
Membrane Insertion of the M13 Minor Coat Protein G3p Is Dependent on YidC and the SecAYEG Translocase


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Proteinase K Mapping
2.3. Immunoprecipitation
2.4. Disulphide Crosslinking
3. Results
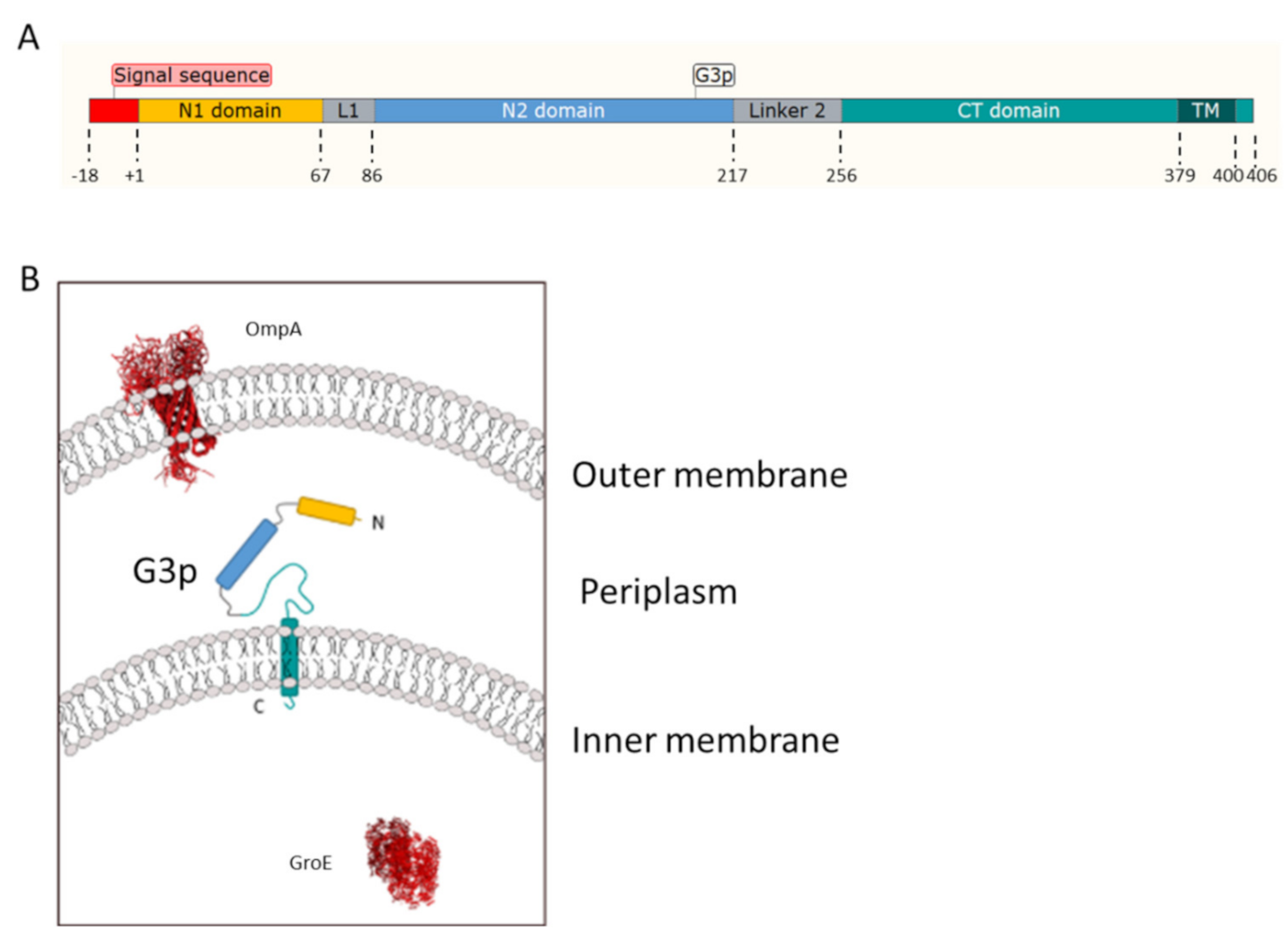
3.1. Membrane Targeting of G3p
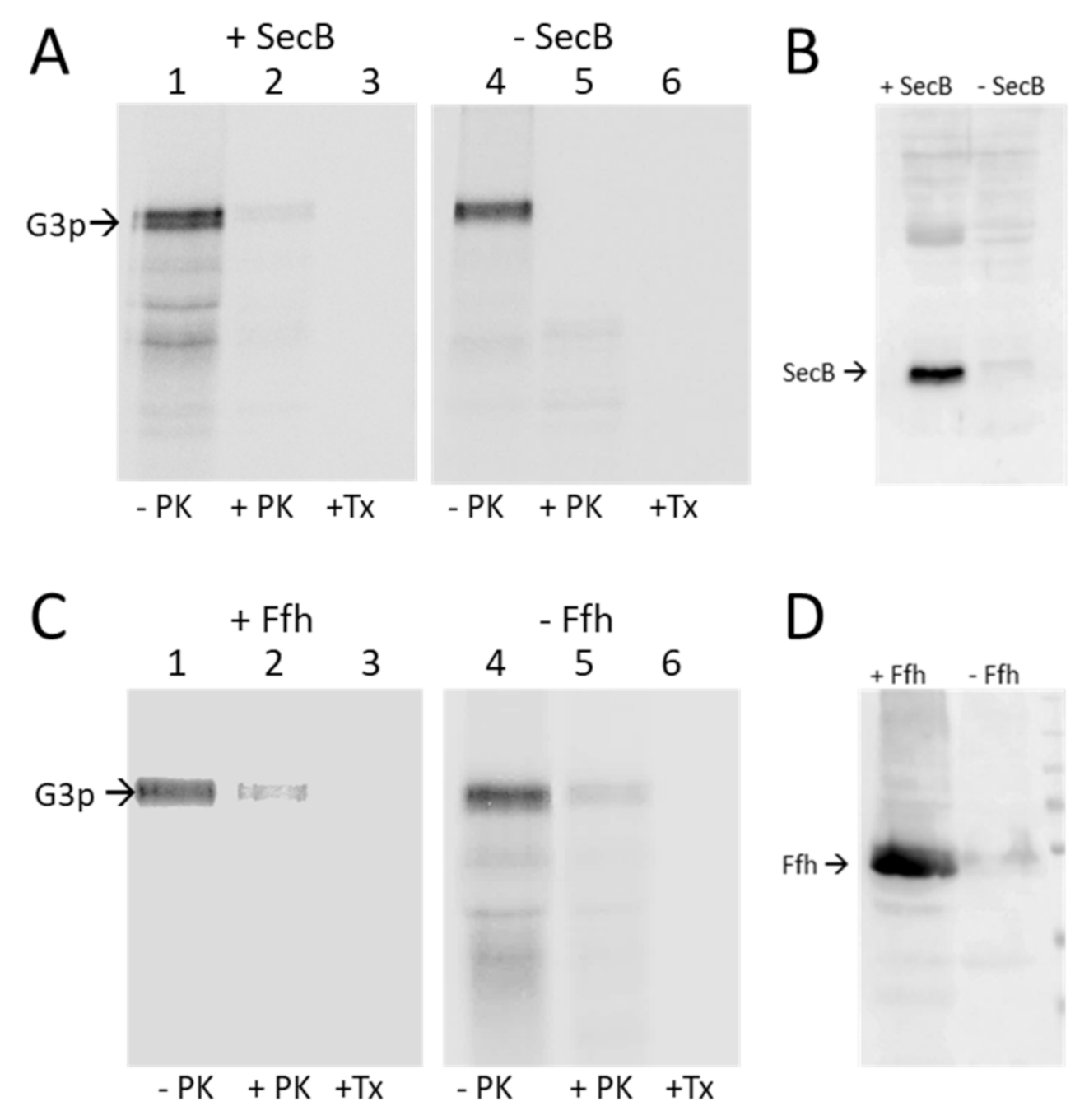
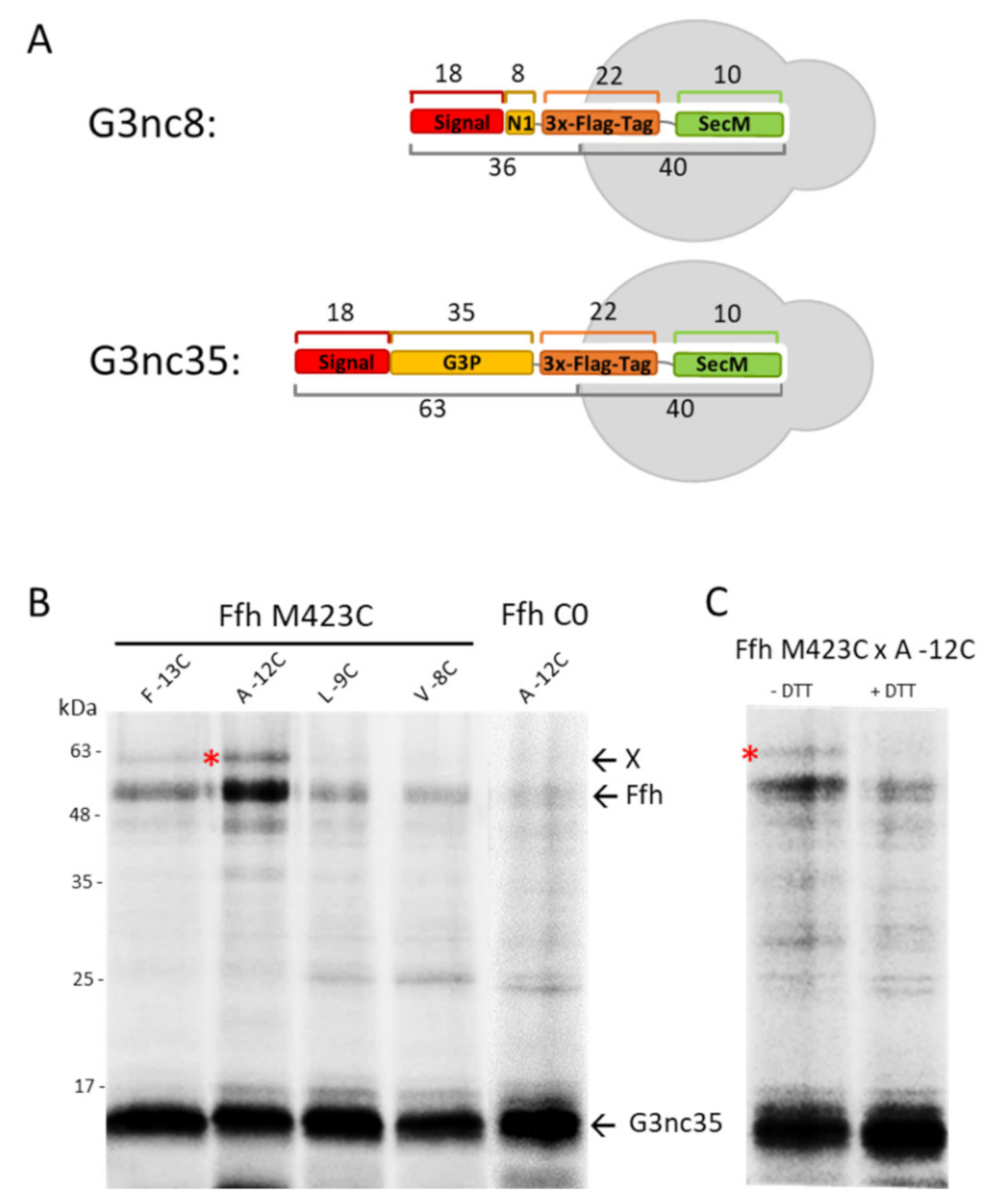
3.2. Binding of G3p Nascent Chains to Ffh
3.3. Membrane Insertion of G3p Requires SecAYEG Translocase and YidC
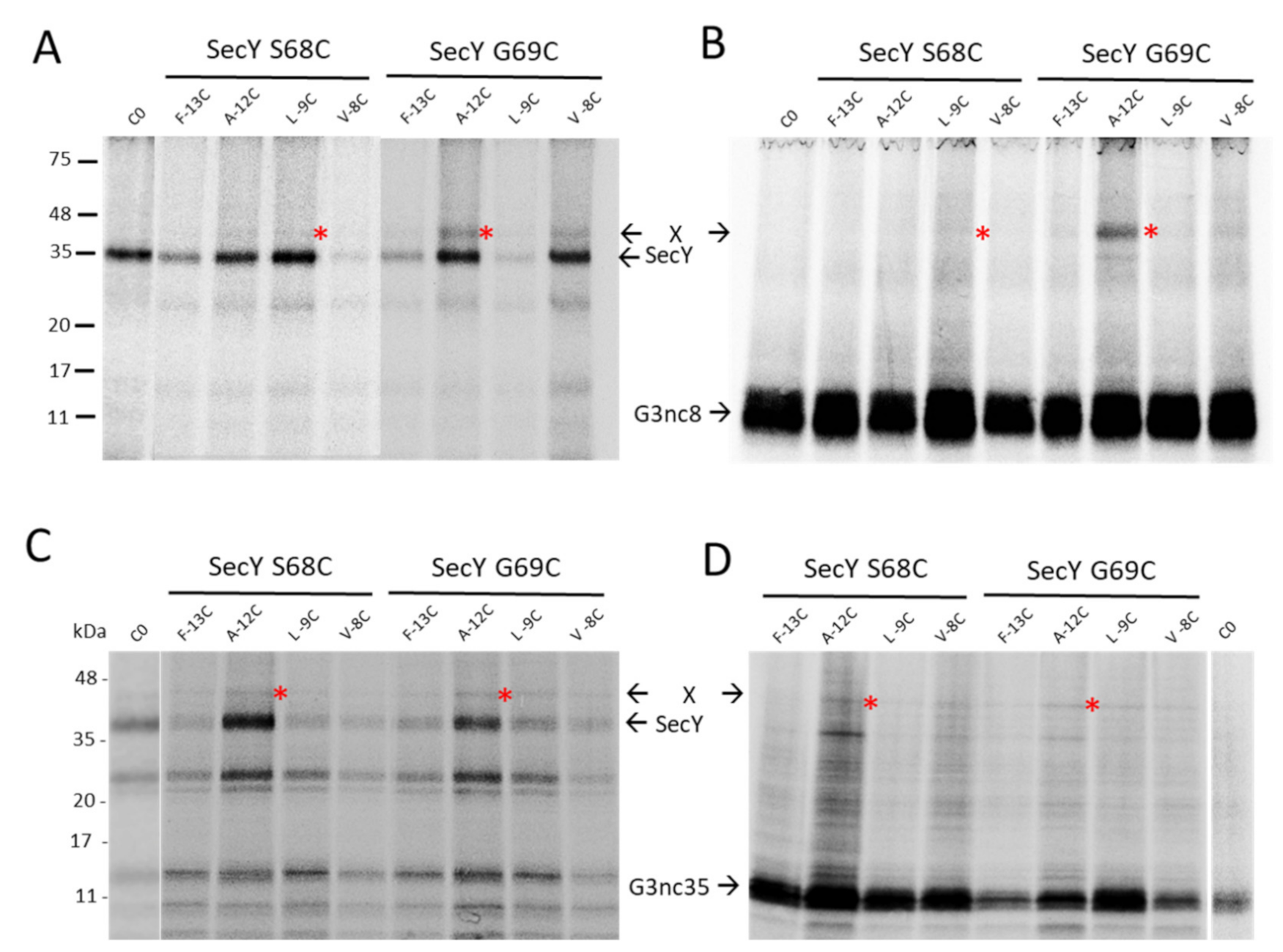
3.4. Binding of G3p Nascent Chains to SecY and YidC
3.5. Binding of the G3p Membrane Anchor to SecY and YidC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Loh, B.; Kuhn, A.; Leptihn, S. The fascinating biology behind phage display: Filamentous phage assembly. Mol. Microbiol. 2021, 111, 1132–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, A.; Schmid, F.X. The Folding Mechanism of a Two-domain Protein: Folding Kinetics and Domain Docking of the Gene-3 Protein of Phage fd. J. Mol. Biol. 2003, 329, 599–610. [Google Scholar] [CrossRef]
- Rakonjac, J.; Feng, J.N.; Model, P. Filamentous phage are released from the bacterial membrane by a two-step mechanism involving a short C-terminal fragment of pIII. J. Mol. Biol. 1999, 289, 1253–1265. [Google Scholar] [CrossRef] [PubMed]
- Boeke, J.D.; Model, P.; Zinder, N.D. Effects of bacteriophage f1 gene III protein on the host cell membrane. Mol. Gen. Genet. 1982, 186, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.; Silveira, N.F.; Fleet, G.H. Occurrence and properties of bacteriophages of Leuconostoc oenos in Australian wines. Appl. Environ. Microbiol. 1985, 50, 872–876. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef]
