
Combinatorial Approaches for Cancer Treatment Using Oncolytic Viruses: Projecting the Perspectives through Clinical Trials Outcomes

 , and
, and
Abstract
:1. Introduction
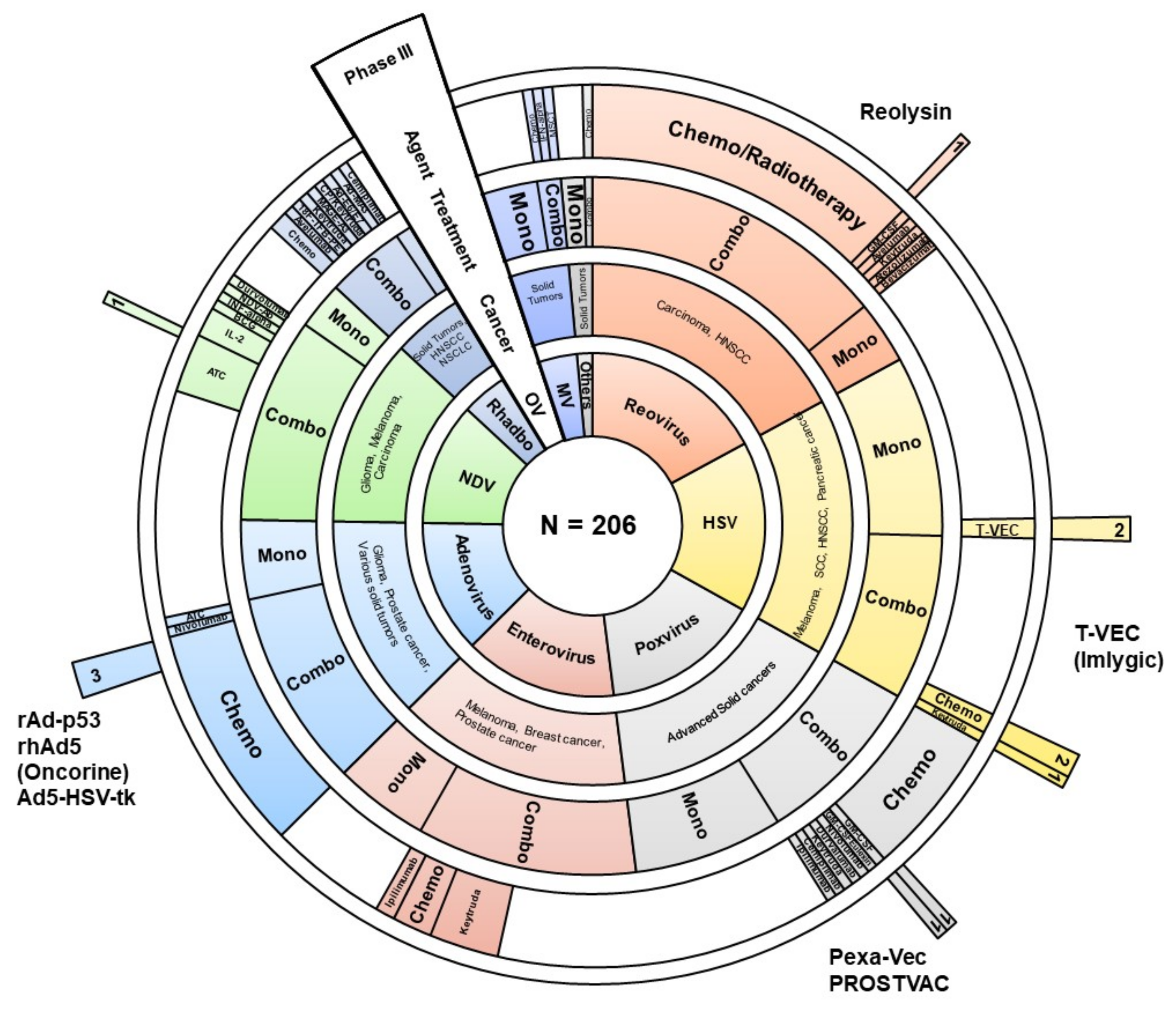
2. Oncolytic Viruses in Clinical Trials
2.1. Adenoviruses
2.2. Herpesviruses
2.3. Poxviruses
2.4. Enteroviruses (Picornaviridae)
2.5. Paramyxoviruses
2.6. Rhabdoviruses
2.7. Parvoviruses
2.8. Reoviruses
3. Combination Therapies and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Williams, J. The Immunotherapy Revolution: The Best New Hope for Saving Cancer Patients; Williams Cancer Institute: Ciudad de Mexico, Mexico, 2019; ISBN 9781700114938. [Google Scholar]
- Farkona, S.; Diamandis, E.P.; Blasutig, I.M. Cancer immunotherapy: The beginning of the end of cancer? BMC Med. 2016, 14, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Liberal, J.; Ochoa de Olza, M.; Hierro, C.; Gros, A.; Rodon, J.; Tabernero, J. The expanding role of immunotherapy. Cancer Treat. Rev. 2017, 54, 74–86. [Google Scholar] [CrossRef]
- Galluzzi, L.; Rudqvist, N.-P. Preface: More than two decades of modern tumor immunology. Methods Enzymol. 2020, 631, xxiii–xlii. [Google Scholar]
- Vile, R.G. The Immune System in Oncolytic Immunovirotherapy: Gospel, Schism and Heresy. Mol. Ther. 2018, 26, 942–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Overwijk, W.W. Intratumoral immunotherapy for melanoma. Cancer Immunol. Immunother. 2015, 64, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Boisgerault, N.; Tangy, F.; Gregoire, M. New perspectives in cancer virotherapy: Bringing the immune system into play. Immunotherapy 2010, 2, 185–199. [Google Scholar] [CrossRef] [PubMed]
- De Gruijl, T.D.; Janssen, A.B.; van Beusechem, V.W. Arming oncolytic viruses to leverage antitumor immunity. Expert Opin. Biol. Ther. 2015, 15, 959–971. [Google Scholar] [CrossRef]
- Puduvalli, V.K. Vaccine Therapies Against Gliomas: Prime Time Yet? Oncology 2016, 30, 222–223. [Google Scholar]
- Ahmad, A.; Uddin, S.; Steinhoff, M. CAR-T Cell Therapies: An Overview of Clinical Studies Supporting Their Approved Use against Acute Lymphoblastic Leukemia and Large B-Cell Lymphomas. Int. J. Mol. Sci. 2020, 21, 3906. [Google Scholar] [CrossRef]
- Hoos, A. Development of immuno-oncology drugs-from CTLA4 to PD1 to the next generations. Nat. Rev. Drug Discov. 2016, 15, 235–247. [Google Scholar] [CrossRef]
- Xu, C.; Li, H.; Su, C.; Li, Z. Viral therapy for pancreatic cancer: Tackle the bad guys with poison. Cancer Lett. 2013, 333, 1–8. [Google Scholar] [CrossRef]
- Dyer, A.; Baugh, R.; Chia, S.L.; Frost, S.; Iris Jacobus, E.J.; Khalique, H.; Pokrovska, T.D.; Scott, E.M.; Taverner, W.K. Turning cold tumours hot: Oncolytic virotherapy gets up close and personal with other therapeutics at the 11th Oncolytic Virus Conference. Cancer Gene Ther. 2019, 26, 59–73. [Google Scholar] [CrossRef]
- Breitbach, C.J.; De Silva, N.S.; Falls, T.J.; Aladl, U.; Evgin, L.; Paterson, J.; Sun, Y.Y.; Roy, D.G.; Rintoul, J.L.; Daneshmand, M.; et al. Targeting tumor vasculature with an oncolytic virus. Mol. Ther. 2011, 19, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Burke, J.; Nieva, J.; Borad, M.J.; Breitbach, C.J. Oncolytic viruses: Perspectives on clinical development. Curr. Opin. Virol. 2015, 13, 55–60. [Google Scholar] [CrossRef]
- Eissa, I.R.; Bustos-Villalobos, I.; Ichinose, T.; Matsumura, S.; Naoe, Y.; Miyajima, N.; Morimoto, D.; Mukoyama, N.; Zhiwen, W.; Tanaka, M.; et al. The Current Status and Future Prospects of Oncolytic Viruses in Clinical Trials against Melanoma, Glioma, Pancreatic, and Breast Cancers. Cancers 2018, 10, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamid, O.; Ismail, R.; Puzanov, I. Intratumoral Immunotherapy-Update 2019. Oncologist 2020, 25, e423–e438. [Google Scholar] [CrossRef] [PubMed]
- Macedo, N.; Miller, D.M.; Haq, R.; Kaufman, H.L. Clinical landscape of oncolytic virus research in 2020. J. Immunother. Cancer 2020, 8, e001486. [Google Scholar] [CrossRef]
- Cook, M.; Chauhan, A. Clinical application of oncolytic viruses: A systematic review. Int. J. Mol. Sci. 2020, 21, 7505. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Guo, F. Recent updates on cancer immunotherapy. Precis. Clin. Med. 2018, 1, 65–74. [Google Scholar] [CrossRef]
- Bommareddy, P.K.; Kaufman, H.L. Unleashing the therapeutic potential of oncolytic viruses. J. Clin. Investig. 2018, 128, 1258–1260. [Google Scholar] [CrossRef] [PubMed]
- Fasullo, M.; Burch, A.D.; Britton, A. Hypoxia enhances the replication of oncolytic herpes simplex virus in p53- breast cancer cells. Cell Cycle 2009, 8, 2194–2197. [Google Scholar] [CrossRef] [Green Version]
- Zamarin, D.; Palese, P. Oncolytic Newcastle disease virus for cancer therapy: Old challenges and new directions. Future Microbiol. 2012, 7, 347–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Vitale, I.; Warren, S.; Adjemian, S.; Agostinis, P.; Martinez, A.B.; Chan, T.A.; Coukos, G.; Demaria, S.; Deutsch, E.; et al. Consensus guidelines for the definition, detection and interpretation of immunogenic cell death. J. Immunother. Cancer 2020, 8, e000337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christian, S.L.; Collier, T.W.; Zu, D.; Licursi, M.; Hough, C.M.; Hirasawa, K. Activated Ras/MEK Inhibits the Antiviral Response of Alpha Interferon by Reducing STAT2 Levels. J. Virol. 2009, 83, 6717–6726. [Google Scholar] [CrossRef] [Green Version]
- Seegers, S.L.; Frasier, C.; Greene, S.; Nesmelova, I.V.; Grdzelishvili, V.Z. Experimental Evolution Generates Novel Oncolytic Vesicular Stomatitis Viruses with Improved Replication in Virus-Resistant Pancreatic Cancer Cells. J. Virol. 2020, 94, e01643-19. [Google Scholar] [CrossRef] [Green Version]
- Altomonte, J.; Marozin, S.; Schmid, R.M.; Ebert, O. Engineered newcastle disease virus as an improved oncolytic agent against hepatocellular carcinoma. Mol. Ther. 2010, 18, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Kahn, J.S.; Schnell, M.J.; Buonocore, L.; Rose, J.K. Recombinant vesicular stomatitis virus expressing respiratory syncytial virus (RSV) glycoproteins: RSV fusion protein can mediate infection and cell fusion. Virology 1999, 254, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wollmann, G.; Rogulin, V.; Simon, I.; Rose, J.K.; van den Pol, A.N. Some attenuated variants of vesicular stomatitis virus show enhanced oncolytic activity against human glioblastoma cells relative to normal brain cells. J. Virol. 2010, 84, 1563–1573. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.C.; Hwang, T.; Park, B.H.; Bell, J.; Kirn, D.H. The targeted oncolytic poxvirus JX-594 demonstrates antitumoral, antivascular, and aanti-HBV activities in patients with hepatocellular carcinoma. Mol. Ther. 2008, 16, 1637–1642. [Google Scholar] [CrossRef]
- Breitbach, C.J.; Arulanandam, R.; De Silva, N.; Thorne, S.H.; Patt, R.; Daneshmand, M.; Moon, A.; Ilkow, C.; Burke, J.; Hwang, T.H.; et al. Oncolytic vaccinia virus disrupts tumor-associated vasculature in humans. Cancer Res. 2013, 73, 1265–1275. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, H.; Goldufsky, J.; Sivendran Harcharik, S.; Pan, M.; Bernardo, S.; Stern, R.H.; Friedlander, P.; Ruby, C.; Saenger, Y. Oncolytic virus therapy for cancer. Oncolytic Virother. 2013, 31, 31–46. [Google Scholar] [CrossRef] [Green Version]
- Diaz, R.M.; Galivo, F.; Kottke, T.; Wongthida, P.; Qiao, J.; Thompson, J.; Valdes, M.; Barber, G.; Vile, R.G. Oncolytic immunovirotherapy for melanoma using vesicular stomatitis virus. Cancer Res. 2007, 67, 2840–2848. [Google Scholar] [CrossRef] [Green Version]
- Prestwich, R.J.; Errington, F.; Ilett, E.J.; Morgan, R.S.M.; Scott, K.J.; Kottke, T.; Thompson, J.; Morrison, E.E.; Harrington, K.J.; Pandha, H.S.; et al. Tumor infection by oncolytic reovirus primes adaptive antitumor immunity. Clin. Cancer Res. 2008, 14, 7358–7366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wongthida, P.; Diaz, R.M.; Galivo, F.; Kottke, T.; Thompson, J.; Pulido, J.; Pavelko, K.; Pease, L.; Melcher, A.; Vile, R. Type II IFN interleukin-28 mediates the antitumor efficacy of oncolytic virus VSV in immune-competent mouse models of cancer. Cancer Res. 2010, 70, 4539–4549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobol, P.T.; Boudreau, J.E.; Stephenson, K.; Wan, Y.; Lichty, B.D.; Mossman, K.L. Adaptive antiviral immunity is a determinant of the therapeutic success of oncolytic virotherapy. Mol. Ther. 2011, 19, 335–344. [Google Scholar] [CrossRef]
