; Mitsuhashi, N.; Klein, A.; Barsky, L.W.; Weinberg, K.; Barr, M.L.; Demetriou, A.; Wu, G.D. The Role of the Hyaluronan Receptor CD44 in Mesenchymal Stem Cell Migration in the Extracellular Matrix. Stem Cells 2006, 24, 928–935. [Google Scholar] [CrossRef]
- Watts, T.L.; Cui, R.; Szaniszlo, P.; Resto, V.A.; Powell, D.W.; Pinchuk, I.V. PDGF-AA Mediates Mesenchymal Stromal Cell Chemotaxis to the Head and Neck Squamous Cell Carcinoma Tumor Microenvironment. J. Transl. Med. 2016, 14, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ponte, A.L.; Marais, E.; Gallay, N.; Langonné, A.; Delorme, B.; Hérault, O.; Charbord, P.; Domenech, J. The In Vitro Migration Capacity of Human Bone Marrow Mesenchymal Stem Cells: Comparison of Chemokine and Growth Factor Chemotactic Activities. Stem Cells 2007, 25, 1737–1745. [Google Scholar] [CrossRef]
- Milner, C.M.; Higman, V.A.; Day, A.J. TSG-6: A Pluripotent Inflammatory Mediator? Biochem. Soc. Trans. 2006, 34, 446–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Chen, L.; Scott, P.G.; Tredget, E.E. Mesenchymal Stem Cells Enhance Wound Healing Through Differentiation and Angiogenesis. Stem Cells 2007, 25, 2648–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maharlooei, M.K.; Bagheri, M.; Solhjou, Z.; Jahromi, B.M.; Akrami, M.; Rohani, L.; Monabati, A.; Noorafshan, A.; Omrani, G.R. Adipose Tissue Derived Mesenchymal Stem Cell (AD-MSC) Promotes Skin Wound Healing in Diabetic Rats. Diabetes Res. Clin. Pract. 2011, 93, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Di, G.; Du, X.; Qi, X.; Zhao, X.; Duan, H.; Li, S.; Xie, L.; Zhou, Q. Mesenchymal Stem Cells Promote Diabetic Corneal Epithelial Wound Healing through TSG-6-Dependent Stem Cell Activation and Macrophage Switch. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4064–4074. [Google Scholar] [CrossRef] [PubMed]
- Meinel, L.; Hofmann, S.; Betz, O.; Fajardo, R.; Merkle, H.P.; Langer, R.; Evans, C.H.; Vunjak-Novakovic, G.; Kaplan, D.L. Osteogenesis by Human Mesenchymal Stem Cells Cultured on Silk Biomaterials: Comparison of Adenovirus Mediated Gene Transfer and Protein Delivery of BMP-2. Biomaterials 2006, 27, 4993–5002. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transduction Enhancer | Total Amount | Concentration during Pre-Incubation |
|---|---|---|
| Polybrene | 18 fg/viral particle | 9 µg/mL |
| Poly-l-Lysine | 4% (v/v) * | 4% (v/v) * |
| Human Lactoferrin | 1250 fg/viral particle | 625 µg/mL |
| Human Factor X | 4 fg/viral particle | 1500 ng/mL |
| Spermine | 1250 fg/viral particle | 625 µg/mL |
| Spermidine | 500 fg/viral particle | 250 µg/mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nilson, R.; Lübbers, O.; Weiß, L.; Singh, K.; Scharffetter-Kochanek, K.; Rojewski, M.; Schrezenmeier, H.; Zeplin, P.H.; Funk, W.; Krutzke, L.; et al. Transduction Enhancers Enable Efficient Human Adenovirus Type 5-Mediated Gene Transfer into Human Multipotent Mesenchymal Stromal Cells. Viruses 2021, 13, 1136. https://doi.org/10.3390/v13061136
Nilson R, Lübbers O, Weiß L, Singh K, Scharffetter-Kochanek K, Rojewski M, Schrezenmeier H, Zeplin PH, Funk W, Krutzke L, et al. Transduction Enhancers Enable Efficient Human Adenovirus Type 5-Mediated Gene Transfer into Human Multipotent Mesenchymal Stromal Cells. Viruses. 2021; 13(6):1136. https://doi.org/10.3390/v13061136
Chicago/Turabian StyleNilson, Robin, Olivia Lübbers, Linus Weiß, Karmveer Singh, Karin Scharffetter-Kochanek, Markus Rojewski, Hubert Schrezenmeier, Philip Helge Zeplin, Wolfgang Funk, Lea Krutzke, and et al. 2021. "Transduction Enhancers Enable Efficient Human Adenovirus Type 5-Mediated Gene Transfer into Human Multipotent Mesenchymal Stromal Cells" Viruses 13, no. 6: 1136. https://doi.org/10.3390/v13061136
APA StyleNilson, R., Lübbers, O., Weiß, L., Singh, K., Scharffetter-Kochanek, K., Rojewski, M., Schrezenmeier, H., Zeplin, P. H., Funk, W., Krutzke, L., Kochanek, S., & Kritzinger, A. (2021). Transduction Enhancers Enable Efficient Human Adenovirus Type 5-Mediated Gene Transfer into Human Multipotent Mesenchymal Stromal Cells. Viruses, 13(6), 1136. https://doi.org/10.3390/v13061136