- Riechmann, L.; Holliger, P. The C-Terminal Domain of TolA Is the Coreceptor for Filamentous Phage Infection of E. coli. Cell 1997, 90, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Lubkowski, J.; Hennecke, F.; Plückthun, A.; Wlodawer, A. Filamentous phage infection: Crystal structure of g3p in complex with its coreceptor, the C-terminal domain of TolA. Structure 1999, 7, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, A.; Koch, H.-G.; Dalbey, R.E. Targeting and Insertion of Membrane Proteins. EcoSal Plus 2017, 7. [Google Scholar] [CrossRef]
- Botte, M.; Zaccai, N.R.; Nijeholt, J.L.À.; Martin, R.; Knoops, K.; Papai, G.; Zou, J.; Deniaud, A.; Karuppasamy, M.; Jiang, Q.; et al. A central cavity within the holo-translocon suggests a mechanism for membrane protein insertion. Sci. Rep. 2016, 6, 38399. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, J.; Nam, Y.; Rapoport, T.A. Structure of a complex of the ATPase SecA and the protein-translocation channel. Nature 2008, 455, 936–943. [Google Scholar] [CrossRef]
- Park, E.; Ménétret, J.-F.; Gumbart, J.C.; Ludtke, S.J.; Li, W.; Whynot, A.; Rapoport, T.A.; Akey, C.W. Structure of the SecY channel during initiation of protein translocation. Nature 2014, 506, 102–106. [Google Scholar] [CrossRef]
- Tanaka, Y.; Izumioka, A.; Abdul Hamid, A.; Fujii, A.; Haruyama, T.; Furukawa, A.; Tsukazaki, T. 2.8-Å crystal structure of Escherichia coli YidC revealing all core regions, including flexible C2 loop. Biochem. Biophys. Res. Commun. 2018, 505, 141–145. [Google Scholar] [CrossRef]