- Fu, X.; Rivera, A.; Tao, L.; De Geest, B.; Zhang, X. Construction of an oncolytic herpes simplex virus that precisely targets hepatocellular carcinoma cells. Mol. Ther. 2012, 20, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Garg, A.D.; Dudek-Peric, A.M.; Romano, E.; Agostinis, P. Immunogenic cell death. Int. J. Dev. Biol. 2015, 59, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Fucikova, J.; Kepp, O.; Kasikova, L.; Petroni, G.; Yamazaki, T.; Liu, P.; Zhao, L.; Spisek, R.; Kroemer, G.; Galluzzi, L. Detection of immunogenic cell death and its relevance for cancer therapy. Cell Death Dis. 2020, 11, 1013. [Google Scholar] [CrossRef]
- Kasala, D.; Choi, J.W.; Kim, S.W.; Yun, C.O. Utilizing adenovirus vectors for gene delivery in cancer. Expert Opin. Drug Deliv. 2014, 11, 379–392. [Google Scholar] [CrossRef]
- Halldén, G.; Portella, G. Oncolytic virotherapy with modified adenoviruses and novel therapeutic targets. Expert Opin. Ther. Targets 2012, 16, 945–958. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.-J.; Wang, L.-J.; Zhang, Z.; Gao, N.; Liang, C.-Y.; Huang, Y.-D.; Han, B. Selective intra-arterial infusion of rAd-p53 with chemotherapy for advanced oral cancer: A randomized clinical trial. BMC Med. 2014, 12, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemminki, O.; Parviainen, S.; Juhila, J.; Turkki, R.; Linder, N.; Lundin, J.; Kankainen, M.; Ristimäki, A.; Koski, A.; Liikanen, I.; et al. Immunological data from cancer patients treated with Ad5/3-E2F-Δ24-GMCSF suggests utility for tumor immunotherapy. Oncotarget 2015, 6, 4467–4481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreno, B.; Urosevic-Maiwald, M.; Kim, Y.; Guitart, J.; Duvic, M.; Dereure, O.; Khammari, A.; Knol, A.-C.; Derbij, A.; Lusky, M.; et al. TG1042 (Adenovirus-interferon-γ) in Primary Cutaneous B-cell Lymphomas: A Phase II Clinical Trial. PLoS ONE 2014, 9, e83670. [Google Scholar] [CrossRef]
- Dong, J.; Li, W.; Dong, A.; Mao, S.; Shen, L.; Li, S.; Gong, X.; Wu, P. Gene therapy for unresectable hepatocellular carcinoma using recombinant human adenovirus type 5. Med. Oncol. 2014, 31, 95. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.X.; Wang, D.; Wang, G.; Zhang, Q.H.; Liu, J.M.; Peng, P.; Liu, X.H. Clinical study of recombinant adenovirus-p53 combined with fractionated stereotactic radiotherapy for hepatocellular carcinoma. J. Cancer Res. Clin. Oncol. 2010, 136, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Westphal, M.; Ylä-Herttuala, S.; Martin, J.; Warnke, P.; Menei, P.; Eckland, D.; Kinley, J.; Kay, R.; Ram, Z. Adenovirus-mediated gene therapy with sitimagene ceradenovec followed by intravenous ganciclovir for patients with operable high-grade glioma (ASPECT): A randomised, open-label, phase 3 trial. Lancet Oncol. 2013, 14, 823–833. [Google Scholar] [CrossRef]
- Reddi, H.V.; Madde, P.; Cohen, Y.C.; Bangio, L.; Breitbart, E.; Harats, D.; Bible, K.C.; Eberhardt, N.L. Antitumor Activity of VB-111, a Novel Antiangiogenic Virotherapeutic, in Thyroid Cancer Xenograft Mouse Models. Genes Ther. Cancer 2011, 2, 993–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arend, R.C.; Beer, H.M.; Cohen, Y.C.; Berlin, S.; Birrer, M.J.; Campos, S.M.; Rachmilewitz Minei, T.; Harats, D.; Wall, J.A.; Foxall, M.E.; et al. Ofranergene obadenovec (VB-111) in platinum-resistant ovarian cancer; favorable response rates in a phase I/II study are associated with an immunotherapeutic effect. Gynecol. Oncol. 2020, 157, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.Z.; Wang, J.K.; Li, W.; Zhang, X.L. Clinical research on recombinant human Ad-p53 injection combined with cisplatin in treatment of malignant pleural effusion induced by lung cancer. Chin. J. Cancer 2009, 28, 84–87. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.S.; Ma, J.G.; Xing, L.N. Efficacy and safety of recombinant human adenovirus p53 combined with chemoradiotherapy in the treatment of recurrent nasopharyngeal carcinoma. Anticancer Drugs 2016, 28, 230–236. [Google Scholar] [CrossRef]
- Xiao, J.; Zhou, J.; Fu, M.; Liang, L.I.; Deng, Q.; Liu, X.; Liu, F. Efficacy of recombinant human adenovirus-p53 combined with chemotherapy for locally advanced cervical cancer: A clinical trial. Oncol. Lett. 2017, 13, 3676–3680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejada, S.; Alonso, M.; Patiño, A.; Fueyo, J.; Gomez-Manzano, C.; Diez-Valle, R. Phase i Trial of DNX-2401 for Diffuse Intrinsic Pontine Glioma Newly Diagnosed in Pediatric Patients. Clin. Neurosurg. 2018, 83, 1050–1056. [Google Scholar] [CrossRef] [PubMed]
- Li, K.-L.; Kang, J.; Zhang, P.; Li, L.; Wang, Y.-B.; Chen, H.-Y.; He, Y. Efficacy of recombinant adenoviral human p53 gene in the treatment of lung cancer-mediated pleural effusion. Oncol. Lett. 2015, 9, 2193–2198. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Li, Z.; Geng, F.; Fu, Q.; Guo, C.; Xiao, Y.; Zhang, Z.; Li, G. Treatment of recurrent malignant gliomas by surgery combined with recombinant adenovirus-p53. Zhonghua Zhong Liu Za Zhi 2010, 32, 709–712. [Google Scholar]
- Hosmani, J.; Mushtaq, S.; Abullais, S.S.; Almubarak, H.M.; Assiri, K.; Testarelli, L.; Mazzoni, A.; Patil, S. Recombinant Human Adenovirus-p53 Therapy for the Treatment of Oral Leukoplakia and Oral Squamous Cell Carcinoma: A Systematic Review. Medicina 2021, 57, 438. [Google Scholar] [CrossRef]
- Liang, M. Oncorine, the World First Oncolytic Virus Medicine and its Update in China. Curr. Cancer Drug Targets 2018, 18, 171–176. [Google Scholar] [CrossRef]
- Nemunaitis, J.; Khuri, F.; Ganly, I.; Arseneau, J.; Posner, M.; Vokes, E.; Kuhn, J.; McCarty, T.; Landers, S.; Blackburn, A.; et al. Phase II trial of intratumoral administration of ONYX-015, a replication-selective adenovirus, in patients with refractory head and neck cancer. J. Clin. Oncol. 2001, 19, 289–298. [Google Scholar] [CrossRef]
- O’Cathail, S.M.; Davis, S.; Holmes, J.; Brown, R.; Fisher, K.; Seymour, L.; Adams, R.; Good, J.; Sebag-Montefiore, D.; Maughan, T.; et al. A phase 1 trial of the safety, tolerability and biological effects of intravenous Enadenotucirev, a novel oncolytic virus, in combination with chemoradiotherapy in locally advanced rectal cancer (CEDAR). Radiat. Oncol. 2020, 15, 151. [Google Scholar] [CrossRef]
- Machiels, J.-P.; Salazar, R.; Rottey, S.; Duran, I.; Dirix, L.; Geboes, K.; Wilkinson-Blanc, C.; Pover, G.; Alvis, S.; Champion, B.; et al. A phase 1 dose escalation study of the oncolytic adenovirus enadenotucirev, administered intravenously to patients with epithelial solid tumors (EVOLVE). J. Immunother. Cancer 2019, 7, 20. [Google Scholar] [CrossRef]
- Davison, A.J. Herpesvirus systematics. Vet. Microbiol. 2010, 143, 52–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martuza, R.L.; Malick, A.; Marker, J.M.; Ruffner, K.I.; Coen, D.M. Experimental Therapy of Human Glioma by Means of. Science 1991, 252, 854–856. [Google Scholar] [CrossRef]
- Reid, V.; Yu, Z.; Schuman, T.; Li, S.; Singh, P.; Fong, Y.; Wong, R.J. Herpes oncolytic therapy of salivary gland carcinomas. Int. J. Cancer 2008, 122, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Aghi, M.; Martuza, R.L. Oncolytic viral therapies—The clinical experience. Oncogene 2005, 24, 7802–7816. [Google Scholar] [CrossRef] [Green Version]
- Currier, M.A.; Gillespie, R.A.; Sawtell, N.M.; Mahller, Y.Y.; Stroup, G.; Collins, M.H.; Kambara, H.; Chiocca, E.A.; Cripe, T.P. Efficacy and safety of the oncolytic herpes simplex virus rRp450 alone and combined with cyclophosphamide. Mol. Ther. 2008, 16, 879–885. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Herpes Simplex Virus. Available online: https://www.who.int/news-room/fact-sheets/detail/herpes-simplex-virus (accessed on 10 May 2021).
