
Binding of Nanoparticles Harboring Recombinant Large Surface Protein of Hepatitis B Virus to Scavenger Receptor Class B Type 1



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of BNCs
2.2. Preparation of Liposomes
2.3. Cell Culture
2.4. Fluorescent Labeling of BNCs and Liposomes
2.5. LSM
2.6. BNC-Binding Assays Using Flow Cytometry
2.7. Competitive BNC-Binding Assays
2.8. RNA Sequencing (RNA-Seq)
2.9. SiRNA Treatment
2.10. Expression of SR-B1-GFP and Its Mutants in HEK293T Cells
2.11. BNC Uptake Assays
3. Results
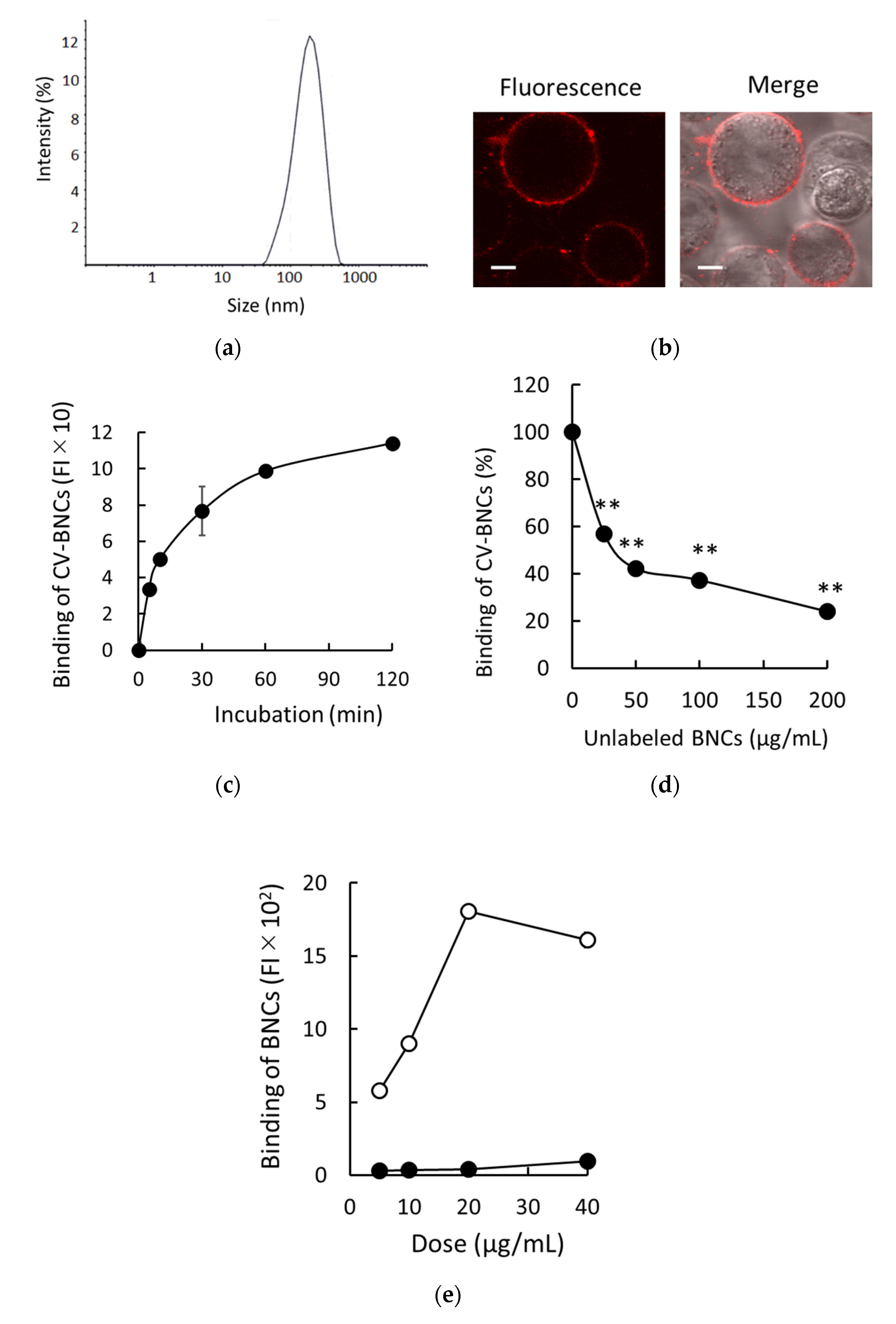
3.1. Binding of BNCs to HEK293T Cells
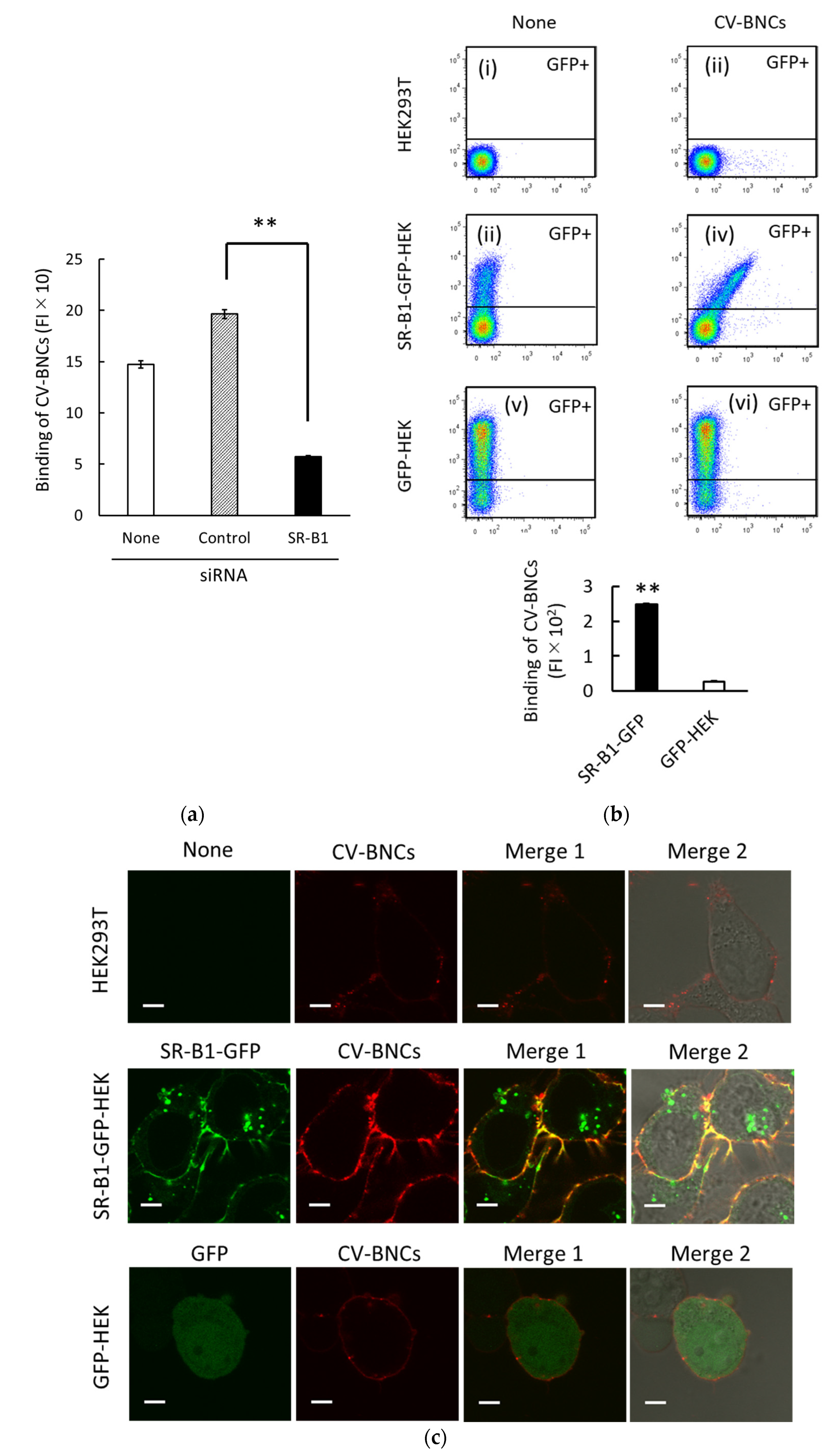
3.2. Binding of BNCs to HEK293T Cells via SR-B1
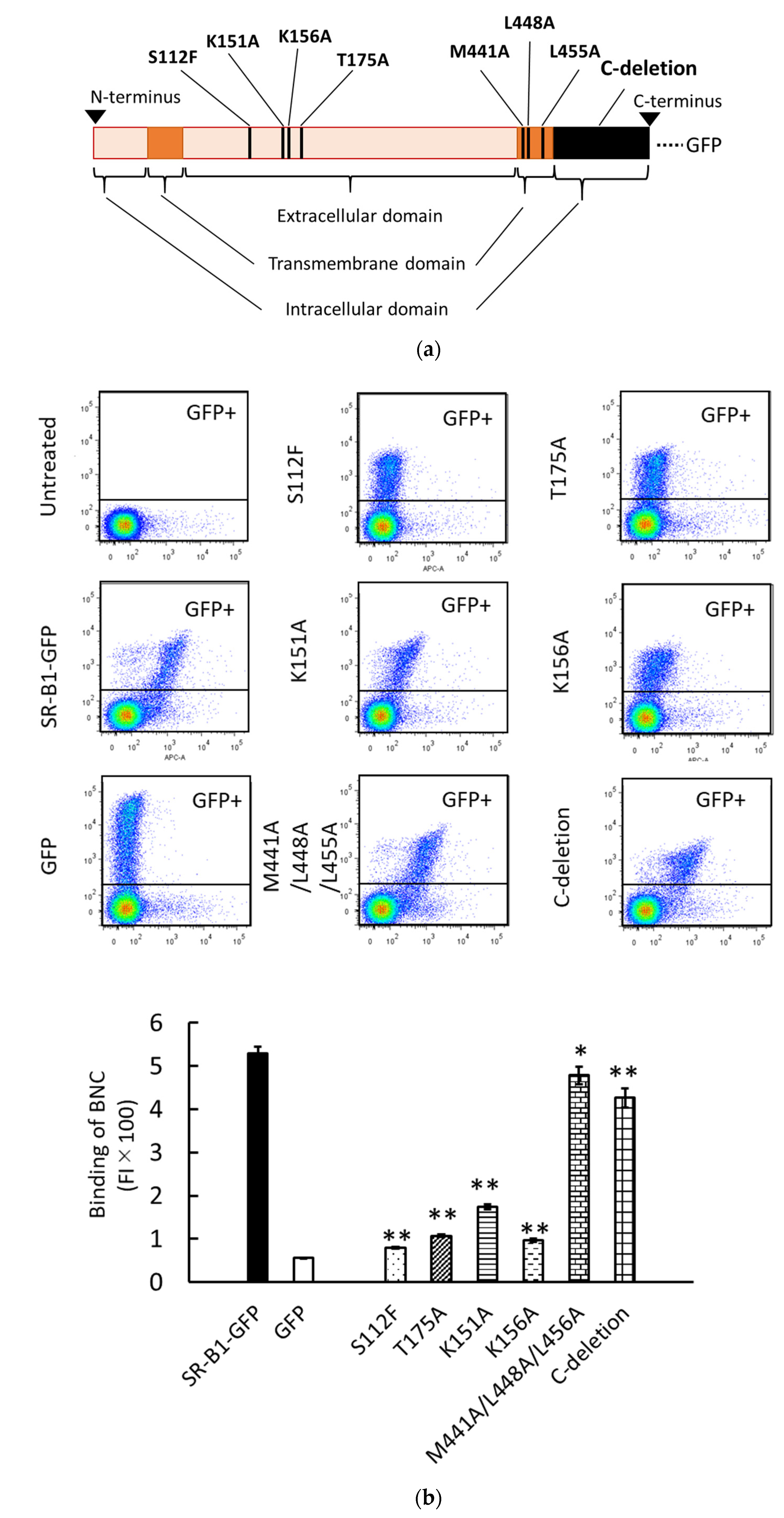
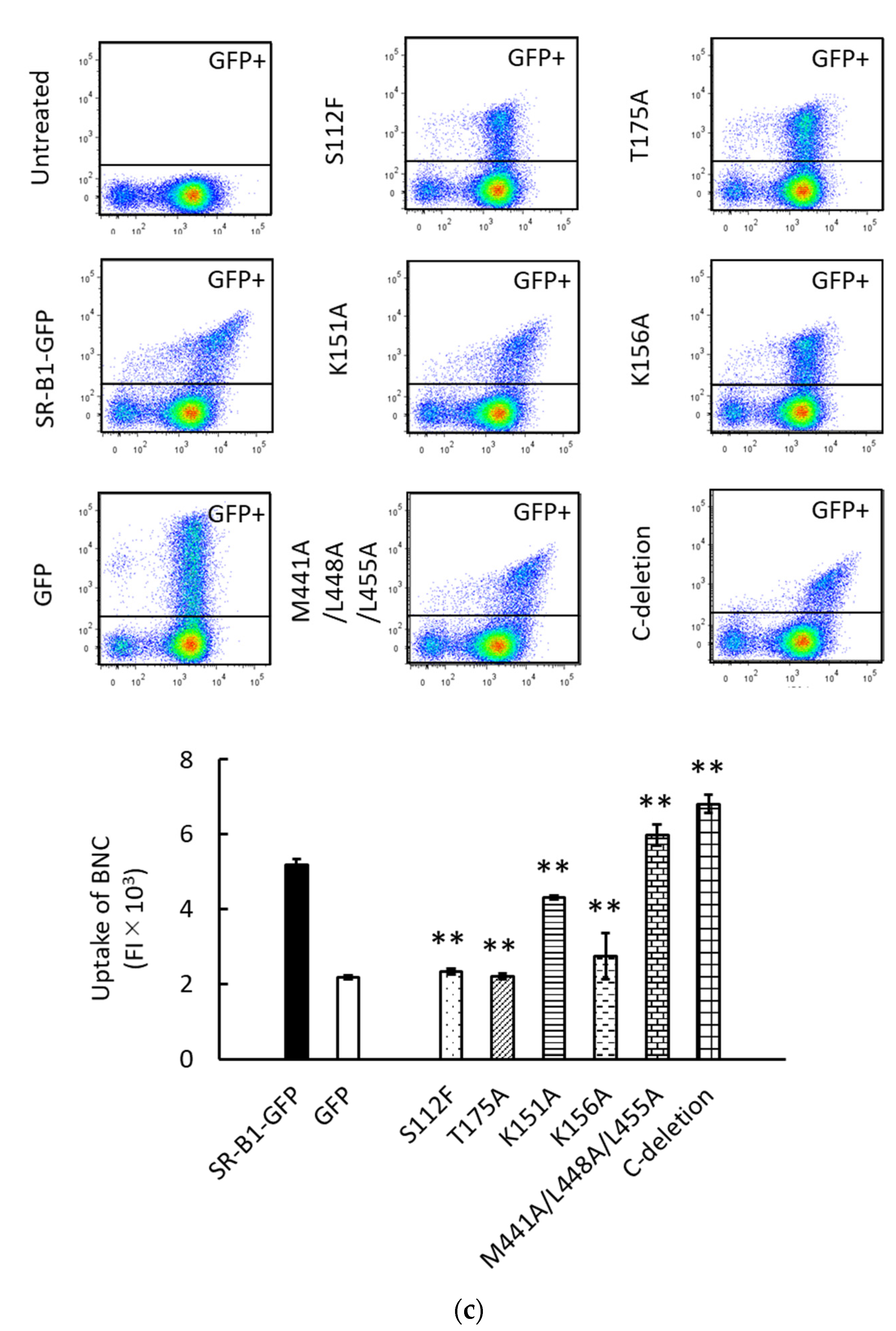
3.3. Binding and Endocytosis of BNCs to HEK293T Cells via SR-B1-GFP Mutants
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heermann, K.H.; Goldmann, U.; Schwartz, W.; Seyffarth, T.; Baumgarten, H.; Gerlich, W.H. Large surface proteins of hepatitis B virus containing the pre-S sequence. J. Virol. 1984, 52, 396–402. [Google Scholar] [CrossRef] [Green Version]
- Block, T.M.; Lu, X.; Platt, F.M.; Foster, G.R.; Gerlich, W.H.; Blumberg, B.S.; Dwek, R.A. Secretion of human hepatitis B virus is inhibited by the imino sugar N-butyldeoxynojirimycin. Proc. Natl. Acad. Sci. USA 1994, 91, 2235–2239. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Amano, C.; Fukuda, T.; Yamada, T.; Kuroda, S.; Tanizawa, K.; Kondo, A.; Ueda, M.; Yamada, H.; Tada, H.; et al. The specific delivery of proteins to human liver cells by engineered bio-nanocapsules. FEBS J. 2005, 272, 3651–3660. [Google Scholar] [CrossRef]