- Li, L.; Park, E.; Ling, J.; Ingram, J.; Ploegh, H.; Rapoport, T.A. Crystal structure of a substrate-engaged SecY protein-translocation channel. Nature 2016, 531, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Park, E.; Rapoport, T.A. Preserving the membrane barrier for small molecules during bacterial protein translocation. Nature 2011, 473, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Cannon, K.S.; Or, E.; Clemons, W.M.; Shibata, Y.; Rapoport, T.A. Disulfide bridge formation between SecY and a translocating polypeptide localizes the translocation pore to the center of SecY. J. Cell Biol. 2005, 169, 219–225. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Kuhn, A.; Dalbey, R.E. Tracking the Stepwise Movement of a Membrane-inserting Protein In Vivo. J. Mol. Biol. 2020, 432, 484–496. [Google Scholar] [CrossRef]
- Geng, Y.; Kedrov, A.; Caumanns, J.J.; Crevenna, A.H.; Lamb, D.C.; Beckmann, R.; Driessen, A.J.M. Role of the Cytosolic Loop C2 and the C Terminus of YidC in Ribosome Binding and Insertion Activity. J. Biol. Chem. 2015, 290, 17250–17261. [Google Scholar] [CrossRef] [Green Version]
- Kedrov, A.; Wickles, S.; Crevenna, A.H.; van der Sluis, E.O.; Buschauer, R.; Berninghausen, O.; Lamb, D.C.; Beckmann, R. Structural Dynamics of the YidC:Ribosome Complex during Membrane Protein Biogenesis. Cell Rep. 2016, 17, 2943–2954. [Google Scholar] [CrossRef] [Green Version]
- Klenner, C.; Yuan, J.; Dalbey, R.E.; Kuhn, A. The Pf3 coat protein contacts TM1 and TM3 of YidC during membrane biogenesis. FEBS Lett. 2008, 582, 3967–3972. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, F.; Fuchs, R.T.; Storz, G. Membrane localization of small proteins in Escherichia coli. J. Biol. Chem. 2011, 286, 32464–32474. [Google Scholar] [CrossRef] [Green Version]
- Steudle, A.; Spann, D.; Pross, E.; Shanmugam, S.K.; Dalbey, R.E.; Kuhn, A. Molecular communication of the membrane insertase YidC with translocase SecYEG affects client proteins. Sci. Rep. 2021, 11, 3940. [Google Scholar] [CrossRef]