- Ma, W.; He, H.; Wang, H. Oncolytic herpes simplex virus and immunotherapy 11 Medical and Health Sciences 1108 Medical Microbiology. BMC Immunol. 2018, 19, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bommareddy, P.K.; Patel, A.; Hossain, S.; Kaufman, H.L. Talimogene Laherparepvec (T-VEC) and Other Oncolytic Viruses for the Treatment of Melanoma. Am. J. Clin. Dermatol. Ther. 2017, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Andtbacka, R.H.; Agarwala, S.S.; Ollila, D.W.; Hallmeyer, S.; Milhem, M.; Amatruda, T.; Nemunaitis, J.J.; Harrington, K.J.; Chen, L.; Shilkrut, M.; et al. Cutaneous head and neck melanoma in OPTiM, a randomized pha se 3 trial oftalimogene laherparepvec versus granulocyte-macrophage colony-stimulatingfactor for the treatment of unresected stage IIIB/I IIC/IV melanoma. Head Neck 2016, 38, 1752–1758. [Google Scholar] [CrossRef]
- Ressler, J.M.; Karasek, M.; Koch, L.; Silmbrod, R.; Mangana, J.; Latifyan, S.; Aedo-Lopez, V.; Kehrer, H.; Weihsengruber, F.; Koelblinger, P.; et al. Real-life use of talimogene laherparepvec (T-VEC) in melanoma patients in centers in Austria, Switzerland and Germany. J. Immunother. Cancer 2021, 9, e001701. [Google Scholar] [CrossRef] [PubMed]
- Chesney, J.; Puzanov, I.; Collichio, F.; Singh, P.; Milhem, M.M.; Glaspy, J.; Hamid, O.; Ross, M.; Friedlander, P.; Garbe, C.; et al. Randomized, Open-Label Phase II Study Evaluating the Efficacy and Safety of Talimogene Laherparepvec in Combination with Ipilimumab Versus Ipilimumab Alone in Patients with Advanced, Unresectable Melanoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2018, 36, 1658–1667. [Google Scholar] [CrossRef]
- Thomas, S.; Kuncheria, L.; Roulstone, V.; Kyula, J.N.; Mansfield, D.; Bommareddy, P.K.; Smith, H.; Kaufman, H.L.; Harrington, K.J.; Coffin, R.S. Development of a new fusion-enhanced oncolytic immunotherapy platform based on herpes simplex virus type 1. J. Immunother. Cancer 2019, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Middleton, M.R.; Aroldi, F.; Sacco, J.; Milhem, M.M.; Curti, B.D.; Vanderwalde, A.M.; Baum, S.; Samson, A.; Pavlick, A.C.; Chesney, J.A.; et al. An open-label, single-arm, phase II clinical trial of RP1, an enhanced potency oncolytic herpes virus, combined with nivolumab in four solid tumor types: Initial results from the skin cancer cohorts. J. Clin. Oncol. 2020, 38, e22050. [Google Scholar] [CrossRef]
- Haddad, D. Genetically Engineered Vaccinia Viruses as Agents for Cancer Treatment, Imaging, and Transgene Delivery. Front. Oncol. 2017, 7, 96. [Google Scholar] [CrossRef] [Green Version]
- Yaghchi, C.A.; Zhang, Z.; Alusi, G.; Lemoine, N.R.; Wang, Y. Vaccinia virus, a promising new therapeutic agent for pancreatic cancer. Immunotherapy 2015, 7, 1249–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Domínguez, L.E.; McFadden, G. Poxvirus oncolytic virotherapy. Expert Opin. Biol. Ther. 2019, 19, 561–573. [Google Scholar] [CrossRef]
- Gomella, L.G.; Mastrangelo, M.J.; McCue, P.A.; Maguire, H.C.; Mulholland, S.G.; Lattime, E.C. Phase I study of intravesical vaccinia virus as a vector for gene therapy of bladder cancer. J. Urol. 2001, 166, 1291–1295. [Google Scholar] [CrossRef]
- Minev, B.R.; Lander, E.; Feller, J.F.; Berman, M.; Greenwood, B.M.; Minev, I.; Santidrian, A.F.; Nguyen, D.; Draganov, D.; Killinc, M.O.; et al. First-in-human study of TK-positive oncolytic vaccinia virus delivered by adipose stromal vascular fraction cells. J. Transl. Med. 2019, 17, 271. [Google Scholar] [CrossRef] [PubMed]
- Zeh, H.J.; Downs-Canner, S.; McCart, J.A.; Guo, Z.S.; Rao, U.N.M.; Ramalingam, L.; Thorne, S.H.; Jones, H.L.; Kalinski, P.; Wieckowski, E.; et al. First-in-man study of western reserve strain oncolytic vaccinia virus: Safety, systemic spread, and antitumor activity. Mol. Ther. 2015, 23, 202–214. [Google Scholar] [CrossRef] [Green Version]
- Downs-Canner, S.; Guo, Z.S.; Ravindranathan, R.; Breitbach, C.J.; O’Malley, M.E.; Jones, H.L.; Moon, A.; McCart, J.A.; Shuai, Y.; Zeh, H.J.; et al. Phase 1 Study of Intravenous Oncolytic Poxvirus (vvDD) in Patients with Advanced Solid Cancers. Mol. Ther. 2016, 24, 1492–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauer, U.M.; Schell, M.; Beil, J.; Berchtold, S.; Koppenhofer, U.; Glatzle, J.; Konigsrainer, A.; Mohle, R.; Nann, D.; Fend, F.; et al. Phase I study of oncolytic vaccinia virus GL-ONC1 in patients with peritoneal carcinomatosis. Clin. Cancer Res. 2018, 24, 4388–4398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mell, L.K.; Brumund, K.T.; Daniels, G.A.; Advani, S.J.; Zakeri, K.; Wright, M.E.; Onyeama, S.J.; Weisman, R.A.; Sanghvi, P.R.; Martin, P.J.; et al. Phase I trial of intravenous oncolytic vaccinia virus (GL-ONC1) with cisplatin and radiotherapy in patients with locoregionally advanced head and neck carcinoma. Clin. Cancer Res. 2017, 23, 5696–5702. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.; Reid, T.; Ruo, L.; Breitbach, C.J.; Rose, S.; Bloomston, M.; Cho, M.; Lim, H.Y.; Chung, H.C.; Kim, C.W.; et al. Randomized dose-finding clinical trial of oncolytic immunotherapeutic vaccinia JX-594 in liver cancer. Nat. Med. 2013, 19, 329–336. [Google Scholar] [CrossRef]
- Moehler, M.; Heo, J.; Lee, H.C.; Tak, W.Y.; Chao, Y.; Paik, S.W.; Yim, H.J.; Byun, K.S.; Baron, A.; Ungerechts, G.; et al. Vaccinia-based oncolytic immunotherapy Pexastimogene Devacirepvec in patients with advanced hepatocellular carcinoma after sorafenib failure: A randomized multicenter Phase IIb trial (TRAVERSE). Oncoimmunology 2019, 8, 1615817. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-G.; Ha, H.K.; Lim, S.; De Silva, N.S.; Pelusio, A.; Mun, J.H.; Patt, R.H.; Breitbach, C.J.; Burke, J.M. Phase II trial of pexa-vec (pexastimogene devacirepvec; JX-594), an oncolytic and immunotherapeutic vaccinia virus, in patients with metastatic, refractory renal cell carcinoma (RCC). J. Clin. Oncol. 2018, 36, 671. [Google Scholar] [CrossRef]
- Samson, A.; West, E.; Turnbull, S.; Scott, K.; Tidswell, E.; Kingston, J.; Johnpulle, M.; Bendjama, K.; Stojkowitz, N.; Lusky, M.; et al. Single intravenous preoperative administration of the oncolytic virus Pexa-Vec to prime anti-tumour immunity. Ann. Oncol. 2019, 30, v496. [Google Scholar] [CrossRef]
- Gulley, J.L.; Borre, M.; Vogelzang, N.J.; Ng, S.; Agarwal, N.; Parker, C.C.; Pook, D.W.; Rathenborg, P.; Flaig, T.W.; Carles, J.; et al. Phase III Trial of PROSTVAC in Asymptomatic or Minimally Symptomatic Metastatic Castration-Resistant Prostate Cancer. J. Clin. Oncol. 2019, 37, 1051–1061. [Google Scholar] [CrossRef] [PubMed]
- Heery, C.R.; Palena, C.; Mcmahon, S.; Donahue, R.N.; Lepone, L.M.; Grenga, I.; Dirmeier, U.; Cordes, L.; Mart, J.; Dahut, W.; et al. Phase I Study of a Poxviral TRICOM-Based Vaccine Directed Against the Transcription Factor Brachyury. Clin Cancer Res. 2017, 23, 6846. [Google Scholar] [CrossRef] [Green Version]
- Simmonds, P.; Gorbalenya, A.E.; Harvala, H.; Hovi, T.; Knowles, N.J.; Lindberg, A.M.; Oberste, M.S.; Palmenberg, A.C.; Reuter, G.; Skern, T.; et al. Recommendations for the nomenclature of enteroviruses and rhinoviruses. Arch. Virol. 2020, 165, 793–797. [Google Scholar] [CrossRef] [Green Version]
- Annels, N.E.; Mansfield, D.; Arif, M.; Ballesteros-Merino, C.; Simpson, G.R.; Denyer, M.; Sandhu, S.S.; Melcher, A.A.; Harrington, K.J.; Davies, B.; et al. Phase I Trial of an ICAM-1-Targeted Immunotherapeutic-Coxsackievirus A21 (CVA21) as an Oncolytic Agent Against Non Muscle-Invasive Bladder Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2019, 25, 5818–5831. [Google Scholar] [CrossRef] [PubMed]
- Doniņa, S.; Strēle, I.; Proboka, G.; Auziņš, J.; Alberts, P.; Jonsson, B.; Venskus, D.; Muceniece, A. Adapted ECHO-7 virus Rigvir immunotherapy (oncolytic virotherapy) prolongs survival in melanoma patients after surgical excision of the tumour in a retrospective study. Melanoma Res. 2015, 25, 421–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voroshilova, M. Virological and immunological aspects of administration of live enteroviral vaccines in oncological disease. In Poleznye Dlya Ogranizma Nepatogennye Shtammy Enterovirusov: Profilakticheskoe i Lechebnoe ikh Primenenie (Useful Nonpathogenic Enterovirus Strains: Preventive and Therapeutic Applications); Meditsina: Moscow, Russia, 1988; pp. 24–29. [Google Scholar]
- Holl, E.K.; Brown, M.C.; Boczkowski, D.; McNamara, M.A.; George, D.J.; Bigner, D.D.; Gromeier, M.; Nair, S.K. Recombinant oncolytic poliovirus, PVSRIPO, has potent cytotoxic and innate inflammatory effects, mediating therapy in human breast and prostate cancer xenograft models. Oncotarget 2016, 7, 79828–79841. [Google Scholar] [CrossRef] [Green Version]
- Ochiai, H.; Campbell, S.A.; Archer, G.E.; Chewning, T.A.; Dragunsky, E.; Ivanov, A.; Gromeier, M.; Sampson, J.H. Targeted therapy for glioblastoma multiforme neoplastic meningitis with intrathecal delivery of an oncolytic recombinant poliovirus. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 1349–1354. [Google Scholar] [CrossRef] [Green Version]