- Yamada, T.; Iwasaki, Y.; Tada, H.; Iwabuki, H.; Chuah, M.K.L.; VandenDriessche, T.; Fukuda, H.; Kondo, A.; Ueda, M.; Seno, M.; et al. Nanoparticles for the delivery of genes and drugs to human hepatocytes. Nat. Biotechnol. 2003, 21, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Somiya, M.; Jeong, S.Y.; Choi, E.K.; Kuroda, S. Low immunogenic bio-nanocapsule based on hepatitis B virus escape mutants. Nanomedicine 2018, 14, 595–600. [Google Scholar] [CrossRef]
- Liu, Q.; Somiya, M.; Kuroda, S. Elucidation of the early infection machinery of hepatitis B virus by using bio-nanocapsule. World J. Gastroenterol. 2016, 22, 8489–8496. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. Elife 2012, 1, e00049. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Watashi, K.; Tsukuda, S.; Aly, H.H.; Fukasawa, M.; Fujimoto, A.; Suzuki, R.; Aizaki, H.; Ito, T.; Koiwai, O.; et al. Evaluation and identification of hepatitis B virus entry inhibitors using HepG2 cells overexpressing a membrane transporter NTCP. Biochem. Biophys. Res. Commun. 2014, 443, 808–813. [Google Scholar] [CrossRef] [Green Version]