- Seitl, I.; Wickles, S.; Beckmann, R.; Kuhn, A.; Kiefer, D. The C-terminal regions of YidC from Rhodopirellula baltica and Oceanicaulis alexandrii bind to ribosomes and partially substitute for SRP receptor function in Escherichia coli. Mol. Microbiol. 2014, 91, 408–421. [Google Scholar] [CrossRef]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef] [Green Version]
- Casadaban, M.J.; Cohen, S.N. Analysis of gene control signals by DNA fusion and cloning in Escherichia coli. J. Mol. Biol. 1980, 138, 179–207. [Google Scholar] [CrossRef]
- Lessl, M.; Balzer, D.; Lurz, R.; Waters, V.L.; Guiney, D.G.; Lanka, E. Dissection of IncP conjugative plasmid transfer: Definition of the transfer region Tra2 by mobilization of the Tra1 region in trans. J. Bacteriol. 1992, 174, 2493–2500. [Google Scholar] [CrossRef] [Green Version]
- Pross, E.; Kuhn, A. The SRP signal sequence of KdpD. Sci. Rep. 2019, 9, 8717. [Google Scholar] [CrossRef]
- Pross, E.; Kuhn, A. Two Signal Recognition Particle Sequences Are Present in the Amino-Terminal Domain of the C-Tailed Protein SciP. J. Bacteriol. 2020, 203. [Google Scholar] [CrossRef]
- Oliver, D.B.; Cabelli, R.J.; Dolan, K.M.; Jarosik, G.P. Azide-resistant mutants of Escherichia coli alter the SecA protein, an azide-sensitive component of the protein export machinery. Proc. Natl. Acad. Sci. USA 1990, 87, 8227–8231. [Google Scholar] [CrossRef] [Green Version]
- Noriega, T.R.; Chen, J.; Walter, P.; Puglisi, J.D. Real-time observation of signal recognition particle binding to actively translating ribosomes. Elife 2014, 3. [Google Scholar] [CrossRef]