- Bradley, S.; Jakes, A.D.; Harrington, K.; Pandha, H.; Melcher, A.; Errington-Mais, F. Applications of coxsackievirus A21 in oncology. Oncolytic Virother. 2014, 3, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Au, G.G.; Beagley, L.G.; Haley, E.S.; Barry, R.D.; Shafren, D.R. Oncolysis of malignant human melanoma tumors by Coxsackieviruses A13, A15 and A18. Virol. J. 2011, 8, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.K.; Hong, J.; Yun, C.-O. Oncolytic Viruses and Immune Checkpoint Inhibitors: Preclinical Developments to Clinical Trials. Int. J. Mol. Sci. 2020, 21, 8627. [Google Scholar] [CrossRef]
- Bodian, D. Histopathologic basis of clinical findings in poliomyelitis. Am. J. Med. 1949, 6, 563–578. [Google Scholar] [CrossRef]
- Desjardins, A.; Gromeier, M.; Herndon, J.E.; Beaubier, N.; Bolognesi, D.P.; Friedman, A.H.; Friedman, H.S.; McSherry, F.; Muscat, A.M.; Nair, S.; et al. Recurrent Glioblastoma Treated with Recombinant Poliovirus. N. Engl. J. Med. 2018, 379, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.W.; Souhrada, M.; Prather, S. Viruses as an Aid to Cancer Therapy: Regression of Solid and Ascites Tumors in Rodents After Treatment with Bovine Enterovirus. Proc. Natl. Acad. Sci. USA 1971, 68, 836–840. [Google Scholar] [CrossRef] [Green Version]
- Voroshilova, M. Enterovirusnyye Infektsii Cheloveka (Human Enteroviral Infections); Meditsina: Moscow, Russia, 1979. [Google Scholar]
- Chumakov, M.; Voroshilova, M.; Shindarov, L.; Lavrova, I.; Gracheva, L.; Koroleva, G.; Vasilenko, S.; Brodvarova, I.; Nikolova, M.; Gyurova, S.; et al. Enterovirus 71 isolated from cases of epidemic poliomyelitis-like disease in Bulgaria. Arch. Virol. 1979, 60, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Chumakov, P.M.; Morozova, V.V.; Babkin, I.V.; Baĭkov, I.K.; Netesov, S.V.; Tikunova, N.V. Oncolytic enteroviruses. Mol. Biol. 2012, 46, 712–725. [Google Scholar] [CrossRef]
- Voroshilova, M.; Tol’skaya, E.; Koroleva, G.; Chumakov, K.; Chumakov, P. Studies on biological and morphological properties of viruses ECHO1 and ECHO12. Tr. Inst. Poliom. Virus. Entsef. Akad. Med. Nauk SSSR 1970, XIV, 269–274. [Google Scholar]
- Voroshilova, M.; Baganova, N. Experience in treating patients with gastrointestinal tumors by live enterovirus vaccines. In Virusnyi Onkoliz i Iskusstvennaya Geterogenizatsiya Opukholei (Viral Oncolysis and Artificial Heterogenization of Tumors); Riga, Latvia, 1969; pp. 23–26. [Google Scholar]
- Soboleva, A.; Lipatova, A.; Kochetkov, D.; Chumakov, P. Changes in the sensitivity of human glioblastoma cells to oncolytic enteroviruses induced by passaging. Bull. RSMU 2018, 37–41. [Google Scholar] [CrossRef]
- Sidorenko, A.; Zheltukhin, A.; Le, T.; Soboleva, A.; Lipatova, A.; Golbin, D.; Chumakov, P. Persistence of oncolytic Coxsackie virus A7 in subcutaneous human glioblastoma xenografts in mice in the context of experimental therapy. Bull. RSMU 2018, 41–46. [Google Scholar] [CrossRef]
- Zheltukhin, A.; Soboleva, A.; Sosnovtseva, A.; Le, T.; Ilyinskaya, G.; Kochetkov, D.; Lipatova, A.; Chumakov, P. Human enteroviruses exhibit selective oncolytic activity in the model of human glioblastoma multiforme xenografts in immunodeficient mice. Bull. RSMU 2018, 42–48. [Google Scholar] [CrossRef]
- Podshivalova, E.S.; Semkina, A.S.; Kravchenko, D.S.; Frolova, E.I.; Chumakov, S.P. Efficient delivery of oncolytic enterovirus by carrier cell line NK-92. Mol. Ther. Oncolytics 2021, 21, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Alberts, P.; Tilgase, A.; Rasa, A.; Bandere, K.; Venskus, D. The advent of oncolytic virotherapy in oncology: The Rigvir® story. Eur. J. Pharm. Ther. 2018, 837, 117–126. [Google Scholar] [CrossRef]
- Pumpure, E.; Dručka, E.; Kigitoviča, D.; Meškauskas, R.; Isajevs, S.; Nemiro, I.; Rasa, A.; Olmane, E.; Zablocka, T.; Alberts, P.; et al. Management of a primary malignant melanoma of uterine cervix stage IVA patient with radical surgery and adjuvant oncolytic virus Rigvir(®) therapy: A case report. Clin. Case Rep. 2020, 8, 1538–1543. [Google Scholar] [CrossRef] [PubMed]
- Jaunalksne, I.; Brokāne, L.; Petroška, D.; Rasa, A.; Alberts, P. ECHO-7 oncolytic virus Rigvir® in an adjuvant setting for stage I uveal melanoma; A retrospective case report. Am. J. Ophthalmol. Case Rep. 2020, 17, 100615. [Google Scholar] [CrossRef]
- Babiker, H.M.; Riaz, I.B.; Husnain, M.; Borad, M.J. Oncolytic virotherapy including Rigvir and standard therapies in malignant melanoma. Oncolytic Virother. 2017, 6, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Proboka, G.; Rasa, A.; Olmane, E.; Isajevs, S.; Tilgase, A.; Alberts, P. Treatment of a stage III rima glottidis patient with the oncolytic virus Rigvir: A retrospective case report. Medicine 2019, 98, e17883. [Google Scholar] [CrossRef]
- Beasley, G.M.; Nair, S.K.; Farrow, N.E.; Landa, K.; Selim, M.A.; Wiggs, C.A.; Jung, S.-H.; Bigner, D.D.; True Kelly, A.; Gromeier, M.; et al. Phase I trial of intratumoral PVSRIPO in patients with unresectable, treatment-refractory melanoma. J. Immunother. Cancer 2021, 9, e002203. [Google Scholar] [CrossRef]
- Fallaux, F.J.; Bout, A.; van der Velde, I.; van den Wollenberg, D.J.; Hehir, K.M.; Keegan, J.; Auger, C.; Cramer, S.J.; van Ormondt, H.; van der Eb, A.J.; et al. New helper cells and matched early region 1-deleted adenovirus vectors prevent generation of replication-competent adenoviruses. Hum. Gene Ther. 1998, 9, 1909–1917. [Google Scholar] [CrossRef] [PubMed]
- Schenk, E.L.; Mandrekar, S.J.; Dy, G.K.; Aubry, M.C.; Tan, A.D.; Dakhil, S.R.; Sachs, B.A.; Nieva, J.J.; Bertino, E.; Hann, C.L.; et al. A Randomized Double-Blind Phase II Study of the Seneca Valley Virus ( NTX-010 ) versus Placebo for Patients with Extensive-Stage SCLC ( ES SCLC ) Who Were Stable or Responding after at Least Four Cycles of Platinum-Based Chemotherapy: North Central Cance. J. Thorac. Oncol. 2019, 15, 110–119. [Google Scholar] [CrossRef]
- Rima, B.; Balkema-Buschmann, A.; Dundon, W.G.; Duprex, P.; Easton, A.; Fouchier, R.; Kurath, G.; Lamb, R.; Lee, B.; Rota, P.; et al. ICTV Virus Taxonomy Profile: Paramyxoviridae. J. Gen. Virol. 2019, 100, 1593–1594. [Google Scholar] [CrossRef] [PubMed]
- Lorence, R.M.; Reichard, K.W.; Katubig, B.B.; Reyes, H.M.; Phuangsab, A.; Mitchell, B.R.; Cascino, C.J.; Walter, R.J.; Peeples, M.E. Complete regression of human neuroblastoma xenografts in athymic mice after local Newcastle disease virus therapy. J. Natl. Cancer Inst. 1994, 86, 1228–1233. [Google Scholar] [CrossRef]
- Galanis, E. Therapeutic potential of oncolytic measles virus: Promises and challenges. Clin. Pharm. Ther. 2010, 88, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.-W.; Hadac, E.M.; Anderson, B.D.; Myers, R.; Harvey, M.; Greiner, S.M.; Soeffker, D.; Federspiel, M.J.; Russell, S.J. Pharmacokinetics of oncolytic measles virotherapy: Eventual equilibrium between virus and tumor in an ovarian cancer xenograft model. Cancer Gene Ther. 2006, 13, 732–738. [Google Scholar] [CrossRef] [Green Version]
- Msaouel, P.; Iankov, I.D.; Allen, C.; Aderca, I.; Federspiel, M.J.; Tindall, D.J.; Morris, J.C.; Koutsilieris, M.; Russell, S.J.; Galanis, E. Noninvasive imaging and radiovirotherapy of prostate cancer using an oncolytic measles virus expressing the sodium iodide symporter. Mol. Ther. 2009, 17, 2041–2048. [Google Scholar] [CrossRef]
- Sinkovics, J.G.; Horvath, J.C. Newcastle disease virus (NDV): Brief history of its oncolytic strains. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2000, 16, 1–15. [Google Scholar] [CrossRef]
- Cassel, W.A.; Murray, D.R. A ten-year follow-up on stage II malignant melanoma patients treated postsurgically with Newcastle disease virus oncolysate. Med. Oncol. Tumor Pharm. Ther. 1992, 9, 169–171. [Google Scholar] [CrossRef]
- Batliwalla, F.M.; Bateman, B.A.; Serrano, D.; Murray, D.; Macphail, S.; Maino, V.C.; Ansel, J.C.; Gregersen, P.K.; Armstrong, C.A. A 15-year follow-up of AJCC stage III malignant melanoma patients treated postsurgically with Newcastle disease virus (NDV) oncolysate and determination of alterations in the CD8 T cell repertoire. Mol. Med. 1998, 4, 783–794. [Google Scholar] [CrossRef]
- Kirchner, H.H.; Anton, P.; Atzpodien, J. Adjuvant treatment of locally advanced renal cancer with autologous virus-modified tumor vaccines. World J. Urol. 1995, 13, 171–173. [Google Scholar] [CrossRef]
- Ahlert, T.; Sauerbrei, W.; Bastert, G.; Ruhland, S.; Bartik, B.; Simiantonaki, N.; Schumacher, J.; Häcker, B.; Schumacher, M.; Schirrmacher, V. Tumor-cell number and viability as quality and efficacy parameters of autologous virus-modified cancer vaccines in patients with breast or ovarian cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 1997, 15, 1354–1366. [Google Scholar] [CrossRef] [PubMed]
- Ockert, D.; Schirrmacher, V.; Beck, N.; Stoelben, E.; Ahlert, T.; Flechtenmacher, J.; Hagmüller, E.; Buchcik, R.; Nagel, M.; Saeger, H.D. Newcastle disease virus-infected intact autologous tumor cell vaccine for adjuvant active specific immunotherapy of resected colorectal carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 1996, 2, 21–28. [Google Scholar]
- Schirrmacher, V. Fifty Years of Clinical Application of Newcastle Disease Virus: Time to Celebrate! Biomedicines 2016, 4, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burman, B.; Pesci, G.; Zamarin, D. Newcastle Disease Virus at the Forefront of Cancer Immunotherapy. Cancers 2020, 12, 3552. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; He, J.; Zhong, L.; Zhao, Y. Advances in the Study of Antitumour Immunotherapy for Newcastle Disease Virus. Int. J. Med. Sci. 2021, 18, 2294–2302. [Google Scholar] [CrossRef]