- Verrier, E.R.; Colpitts, C.C.; Bach, C.; Heydmann, L.; Weiss, A.; Renaud, M.; Durand, S.C.; Habersetzer, F.; Durantel, D.; Abou-Jaoudé, G.; et al. A targeted functional RNA interference screen uncovers glypican 5 as an entry factor for hepatitis B and D viruses. Hepatology 2016, 63, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Neurath, A.R.; Kent, S.B.; Strick, N.; Parker, K. Identification and chemical synthesis of a host cell receptor binding site on hepatitis B virus. Cell 1986, 46, 429–436. [Google Scholar] [CrossRef]
- Somiya, M.; Liu, Q.; Yoshimoto, N.; Iijima, M.; Tatematsu, K.; Nakai, T.; Okajima, T.; Kuroki, K.; Ueda, K.; Kuroda, S. Cellular uptake of hepatitis B virus envelope L particles is independent of sodium taurocholate cotransporting polypeptide, but dependent on heparan sulfate proteoglycan. Virology 2016, 497, 23–32. [Google Scholar] [CrossRef]
- Fujita, K.; Somiya, M.; Kuroda, S.; Hinuma, S. Induction of lipid droplets in non-macrophage cells as well as macrophages by liposomes and exosomes. Biochem. Biophys. Res. Commun. 2019, 510, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Koide, N.; Somiya, M.; Kuroda, S.; Hinuma, S. A regulatory role of scavenger receptor class B type 1 in endocytosis and lipid droplet formation induced by liposomes containing phosphatidylethanolamine in HEK293T cells. Biochim. Biophys. Acta. Mol. Cell. Res. 2021, 1868, 118859. [Google Scholar] [CrossRef]