- Ariosa, A.; Lee, J.H.; Wang, S.; Saraogi, I.; Shan, S. Regulation by a chaperone improves substrate selectivity during cotranslational protein targeting. Proc. Natl. Acad. Sci. USA 2015, 112, E3169–E3178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodbury, R.L.; Topping, T.B.; Diamond, D.L.; Suciu, D.; Kumamoto, C.A.; Hardy, S.J.; Randall, L.L. Complexes between protein export chaperone SecB and SecA. Evidence for separate sites on SecA providing binding energy and regulatory interactions. J. Biol. Chem. 2000, 275, 24191–24198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Xu, Z. Structural determinants of SecB recognition by SecA in bacterial protein translocation. Nat. Struct. Biol. 2003, 10, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Suo, Y.; Hardy, S.J.S.; Randall, L.L. The basis of asymmetry in the SecA:SecB complex. J. Mol. Biol. 2015, 427, 887–900. [Google Scholar] [CrossRef] [Green Version]
- Cymer, F.; Hedman, R.; Ismail, N.; von Heijne, G. Exploration of the arrest peptide sequence space reveals arrest-enhanced variants. J. Biol. Chem. 2015, 290, 10208–10215. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.D.; Bernstein, H.D.; Walter, P. Interaction of E. coli Ffh/4.5S ribonucleoprotein and FtsY mimics that of mammalian signal recognition particle and its receptor. Nature 1994, 367, 657–659. [Google Scholar] [CrossRef]
- Kuhn, A. Alterations in the extracellular domain of M13 procoat protein make its membrane insertion dependent on secA and secY. Eur. J. Biochem. 1988, 177, 267–271. [Google Scholar] [CrossRef]
- Roos, T.; Kiefer, D.; Hugenschmidt, S.; Economou, A.; Kuhn, A. Indecisive M13 procoat protein mutants bind to SecA but do not activate the translocation ATPase. J. Biol. Chem. 2001, 276, 37909–37915. [Google Scholar] [CrossRef]