- Cassel, W.A.; Garrett, R.E. Newcastle disease virus as an antineoplastic agent. Cancer 1965, 18, 863–868. [Google Scholar] [CrossRef]
- Freeman, A.I.; Zakay-Rones, Z.; Gomori, J.M.; Linetsky, E.; Rasooly, L.; Greenbaum, E.; Rozenman-Yair, S.; Panet, A.; Libson, E.; Irving, C.S.; et al. Phase I/II trial of intravenous NDV-HUJ oncolytic virus in recurrent glioblastoma multiforme. Mol. Ther. 2006, 13, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Pecora, A.L.; Rizvi, N.; Cohen, G.I.; Meropol, N.J.; Sterman, D.; Marshall, J.L.; Goldberg, S.; Gross, P.; O’Neil, J.D.; Groene, W.S.; et al. Phase I trial of intravenous administration of PV701, an oncolytic virus, in patients with advanced solid cancers. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2002, 20, 2251–2266. [Google Scholar] [CrossRef]
- Csatary, L.K.; Gosztonyi, G.; Szeberenyi, J.; Fabian, Z.; Liszka, V.; Bodey, B.; Csatary, C.M. MTH-68/H oncolytic viral treatment in human high-grade gliomas. J. Neurooncol. 2004, 67, 83–93. [Google Scholar] [CrossRef]
- Kobayashi, H. Viral xenogenization of intact tumor cells. Adv. Cancer Res. 1979, 30, 279–299. [Google Scholar] [CrossRef] [PubMed]
- Heicappell, R.; Schirrmacher, V.; von Hoegen, P.; Ahlert, T.; Appelhans, B. Prevention of metastatic spread by postoperative immunotherapy with virally modified autologous tumor cells. I. Parameters for optimal therapeutic effects. Int. J. Cancer 1986, 37, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, V.; Heicappell, R. Prevention of metastatic spread by postoperative immunotherapy with virally modified autologous tumor cells. II. Establishment of specific systemic anti-tumor immunity. Clin. Exp. Metastasis 1987, 5, 147–156. [Google Scholar] [CrossRef]
- Von Hoegen, P.; Weber, E.; Schirrmacher, V. Modification of tumor cells by a low dose of Newcastle disease virus. Augmentation of the tumor-specific T cell response in the absence of an anti-viral response. Eur. J. Immunol. 1988, 18, 1159–1166. [Google Scholar] [CrossRef]
- Schild, H.; von Hoegen, P.; Schirrmacher, V. Modification of tumor cells by a low dose of Newcastle disease virus. II. Augmented tumor-specific T cell response as a result of CD4+ and CD8+ immune T cell cooperation. Cancer Immunol. Immunother. 1989, 28, 22–28. [Google Scholar] [CrossRef] [PubMed]
- von Hoegen, P.; Zawatzky, R.; Schirrmacher, V. Modification of tumor cells by a low dose of Newcastle disease virus. III. Potentiation of tumor-specific cytolytic T cell activity via induction of interferon-alpha/beta. Cell. Immunol. 1990, 126, 80–90. [Google Scholar] [CrossRef]
- Karcher, J.; Dyckhoff, G.; Beckhove, P.; Reisser, C.; Brysch, M.; Ziouta, Y.; Helmke, B.H.; Weidauer, H.; Schirrmacher, V.; Herold-Mende, C. Antitumor vaccination in patients with head and neck squamous cell carcinomas with autologous virus-modified tumor cells. Cancer Res. 2004, 64, 8057–8061. [Google Scholar] [CrossRef] [Green Version]
- Steiner, H.H.; Bonsanto, M.M.; Beckhove, P.; Brysch, M.; Geletneky, K.; Ahmadi, R.; Schuele-Freyer, R.; Kremer, P.; Ranaie, G.; Matejic, D.; et al. Antitumor vaccination of patients with glioblastoma multiforme: A pilot study to assess feasibility, safety, and clinical benefit. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2004, 22, 4272–4281. [Google Scholar] [CrossRef]
- Schirrmacher, V.; Fournier, P.; Schlag, P. Autologous tumor cell vaccines for post-operative active-specific immunotherapy of colorectal carcinoma: Long-term patient survival and mechanism of function. Expert Rev. Vaccines 2014, 13, 117–130. [Google Scholar] [CrossRef]
- Pomer, S.; Schirrmacher, V.; Thiele, R.; Lohrke, H.; Brkovic, D.; Staehler, G. Tumor Response and 4 year survival-data of patients with advanced renal-cell carcinoma treated with autologous tumor vaccine and subcutaneous R-Il-2 And Ifn-Alpha(2b). Int. J. Oncol. 1995, 6, 947–954. [Google Scholar] [CrossRef]
- Heinzerling, L.; Künzi, V.; Oberholzer, P.A.; Kündig, T.; Naim, H.; Dummer, R. Oncolytic measles virus in cutaneous T-cell lymphomas mounts antitumor immune responses in vivo and targets interferon-resistant tumor cells. Blood 2005, 106, 2287–2294. [Google Scholar] [CrossRef] [Green Version]
- Matveeva, O.V.; Guo, Z.S.; Senin, V.M.; Senina, A.V.; Shabalina, S.A.; Chumakov, P.M. Oncolysis by paramyxoviruses: Preclinical and clinical studies. Mol. Ther. Oncolytics 2015, 2, 15017. [Google Scholar] [CrossRef]
- Belova, A.A.; Sosnovtseva, A.O.; Lipatova, A.V.; Njushko, K.M.; Volchenko, N.N.; Belyakov, M.M.; Sudalenko, O.V.; Krasheninnikov, A.A.; Shegai, P.V.; Sadritdinova, A.F.; et al. Biomarkers of prostate cancer sensitivity to the Sendai virus. Mol. Biol. 2017, 51, 80–88. [Google Scholar] [CrossRef]
- Zainutdinov, S.S.; Tikunov, A.Y.; Matveeva, O.V.; Netesov, S.V.; Kochneva, G. V Complete Genome Sequence of the Oncolytic Sendai virus Strain Moscow. Genome Announc. 2016, 4, e00818-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senin, V.M.; Matveeva, O.V.S.A.V. Method for cancer immunotherapy and pharmaceutical compositions based on oncolytic non-pathogenic sendai virus. US Patent 9526779B2, 27 December 2014. [Google Scholar]
- Nakamura, T.; Russell, S.J. Oncolytic measles viruses for cancer therapy. Expert Opin. Biol. Ther. 2004, 4, 1685–1692. [Google Scholar] [CrossRef] [PubMed]
- Blechacz, B.; Russell, S.J. Measles virus as an oncolytic vector platform. Curr. Gene Ther. 2008, 8, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Galanis, E.; Hartmann, L.C.; Cliby, W.A.; Long, H.J.; Peethambaram, P.P.; Barrette, B.A.; Kaur, J.S.; Haluska, P.J.J.; Aderca, I.; Zollman, P.J.; et al. Phase I trial of intraperitoneal administration of an oncolytic measles virus strain engineered to express carcinoembryonic antigen for recurrent ovarian cancer. Cancer Res. 2010, 70, 875–882. [Google Scholar] [CrossRef] [Green Version]
- Galanis, E.; Atherton, P.J.; Maurer, M.J.; Knutson, K.L.; Dowdy, S.C.; Cliby, W.A.; Haluska, P.J.; Long, H.J.; Oberg, A.; Aderca, I.; et al. Oncolytic measles virus expressing the sodium iodide symporter to treat drug-resistant ovarian cancer. Cancer Res. 2015, 75, 22–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dispenzieri, A.; Tong, C.; LaPlant, B.; Lacy, M.Q.; Laumann, K.; Dingli, D.; Zhou, Y.; Federspiel, M.J.; Gertz, M.A.; Hayman, S.; et al. Phase I trial of systemic administration of Edmonston strain of measles virus genetically engineered to express the sodium iodide symporter in patients with recurrent or refractory multiple myeloma. Leukemia 2017, 31, 2791–2798. [Google Scholar] [CrossRef] [PubMed]
- Masson, P.; Hulo, C.; Castro, E.; De Bitter, H.; Gruenbaum, L.; Essioux, L.; Bougueleret, L.; Xenarios, I.; Mercier, P.; Le Geneva, C. ViralZone: Recent updates to the virus knowledge resource. Nucleic Acid Res. 2013, 41, 579–583. [Google Scholar] [CrossRef] [Green Version]
- Rose, J.; Schubert, M. Rhabdovirus Genomes and Their Products BT-The Rhabdoviruses; Wagner, R.R., Ed.; Springer US: Boston, MA, USA, 1987; pp. 129–166. ISBN 978-1-4684-7032-1. [Google Scholar]
- Faria, P.A.; Chakraborty, P.; Levay, A.; Barber, G.N.; Ezelle, H.J.; Enninga, J.; Arana, C.; van Deursen, J.; Fontoura, B.M.A. VSV disrupts the Rae1/mrnp41 mRNA nuclear export pathway. Mol. Cell 2005, 17, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Jenni, S.; Bloyet, L.M.; Diaz-Avalos, R.; Liang, B.; Whelan, S.P.J.; Grigorieff, N.; Harrison, S.C. Structure of the Vesicular Stomatitis Virus L Protein in Complex with Its Phosphoprotein Cofactor. Cell Rep. 2020, 30, 53.e5–60.e5. [Google Scholar] [CrossRef] [Green Version]
- Rahmeh, A.A.; Morin, B.; Schenk, A.D.; Liang, B.; Heinrich, B.S.; Brusic, V.; Walz, T.; Whelan, S.P.J. Critical phosphoprotein elements that regulate polymerase architecture and function in vesicular stomatitis virus. Proc. Natl. Acad. Sci. USA 2012, 109, 14628–14633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, B.; Whelan, S.P.J. Sensitivity of the polymerase of vesicular stomatitis virus to 2’ substitutions in the template and nucleotide triphosphate during initiation and elongation. J. Biol. Chem. 2014, 289, 9961–9969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisbert, T.W.; Feldmann, H. Recombinant vesicular stomatitis virus-based vaccines against Ebola and Marburg virus infections. J. Infect. Dis. 2011, 204, 1075–1081. [Google Scholar] [CrossRef]
- Zemp, F.; Rajwani, J.; Mahoney, D.J. Rhabdoviruses as vaccine platforms for infectious disease and cancer. Biotechnol. Genet. Eng. Rev. 2018, 34, 122–138. [Google Scholar] [CrossRef]
- Velazquez-Salinas, L.; Naik, S.; Pauszek, S.J.; Peng, K.-W.; Russell, S.J.; Rodriguez, L.L. Oncolytic Recombinant Vesicular Stomatitis Virus (VSV) Is Nonpathogenic and Nontransmissible in Pigs, a Natural Host of VSV. Hum. Gene Ther. Clin. Dev. 2017, 28, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Prioritizing diseases for research and development in emergency contexts. Available online: https://www.who.int/activities/prioritizing-diseases-for-research-and-development-in-emergency-contexts (accessed on 10 May 2021).