- Koide, N.; Fujita, K.; Kuroda, S.; Hinuma, S. Binding of liposomes composed of phosphatidylcholine to scavenger receptor class B type 1 and its modulation by phosphatidic acid in HEK293T cells. Biochim. Biophys. Acta. Mol. Cell. Res. 2021, 1868, 119043. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.J.; Asthana, S.; Kraemer, F.B.; Azhar, S. Scavenger receptor B type 1: Expression, molecular regulation, and cholesterol transport function. J. Lipid Res. 2018, 59, 1114–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanlandschoot, P.; Van Houtte, F.; Hoek, F.; Nieuwland, R.; Leroux-Roels, G. Saccharomyces cerevisiae-derived HBsAg preparations differ in their attachment to monocytes, immune-suppressive potential, and T-cell immunogenicity. J. Med. Virol. 2003, 70, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Meeren, P.V.; Criekinge, W.V.; Vanderdeelen, J.; Baert, L. Phospholipid composition of r-DNA hepatitis B surface antigen. Int. J. Pharm. 1994, 106, 89–92. [Google Scholar] [CrossRef]
- Diminsky, D.; Schirmbeck, R.; Reimann, J.; Barenholz, Y. Comparison between hepatitis B surface antigen (HBsAg) particles derived from mammalian cells (CHO) and yeast cells (Hansenula polymorpha): Composition, structure and immunologenicity. Vaccine 1997, 15, 637–647. [Google Scholar] [CrossRef]
- Jung, J.; Iijima, M.; Yoshimoto, N.; Sasaki, M.; Niimi, T.; Tatematsu, K.; Jeong, S.Y.; Choi, E.K.; Tanizawa, K.; Kuroda, S. Efficient and rapid purification of drug- and gene-carrying bio-nanocapsules, hepatitis B virus surface antigen L particles, from Saccharomyces cerevisiae. Protein Expr. Purif. 2011, 78, 149–155. [Google Scholar] [CrossRef]