- Brundage, L.; Hendrick, J.P.; Schiebel, E.; Driessen, A.J.; Wickner, W. The purified E. coli integral membrane protein SecYE is sufficient for reconstitution of SecA-dependent precursor protein translocation. Cell 1990, 62, 649–657. [Google Scholar] [CrossRef] [Green Version]
- Osborne, A.R.; Rapoport, T.A. Protein translocation is mediated by oligomers of the SecY complex with one SecY copy forming the channel. Cell 2007, 129, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Spann, D.; Pross, E.; Chen, Y.; Dalbey, R.E.; Kuhn, A. Each protomer of a dimeric YidC functions as a single membrane insertase. Sci. Rep. 2018, 8, 589. [Google Scholar] [CrossRef] [Green Version]
- Yi, L.; Jiang, F.; Chen, M.; Cain, B.; Bolhuis, A.; Dalbey, R.E. YidC is strictly required for membrane insertion of subunits a and c of the F(1)F(0)ATP synthase and SecE of the SecYEG translocase. Biochemistry 2003, 42, 10537–10544. [Google Scholar] [CrossRef]
- Neugebauer, S.A.; Baulig, A.; Kuhn, A.; Facey, S.J. Membrane protein insertion of variant MscL proteins occurs at YidC and SecYEG of Escherichia coli. J. Mol. Biol. 2012, 417, 375–386. [Google Scholar] [CrossRef]
- Pross, E.; Soussoula, L.; Seitl, I.; Lupo, D.; Kuhn, A. Membrane Targeting and Insertion of the C-Tail Protein SciP. J. Mol. Biol. 2016, 428, 4218–4227. [Google Scholar] [CrossRef]
- Houbiers, M.; Spruijt, R.B.; Demel, R.A.; Hemminga, M.A.; Wolfs, C.J. Spontaneous insertion of gene 9 minor coat protein of bacteriophage M13 in model membranes. Biochim. Biophys. Acta Biomembr. 2001, 1511, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Jespers, L.S.; Messens, J.H.; de Keyser, A.; Eeckhout, D.; van den Brande, I.; Gansemans, Y.G.; Lauwereys, M.J.; Vlasuk, G.P.; Stanssens, P.E. Surface expression and ligand-based selection of cDNAs fused to filamentous phage gene VI. Biotechnology 1995, 13, 378–382. [Google Scholar] [CrossRef]