- Fathi, A.; Dahlke, C.; Addo, M.M. Recombinant vesicular stomatitis virus vector vaccines for WHO blueprint priority pathogens. Hum. Vaccines Immunother. 2019, 15, 2269–2285. [Google Scholar] [CrossRef] [Green Version]
- Felt, S.A.; Grdzelishvili, V.Z. Recent advances in vesicular stomatitis virus-based oncolytic virotherapy: A 5-year update. J. Gen. Virol. 2017, 98, 2895–2911. [Google Scholar] [CrossRef] [PubMed]
- Bishnoi, S.; Tiwari, R.; Gupta, S.; Byrareddy, S.N.; Nayak, D. Oncotargeting by Vesicular Stomatitis Virus (VSV): Advances in Cancer Therapy. Viruses 2018, 10, 90. [Google Scholar] [CrossRef] [Green Version]
- Willmon, C.L.; Saloura, V.; Fridlender, Z.G.; Wongthida, P.; Diaz, R.M.; Thompson, J.; Kottke, T.; Federspiel, M.; Barber, G.; Albelda, S.M.; et al. Expression of IFN-beta enhances both efficacy and safety of oncolytic vesicular stomatitis virus for therapy of mesothelioma. Cancer Res. 2009, 69, 7713–7720. [Google Scholar] [CrossRef] [Green Version]
- Trottier, M.D.; Palian, B.M.; Reiss, C.S. VSV replication in neurons is inhibited by type I IFN at multiple stages of infection. Virology 2005, 333, 215–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merchan, J.R.; Patel, M.; Cripe, T.P.; Old, M.O.; Strauss, J.F.; Thomassen, A.; Diaz, R.M.; Peng, K.W.; Russell, S.J.; Russell, L.; et al. Relationship of infusion duration to safety, efficacy, and pharmacodynamics (PD): Second part of a phase I-II study using VSV-IFNβ-NIS (VV1) oncolytic virus in patients with refractory solid tumors. J. Clin. Oncol. 2020, 38, 3090. [Google Scholar] [CrossRef]
- Bakkum-Gamez, J.; Block, M.S.; Packiriswamy, N.; Brunton, B.A.; Deepak, U.; Mitchell, J.M.; Suksanpaisan, L.; Atherton, P.; Dueck, A.; Russell, S.J.; et al. Abstract CT072: First in human (FIH) dose escalation studies of intravenous administration of VSV-IFNβ-NIS (Voyager-V1™) in Stage IV or recurrent endometrial cancer. Cancer Res. 2018, 78, CT072. [Google Scholar] [CrossRef]
- Phase I Trial of Systemic Administration of Vesicular Stomatitis Virus Genetically Engineered to Express NIS and Human Interferon Beta, in Patients with Relapsed or Refractory Multiple Myeloma (MM), Acute Myeloid Leukemia (AML), and T-Cell Neoplasms (TCL). Available online: https://ash.confex.com/ash/2020/webprogram/Paper140853.html (accessed on 12 May 2021).
- Qiu, C.; Li, P.; Bi, J.; Wu, Q.; Lu, L.; Qian, G.; Jia, R.; Jia, R. Differential expression of TYRP1 in adult human retinal pigment epithelium and uveal melanoma cells. Oncol. Lett. 2016, 11, 2379–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Camino, V.; Rajani, K.; Kottke, T.; Rommelfanger-Konkol, D.; Zaidi, S.; Thompson, J.; Pulido, J.; Ilett, E.; Donnelly, O.; Selby, P.; et al. The profile of tumor antigens which can be targeted by immunotherapy depends upon the tumor’s anatomical site. Mol. Ther. 2014, 22, 1936–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulido, J.; Kottke, T.; Thompson, J.; Galivo, F.; Wongthida, P.; Diaz, R.M.; Rommelfanger, D.; Ilett, E.; Pease, L.; Pandha, H.; et al. Articles Using virally expressed melanoma cDNA libraries to identify tumor-associated antigens that cure melanoma. Nat. Biotechnol. 2012, 30, 337–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeping, S.; Xu, Y.; Chen, C.I.; Cope, S.; Mojebi, A.; Kuznik, A.; Konidaris, G.; Ayers, D.; Sasane, M.; Allen, R.; et al. Comparative efficacy of cemiplimab versus other systemic treatments for advanced cutaneous squamous cell carcinoma. Futur. Oncol. 2021, 17, 611–627. [Google Scholar] [CrossRef] [PubMed]
- Peled, N.; Oton, A.B.; Hirsch, F.R.; Bunn, P. MAGE A3 antigen-specific cancer immunotherapeutic. Immunotherapy 2009, 1, 19–25. [Google Scholar] [CrossRef]
- Pal, A.; Kundu, R. Human Papillomavirus E6 and E7: The Cervical Cancer Hallmarks and Targets for Therapy. Front. Microbiol. 2019, 10, 3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moehler, M.H.; Zeidler, M.; Wilsberg, V.; Cornelis, J.J.; Woelfel, T.; Rommelaere, J.; Galle, P.R.; Heike, M. Parvovirus H-1-induced tumor cell death enhances human immune response in vitro via increased phagocytosis, maturation, and cross-presentation by dendritic cells. Hum. Gene Ther. 2005, 16, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Rommelaere, J.; Geletneky, K.; Angelova, A.L.; Daeffler, L.; Dinsart, C.; Kiprianova, I.; Schlehofer, J.R.; Raykov, Z. Oncolytic parvoviruses as cancer therapeutics. Cytokine Growth Factor Rev. 2010, 21, 185–195. [Google Scholar] [CrossRef]
- Bhat, R.; Dempe, S.; Dinsart, C.; Rommelaere, J. Enhancement of NK cell antitumor responses using an oncolytic parvovirus. Int. J. Cancer 2011, 128, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Geletneky, K.; Herrero, Y.; Calle, M.; Rommelaere, J.; Schlehofer, J.R. Oncolytic potential of rodent parvoviruses for cancer therapy in humans: A brief review. J. Vet. Med. B. Infect. Dis. Vet. Public Health 2005, 52, 327–330. [Google Scholar] [CrossRef]
- Geletneky, K.; Hajda, J.; Angelova, A.L.; Leuchs, B.; Capper, D.; Bartsch, A.J.; Neumann, J.-O.; Schöning, T.; Hüsing, J.; Beelte, B.; et al. Oncolytic H-1 Parvovirus Shows Safety and Signs of Immunogenic Activity in a First Phase I/IIa Glioblastoma Trial. Mol. Ther. 2017, 25, 2620–2634. [Google Scholar] [CrossRef] [Green Version]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; Junglen, S.; et al. Changes to virus taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2019). Arch. Virol. 2019, 164, 2417–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertens, P. The dsRNA viruses. Virus Res. 2004, 101, 3–13. [Google Scholar] [CrossRef]
- Coffey, M.C.; Strong, J.E.; Forsyth, P.A.; Lee, P.W. Reovirus therapy of tumors with activated Ras pathway. Science 1998, 282, 1332–1334. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.; Tyler, K.L. Down-regulation of cFLIP following reovirus infection sensitizes human ovarian cancer cells to TRAIL-induced apoptosis. Apoptosis 2007, 12, 211–223. [Google Scholar] [CrossRef]
- Kilani, R.T.; Tamimi, Y.; Hanel, E.G.; Wong, K.K.; Karmali, S.; Lee, P.W.K.; Moore, R.B. Selective reovirus killing of bladder cancer in a co-culture spheroid model. Virus Res. 2003, 93, 1–12. [Google Scholar] [CrossRef]
- Samson, A.; Bentham, M.J.; Scott, K.; Nuovo, G.; Bloy, A.; Appleton, E.; Adair, R.A.; Dave, R.; Peckham-Cooper, A.; Toogood, G.; et al. Oncolytic reovirus as a combined antiviral and anti-tumour agent for the treatment of liver cancer. Gut 2018, 67, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Thirukkumaran, C.M.; Nodwell, M.J.; Hirasawa, K.; Shi, Z.-Q.; Diaz, R.; Luider, J.; Johnston, R.N.; Forsyth, P.A.; Magliocco, A.M.; Lee, P.; et al. Oncolytic viral therapy for prostate cancer: Efficacy of reovirus as a biological therapeutic. Cancer Res. 2010, 70, 2435–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahalingam, D.; Fountzilas, C.; Moseley, J.; Noronha, N.; Tran, H.; Chakrabarty, R.; Selvaggi, G.; Coffey, M.; Thompson, B.; Sarantopoulos, J. A phase II study of REOLYSIN(®) (pelareorep) in combination with carboplatin and paclitaxel for patients with advanced malignant melanoma. Cancer Chemother. Pharm. Ther. 2017, 79, 697–703. [Google Scholar] [CrossRef]
- Roulstone, V.; Twigger, K.; Zaidi, S.; Pencavel, T.; Kyula, J.N.; White, C.; McLaughlin, M.; Seth, R.; Karapanagiotou, E.M.; Mansfield, D.; et al. Synergistic cytotoxicity of oncolytic reovirus in combination with cisplatin-paclitaxel doublet chemotherapy. Gene Ther. 2013, 20, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Karapanagiotou, E.M.; Roulstone, V.; Twigger, K.; Ball, M.; Tanay, M.; Nutting, C.; Newbold, K.; Gore, M.E.; Larkin, J.; Syrigos, K.N.; et al. Phase I/II trial of carboplatin and paclitaxel chemotherapy in combination with intravenous oncolytic reovirus in patients with advanced malignancies. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 2080–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lolkema, M.P.; Arkenau, H.-T.; Harrington, K.; Roxburgh, P.; Morrison, R.; Roulstone, V.; Twigger, K.; Coffey, M.; Mettinger, K.; Gill, G.; et al. A phase I study of the combination of intravenous reovirus type 3 Dearing and gemcitabine in patients with advanced cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Gao, L.; Yeagy, B.; Reid, T. Virus combinations and chemotherapy for the treatment of human cancers. Curr. Opin. Mol. Ther. 2008, 10, 371–379. [Google Scholar] [PubMed]
- LaRocca, C.J.; Warner, S.G. Oncolytic viruses and checkpoint inhibitors: Combination therapy in clinical trials. Clin. Transl. Med. 2018, 7, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrington, K.J.; Vile, R.G.; Melcher, A.; Chester, J.; Pandha, H.S. Clinical trials with oncolytic reovirus: Moving beyond phase I into combinations with standard therapeutics. Cytokine Growth Factor Rev. 2010, 21, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Sampath, P.; Thorne, S.H. Novel therapeutic strategies in human malignancy: Combining immunotherapy and oncolytic virotherapy. Oncolytic Virother. 2015, 4, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Jonker, D.J.; Tang, P.A.; Kennecke, H.; Welch, S.A.; Cripps, M.C.; Asmis, T.; Chalchal, H.; Tomiak, A.; Lim, H.; Ko, Y.-J.; et al. A Randomized Phase II Study of FOLFOX6/Bevacizumab With or Without Pelareorep in Patients With Metastatic Colorectal Cancer: IND.210, a Canadian Cancer Trials Group Trial. Clin. Colorectal Cancer 2018, 17, 231–239.e7. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, D.; Wilkinson, G.A.; Eng, K.H.; Fields, P.; Raber, P.; Moseley, J.L.; Cheetham, K.; Coffey, M.; Nuovo, G.; Kalinski, P.; et al. Pembrolizumab in Combination with the Oncolytic Virus Pelareorep and Chemotherapy in Patients with Advanced Pancreatic Adenocarcinoma: A Phase Ib Study. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020, 26, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, A.A.; Reid, C.; Crawford, M.; Cheetham, K.; Dzugalo, A.; Parsi, M.; Penman, A.; Noronha, N.; Galindez, D.; O’Flynn, R.; et al. Pooled data analysis of the safety and tolerability of intravenous pelareorep in combination with chemotherapy in 500 + cancer patients. Ann. Oncol. 2017, 28, v422. [Google Scholar] [CrossRef] [Green Version]
- Mehr, S. Onco’Zine Editorial Team Positive Top Line Data Presented for Reolysin? in Randomized Phase III Study in Head and Neck Cancers. Available online: https://www.oncozine.com/positive-top-line-data-presented-for-reolysin-in-randomized-phase-iii-study-in-head-and-neck-cancers/ (accessed on 12 May 2021).