- Chadwick, A.C.; Sahoo, D. Functional characterization of newly-discovered mutations in human SR-BI. PLoS ONE 2012, 7, e45660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsugita, M.; Morimoto, N.; Tashiro, M.; Kinoshita, K.; Nakayama, M. SR-B1 is a silica receptor that mediates canonical inflammasome activation. Cell Rep. 2017, 18, 1298–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, P.E.; Nyegaard, S.; Collins, R.F.; Troise, F.; Freeman, S.A.; Trimble, W.S.; Grinstein, S. Multimerization and retention of the scavenger receptor SR-B1 in the plasma membrane. Developmental Cell 2019, 50, 283–295.e5. [Google Scholar] [CrossRef] [PubMed]
- Sahebi, R.; Hassanian, S.M.; Ghayour-Mobarhan, M.; Farrokhi, E.; Rezayi, M.; Samadi, S.; Bahramian, S.; Ferns, G.A.; Avan, A. Scavenger receptor class B type I as a potential risk stratification biomarker and therapeutic target in cardiovascular disease. J. Cell Physiol. 2019, 234, 16925–16932. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, L.P.; Mates, J.M.; Cheplowitz, A.M.; Avila, C.L.; Zimmerer, J.M.; Yao, Z.; Maiseyeu, A.; Rajaram, M.V.; Robinson, J.M.; Anderson, C.L. Scavenger receptor B1, the HDL receptor, is expressed abundantly in liver sinusoidal endothelial cells. Sci. Rep. 2016, 6, 20646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Somiya, M.; Iijima, M.; Tatematsu, K.; Kuroda, S. A hepatitis B virus-derived human hepatic cell-specific heparin-binding peptide: Identification and application to a drug delivery system. Biomater. Sci. 2018, 7, 322–335. [Google Scholar] [CrossRef]