- Gailus, V.; Rasched, I. The adsorption protein of bacteriophage fd and its neighbour minor coat protein build a structural entity. Eur. J. Biochem. 1994, 222, 927–931. [Google Scholar] [CrossRef]
- Ploss, M.; Kuhn, A. Kinetics of filamentous phage assembly. Phys. Biol. 2010, 7, 45002. [Google Scholar] [CrossRef]
- Winterfeld, S.; Ernst, S.; Börsch, M.; Gerken, U.; Kuhn, A. Real time observation of single membrane protein insertion events by the Escherichia coli insertase YidC. PLoS ONE 2013, 8, e59023. [Google Scholar] [CrossRef] [Green Version]
- Kudva, R.; Denks, K.; Kuhn, P.; Vogt, A.; Müller, M.; Koch, H.-G. Protein translocation across the inner membrane of Gram-negative bacteria: The Sec and Tat dependent protein transport pathways. Res. Microbiol. 2013, 164, 505–534. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kleinbeck, F.; Kuhn, A. Membrane Insertion of the M13 Minor Coat Protein G3p Is Dependent on YidC and the SecAYEG Translocase. Viruses 2021, 13, 1414. https://doi.org/10.3390/v13071414
Kleinbeck F, Kuhn A. Membrane Insertion of the M13 Minor Coat Protein G3p Is Dependent on YidC and the SecAYEG Translocase. Viruses. 2021; 13(7):1414. https://doi.org/10.3390/v13071414
Chicago/Turabian StyleKleinbeck, Farina, and Andreas Kuhn. 2021. "Membrane Insertion of the M13 Minor Coat Protein G3p Is Dependent on YidC and the SecAYEG Translocase" Viruses 13, no. 7: 1414. https://doi.org/10.3390/v13071414
APA StyleKleinbeck, F., & Kuhn, A. (2021). Membrane Insertion of the M13 Minor Coat Protein G3p Is Dependent on YidC and the SecAYEG Translocase. Viruses, 13(7), 1414. https://doi.org/10.3390/v13071414