- Falese, L.; Sandza, K.; Yates, B.; Triffault, S.; Gangar, S.; Long, B.; Tsuruda, L.; Carter, B.; Vettermann, C.; Zoog, S.J.; et al. Strategy to detect pre-existing immunity to AAV gene therapy. Gene Ther. 2017, 24, 768–778. [Google Scholar] [CrossRef] [Green Version]
- Fausther-Bovendo, H.; Kobinger, G.P. Pre-existing immunity against Ad vectors: Humoral, cellular, and innate response, what’s important? Hum. Vaccines Immunother. 2014, 10, 2875–2884. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xing, M.; Zhang, C.; Yang, Y.; Chi, Y.; Tang, X.; Zhang, H.; Xiong, S.; Yu, L.; Zhou, D. Neutralizing antibody responses to enterovirus and adenovirus in healthy adults in China. Emerg. Microbes Infect. 2014, 3, e30. [Google Scholar] [CrossRef]
- Ricca, J.M.; Oseledchyk, A.; Walther, T.; Liu, C.; Mangarin, L.; Merghoub, T.; Wolchok, J.D.; Zamarin, D. Pre-existing Immunity to Oncolytic Virus Potentiates Its Immunotherapeutic Efficacy. Mol. Ther. 2018, 26, 1008–1019. [Google Scholar] [CrossRef] [Green Version]
- Tähtinen, S.; Feola, S.; Capasso, C.; Laustio, N.; Groeneveldt, C.; Ylösmäki, E.O.; Ylösmäki, L.; Martins, B.; Fusciello, M.; Medeot, M.; et al. Exploiting Preexisting Immunity to Enhance Oncolytic Cancer Immunotherapy. Cancer Res. 2020, 80, 2575–2585. [Google Scholar] [CrossRef] [Green Version]
- Niemann, J.; Woller, N.; Brooks, J.; Fleischmann-Mundt, B.; Martin, N.T.; Kloos, A.; Knocke, S.; Ernst, A.M.; Manns, M.P.; Kubicka, S.; et al. Molecular retargeting of antibodies converts immune defense against oncolytic viruses into cancer immunotherapy. Nat. Commun. 2019, 10, 3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, H.L.; Bommareddy, P.K. Two roads for oncolytic immunotherapy development. J. Immunother. Cancer 2019, 7, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, J.R.; Bedford, R.; Abbruzzese, J.L.; Lahoti, S.; Reid, T.R.; Soetikno, R.M.; Kirn, D.H.; Freeman, S.M. A phase I/II trial of intratumoral endoscopic ultrasound injection of ONYX-015 with intravenous gemcitabine in unresectable pancreatic carcinoma. Clin. Cancer Res. 2003, 9, 555–561. [Google Scholar]
- Khuri, F.R.; Nemunaitis, J.; Ganly, I.; Arseneau, J.; Tannock, I.F.; Romel, L.; Gore, M.; Ironside, J.; MacDougall, R.H.; Heise, C.; et al. A controlled trial of intratumoral ONYX-015, a selectively-replicating adenovirus, in combination with cisplatin and 5-fluorouracil in patients with recurrent head and neck cancer. Nat. Med. 2000, 6, 879–885. [Google Scholar] [CrossRef] [PubMed]
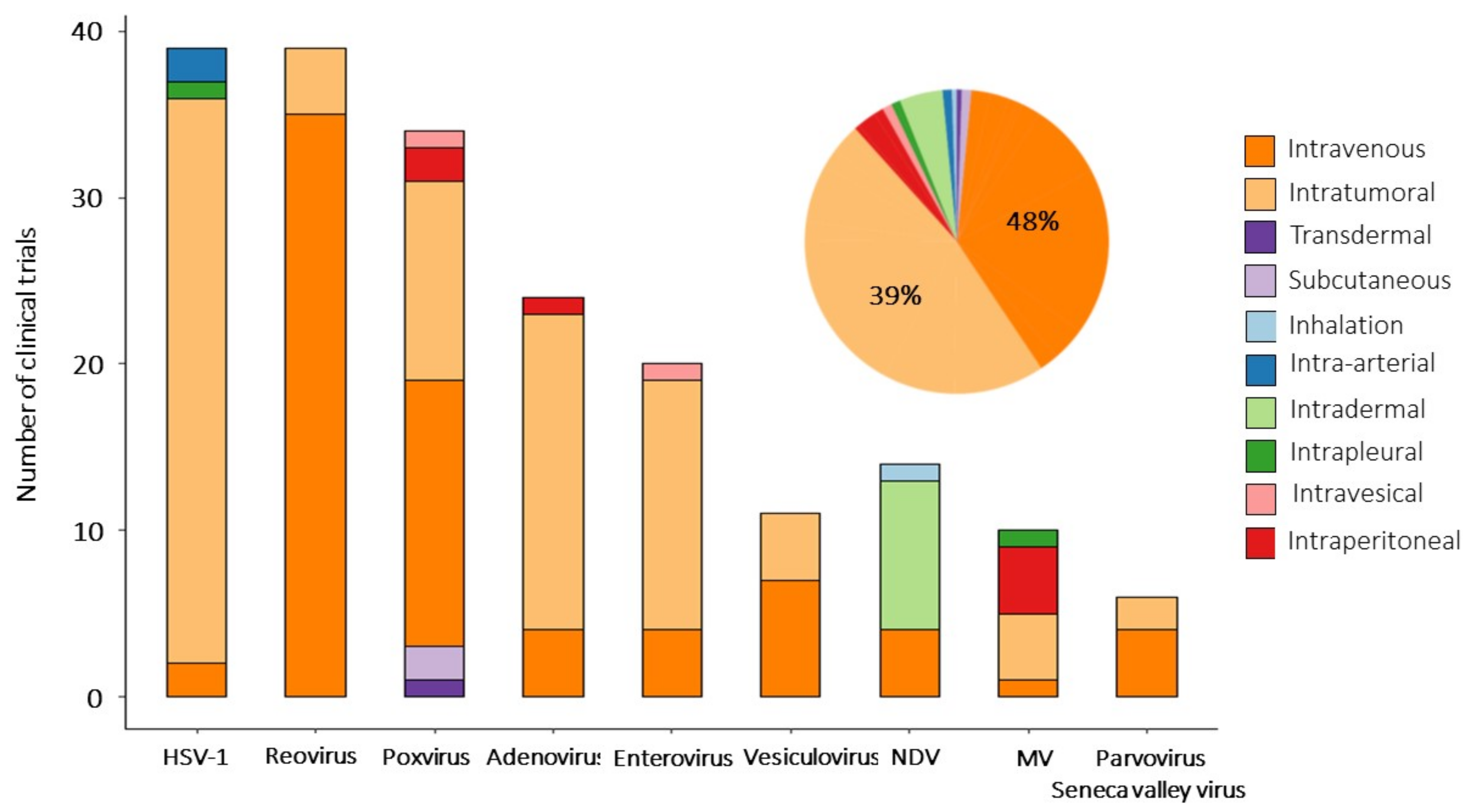
- Hill, C.; Carlisle, R. Achieving systemic delivery of oncolytic viruses. Expert Opin. Drug Deliv. 2019, 16, 607–620. [Google Scholar] [CrossRef] [Green Version]
- Wirsching, H.-G.; Zhang, H.; Szulzewsky, F.; Arora, S.; Grandi, P.; Cimino, P.J.; Amankulor, N.; Campbell, J.S.; McFerrin, L.; Pattwell, S.S.; et al. Arming oHSV with ULBP3 drives abscopal immunity in lymphocyte-depleted glioblastoma. Jci Insight 2019, 4, e128217. [Google Scholar] [CrossRef]
- Kuryk, L.; Møller, A.-S.W.; Jaderberg, M. Abscopal effect when combining oncolytic adenovirus and checkpoint inhibitor in a humanized NOG mouse model of melanoma. J. Med. Virol. 2019, 91, 1702–1706. [Google Scholar] [CrossRef] [Green Version]
- Havunen, R.; Santos, J.M.; Sorsa, S.; Rantapero, T.; Lumen, D.; Siurala, M.; Airaksinen, A.J.; Cervera-Carrascon, V.; Tähtinen, S.; Kanerva, A.; et al. Abscopal Effect in Non-injected Tumors Achieved with Cytokine-Armed Oncolytic Adenovirus. Mol. Ther. Oncolytics 2018, 11, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, P.; Li, H.; Du, X.; Liu, M.; Huang, Q.; Wang, Y.; Wang, S. The Efficacy of Oncolytic Adenovirus Is Mediated by T-cell Responses against Virus and Tumor in Syrian Hamster Model. Clin. Cancer Res. Ther. Off. J. Am. Assoc. Cancer Res. 2017, 23, 239–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alemany, R.; Suzuki, K.; Curiel, D.T. Blood clearance rates of adenovirus type 5 in mice. J. Gen. Virol. 2000, 81, 2605–2609. [Google Scholar] [CrossRef]
- Tesfay, M.Z.; Ammayappan, A.; Federspiel, M.J.; Barber, G.N.; Stojdl, D.; Peng, K.-W.; Russell, S.J. Vesiculovirus neutralization by natural IgM and complement. J. Virol. 2014, 88, 6148–6157. [Google Scholar] [CrossRef] [Green Version]
- Jain, R.K.; Stylianopoulos, T. Delivering nanomedicine to solid tumors. Nat. Rev. Clin. Oncol. 2010, 7, 653–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, Y.; Murai, C.; Shibata, S.; Munakata, Y.; Ishii, T.; Ishii, K.; Saitoh, T.; Sawai, T.; Sugamura, K.; Sasaki, T. Human parvovirus B19 as a causative agent for rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 1998, 95, 8227–8232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sioofy-Khojine, A.B.; Lehtonen, J.; Nurminen, N.; Laitinen, O.H.; Oikarinen, S.; Huhtala, H.; Pakkanen, O.; Ruokoranta, T.; Hankaniemi, M.M.; Toppari, J.; et al. Coxsackievirus B1 infections are associated with the initiation of insulin-driven autoimmunity that progresses to type 1 diabetes. Diabetologia 2018, 61, 1193–1202. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Xu, L.; Yi, M.; Yu, S.; Wu, K.; Luo, S. Novel immune checkpoint targets: Moving beyond PD-1 and CTLA-4. Mol. Cancer 2019, 18, 155. [Google Scholar] [CrossRef] [PubMed]
- Rujas, E.; Cui, H.; Sicard, T.; Semesi, A.; Julien, J.-P. Structural characterization of the ICOS/ICOS-L immune complex reveals high molecular mimicry by therapeutic antibodies. Nat. Commun. 2020, 11, 5066. [Google Scholar] [CrossRef] [PubMed]
- Solinas, C.; Gu-Trantien, C.; Willard-Gallo, K. The rationale behind targeting the ICOS-ICOS ligand costimulatory pathway in cancer immunotherapy. ESMO Open 2020, 5, e000544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francisco-Cruz, A.; Aguilar-Santelises, M.; Ramos-Espinosa, O.; Mata-Espinosa, D.; Marquina-Castillo, B.; Barrios-Payan, J.; Hernandez-Pando, R. Granulocyte–macrophage colony-stimulating factor: Not just another haematopoietic growth factor. Med. Oncol. 2014, 31, 774. [Google Scholar] [CrossRef]
- Bramante, S.; Koski, A.; Kipar, A.; Diaconu, I.; Liikanen, I.; Hemminki, O.; Vassilev, L.; Parviainen, S.; Cerullo, V.; Pesonen, S.K.; et al. Serotype chimeric oncolytic adenovirus coding for GM-CSF for treatment of sarcoma in rodents and humans. Int. J. Cancer 2014, 135, 720–730. [Google Scholar] [CrossRef]
- Andtbacka, R.H.I.; Kaufman, H.L.; Collichio, F.; Amatruda, T.; Senzer, N.; Chesney, J.; Delman, K.A.; Spitler, L.E.; Puzanov, I.; Agarwala, S.S.; et al. Talimogene Laherparepvec Improves Durable Response Rate in Patients with Advanced Melanoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2015, 33, 2780–2788. [Google Scholar] [CrossRef]
- Lambright, E.S.; Caparrelli, D.J.; Abbas, A.E.; Toyoizumi, T.; Coukos, G.; Molnar-Kimber, K.L.; Kaiser, L.R. Oncolytic therapy using a mutant type-1 herpes simplex virus and the role of the immune system. Ann. Thorac. Surg. 1999, 68, 1752–1756. [Google Scholar] [CrossRef]
- Pol, J.G.; Acuna, S.A.; Yadollahi, B.; Tang, N.; Stephenson, K.B.; Atherton, M.J.; Hanwell, D.; El-Warrak, A.; Goldstein, A.; Moloo, B.; et al. Preclinical evaluation of a MAGE-A3 vaccination utilizing the oncolytic Maraba virus currently in first-in-human trials. Oncoimmunology 2019, 8, e1512329. [Google Scholar] [CrossRef] [Green Version]
- Aznar, M.A.; Molina, C.; Teijeira, A.; Rodriguez, I.; Azpilikueta, A.; Garasa, S.; Sanchez-Paulete, A.R.; Cordeiro, L.; Etxeberria, I.; Alvarez, M.; et al. Repurposing the yellow fever vaccine for intratumoral immunotherapy. Embo Mol. Med. 2020, 12, e10375. [Google Scholar] [CrossRef] [PubMed]
- Kaid, C.; Madi, R.A.D.S.; Astray, R.; Goulart, E.; Caires-Junior, L.C.; Mitsugi, T.G.; Moreno, A.C.R.; Castro-Amarante, M.F.; Pereira, L.R.; Porchia, B.F.M.M.; et al. Safety, Tumor Reduction, and Clinical Impact of Zika Virus Injection in Dogs with Advanced-Stage Brain Tumors. Mol. Ther. 2020, 28, 1276–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero, R.A.; Guerrero, C.A.; Guzmán, F.; Acosta, O. Assessing the oncolytic potential of rotavirus on mouse myeloma cell line Sp2/0-Ag14. Biomedica 2020, 40, 362–381. [Google Scholar] [CrossRef]