- Nanahara, M.; Chang, Y.-T.; Somiya, S.; Kuroda, S. HBV pre-S1-derived myristoylated pepetide (Myr47): Identification of the inhibitory activity on the cellular uptake of lipid nanoparticles. Viruses 2021, 13, 929. [Google Scholar] [CrossRef]
- Jeulin, H.; Velay, A.; Murray, J.; Schvoerer, E. Clinical impact of hepatitis B and C virus envelope glycoproteins. World J. Gastroenterol. 2013, 19, 654–664. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinuma, S.; Fujita, K.; Kuroda, S. Binding of Nanoparticles Harboring Recombinant Large Surface Protein of Hepatitis B Virus to Scavenger Receptor Class B Type 1. Viruses 2021, 13, 1334. https://doi.org/10.3390/v13071334
Hinuma S, Fujita K, Kuroda S. Binding of Nanoparticles Harboring Recombinant Large Surface Protein of Hepatitis B Virus to Scavenger Receptor Class B Type 1. Viruses. 2021; 13(7):1334. https://doi.org/10.3390/v13071334
Chicago/Turabian StyleHinuma, Shuji, Kazuyo Fujita, and Shun’ichi Kuroda. 2021. "Binding of Nanoparticles Harboring Recombinant Large Surface Protein of Hepatitis B Virus to Scavenger Receptor Class B Type 1" Viruses 13, no. 7: 1334. https://doi.org/10.3390/v13071334
APA StyleHinuma, S., Fujita, K., & Kuroda, S. (2021). Binding of Nanoparticles Harboring Recombinant Large Surface Protein of Hepatitis B Virus to Scavenger Receptor Class B Type 1. Viruses, 13(7), 1334. https://doi.org/10.3390/v13071334