- Melero, I.; Gato, M.; Shekarian, T.; Aznar, A.; Valsesia-Wittmann, S.; Caux, C.; Etxeberrria, I.; Teijeira, A.; Marabelle, A. Repurposing infectious disease vaccines for intratumoral immunotherapy. J. Immunother. Cancer 2020, 8, e000443. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Combinatorial Agent | n | Function |
|---|---|---|
| Check-point inhibitors | ||
| Pembrolizumab | 14 | Targets and blocks a PD-1 protein on the surface of T-cells. Blocking PD-1 triggers the T-cells activation towards finding and killing cancer cells. Pemrolizumab known under brand name Keytruda. |
| Nivolumab | 8 | An anti-PD-1 monoclonal antibody (brand name Opdivo). |
| Ipililumab | 5 | Humanized immune checkpoint inhibitor which blocks CTLA-4 receptor and upregulates cytotoxic T–lymphocytes (brand name Yervoy). |
| Avelumab | 3 | An anti-PD-1 monoclonal antibody (brand name Bavencio). |
| Durvalumab | 3 | An anti-PD-L1 specific human IgG1 kappa monoclonal antibody. |
| Cemiplimab | 3 | An anti-PD-1 monoclonal antibody (brand name Libtayo). |
| Atezolizumab | 2 | An anti-PDL-1 monoclonal antibody (brand name Tecentriq). |
| Socazolimab | 1 | Anti-PD-L1 monoclonal antibody (ZKAB001). |
| HX008 | 1 | Anti-PD-L1 monoclonal antibody. |
| Vibostolimab | 1 | Vibostolimab is a monoclonal antibody against T-cell immunoreceptor with Ig and ITIM domains (TIGIT). Vibostolimab blocks the interaction between TIGIT and its ligands (CD112 and CD155) thereby activating T cells. |
| Bevacizumab | 1 | Bevacizumab (Avastin) targets cellular vascular endothelial growth factor (VEGF), a protein that is essential for blood vessel growth. |
| Trasuzumab | 1 | Trasuzumab (Herceptin) is an anti-HER2 monoclonal antibody targeting breast cancer and stomach cancer cells expressing HER2 receptors. |
| Tremelimumab | 1 | An anti- CTLA-4 monoclonal antibody. |
| Immunomodulatory factors | ||
| Interleukine-2 (IL-2) | 3 | Stimulates cytotoxic T cells (CD8+) and NK cells, controls both the primary and secondary expansion of antigen-specific CD8+ T cell populations. |
| Interferon-α | 3 | Cytokine that activates immune cells (NK cells and T-cells) and suppresses tumor cell division by inhibiting protein and hormone synthesis. It also reduces angiogenesis through inhibition of angiogenic factors b-FGF and VEGF. |
| Granulocyte-macrophage colony-stimulating factor (GM-CSF) | 3 | GM-CSF enhances the number of circulating white blood cells and increases neutrophil and monocyte function. It also actively shapes the dendritic cell profile leading to enhanced anti-tumor effect. |
| Antigens | ||
| Autologous tumor cells | 14 | Therapeutic agent produced from patient tumor cells. Processed and treated tumor cells are a great source of cancer antigens that, after administration, boost the immune system of the individual that they have been isolated. |
| Melanoma-associated antigen 3(MAGE-A3) | 3 | MAGE-A3 is a tumor-specific shared antigen often expressed in lung cancer and melanoma. Immunization with MAGE-A3 tends to stimulate the immune response to cancer, which has been traditionally considered as poorly immunogenic. |
| ag-E6E7 | 1 | Human papillomavirus oncoproteins E6 and E7. Immunization with E6 and E7 antigens improves antitumor immunity against HPV-related tumors and enhances the immunogenicity of dendritic cells. |
| Bacillus Calmette-Guerin (BCG) | 1 | Nontumor antigen initially used as a tuberculosis vaccine. High immunogenic BCG mounts overall immune response that potentially decreases the reoccurrence of cancer. |
| Radiotherapy/Chemotherapy/Surgery | 80 | Various drugs, radiotherapy regimes accompanied by tumor resection (where possible) are in use in combination with virotherapy. The reader may find specific details in decent reviews and supplementary materials. |
| Single-agent virotherapy | 88 | Wild-type viruses attenuated or genetically engineered variants armored with immunomodulatory molecules are frequently used as a monotherapy. Variants of the used genetic modifications of oncolytics (mainly for stimulating the immune system) are shown in Table 2. |
| Payload and Modifications | n | Function |
|---|---|---|
| GM-CSF (CSF2) | 49 | GM-CSF is a growth factor that stimulates differentiation, proliferation and migration of myeloid cells. |
| Thymidine kinase (TK) | 24 | HSV-1 TK is a virulence factor deletion of which attenuates virus, but not essential for virus replication. In addition, TK being used as a suicide gene to specifically target tumor cells. |
| Human sodium iodide symporter (hNIS) | 14 | NIS mediates a transport process of iodide uptake. Overexpression of NIS in cancer cells increases iodide concentration within the cells that benefit from radioiodine therapy. |
| p53 (TP53) | 10 | Tumor protein is a major tumor suppressor factor that acts through the regulation of the cell cycle. p53 is often malfunction in tumor cells. |
| Interferon β (IFN-beta) | 8 | IFN-beta is a cytokine, which has an antiviral and anti-proliferative effect. IFN-beta stimulates innate and adaptive immunity and has confirmed antitumor activity. |
| MAGE-A3 | 3 | Tumor-specific antigen. MAGE-A3 immunization elicits antigen-specific immune response. |
| PSA-TRICOM (B7.1, ICAM-1, LFA-3) | 2 | Prostate-specific antigen (PSA). B7.1, ICAM-1, LFA-3 are T-cell costimulatory molecules. |
| Carcinoembryonic antigen (CEA) | 2 | CEA is a glycoprotein, which rarely found in the blood of adults. Expression of CEA serves as a marker for noninvasive monitoring of virus dissemination in vivo. |
| Interleukine-12 (IL-12) | 1 | IL-12 plays a central role in T-cell and natural killer cell responses, induces the production of interferon-γ (IFN-γ). |
| Fas-c and PPE-1 promoter | 1 | Chimeric death receptor Fas and TNF receptor 1 and modified endothelium-specific pre-proendothelin-1 (PPE-1) promoter delivered by virus vector may trigger apoptosis of endothelial cells and reduce tumor angiogenesis. |
| HPV E6/HPV E7 | 1 | Human papillomavirus oncoproteins. |
| TERT promoter | 1 | Telomerase reverse transcriptase promoter (TERT) is used to attenuate virus replication. |
| Interferon-gamma (IFN-ɣ) | 1 | IFN-ɣ is a cytokine molecule with pronounced cytostatic, pro-apoptotic and immune-stimulating effects. |
| Tyrosinase-related protein (TYRP1) | 1 | TYRP1 is expressed in melanomas and on the surface of melanocytes and is an immunoreactive protein. |
| Anti-CTLA4 | 1 | blocks CTLA-4 receptor and upregulate cytotoxic T –lymphocytes |
| None | 109 | Many wild-type viruses have an oncolytic potential and are frequently used without payload. Attenuated or evolutionary selected viruses also demonstrate strong antitumor effect. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malogolovkin, A.; Gasanov, N.; Egorov, A.; Weener, M.; Ivanov, R.; Karabelsky, A. Combinatorial Approaches for Cancer Treatment Using Oncolytic Viruses: Projecting the Perspectives through Clinical Trials Outcomes. Viruses 2021, 13, 1271. https://doi.org/10.3390/v13071271
Malogolovkin A, Gasanov N, Egorov A, Weener M, Ivanov R, Karabelsky A. Combinatorial Approaches for Cancer Treatment Using Oncolytic Viruses: Projecting the Perspectives through Clinical Trials Outcomes. Viruses. 2021; 13(7):1271. https://doi.org/10.3390/v13071271
Chicago/Turabian StyleMalogolovkin, Alexander, Nizami Gasanov, Alexander Egorov, Marianna Weener, Roman Ivanov, and Alexander Karabelsky. 2021. "Combinatorial Approaches for Cancer Treatment Using Oncolytic Viruses: Projecting the Perspectives through Clinical Trials Outcomes" Viruses 13, no. 7: 1271. https://doi.org/10.3390/v13071271
APA StyleMalogolovkin, A., Gasanov, N., Egorov, A., Weener, M., Ivanov, R., & Karabelsky, A. (2021). Combinatorial Approaches for Cancer Treatment Using Oncolytic Viruses: Projecting the Perspectives through Clinical Trials Outcomes. Viruses, 13(7), 1271. https://doi.org/10.3390/v13071271