
Intravirion DNA Can Access the Space Occupied by the Bacteriophage P22 Ejection Proteins

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phage and Bacteria Strains
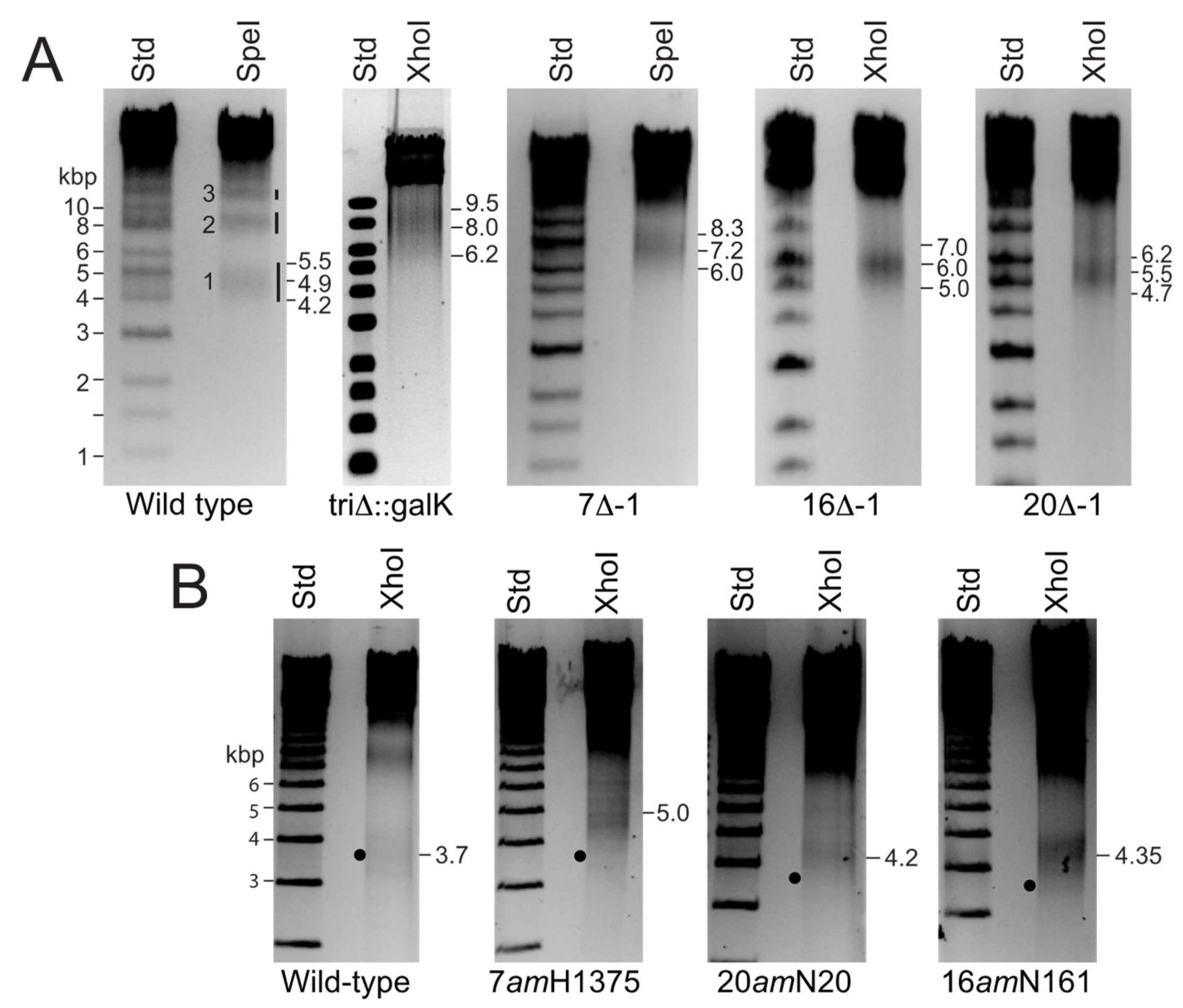
2.2. P22 Virion-Like Particle Purification, DNA Preparation and DNA Electrophoresis
3. Results
3.1. Some P22 E-Protein N-Terminal Fragments Are Assembly Proficient
3.2. P22 E-Proteins Compete with DNA for Internal Virion Space
3.3. Does Missing Protein Volume Correlate with Extra DNA Volume?
4. Discussion
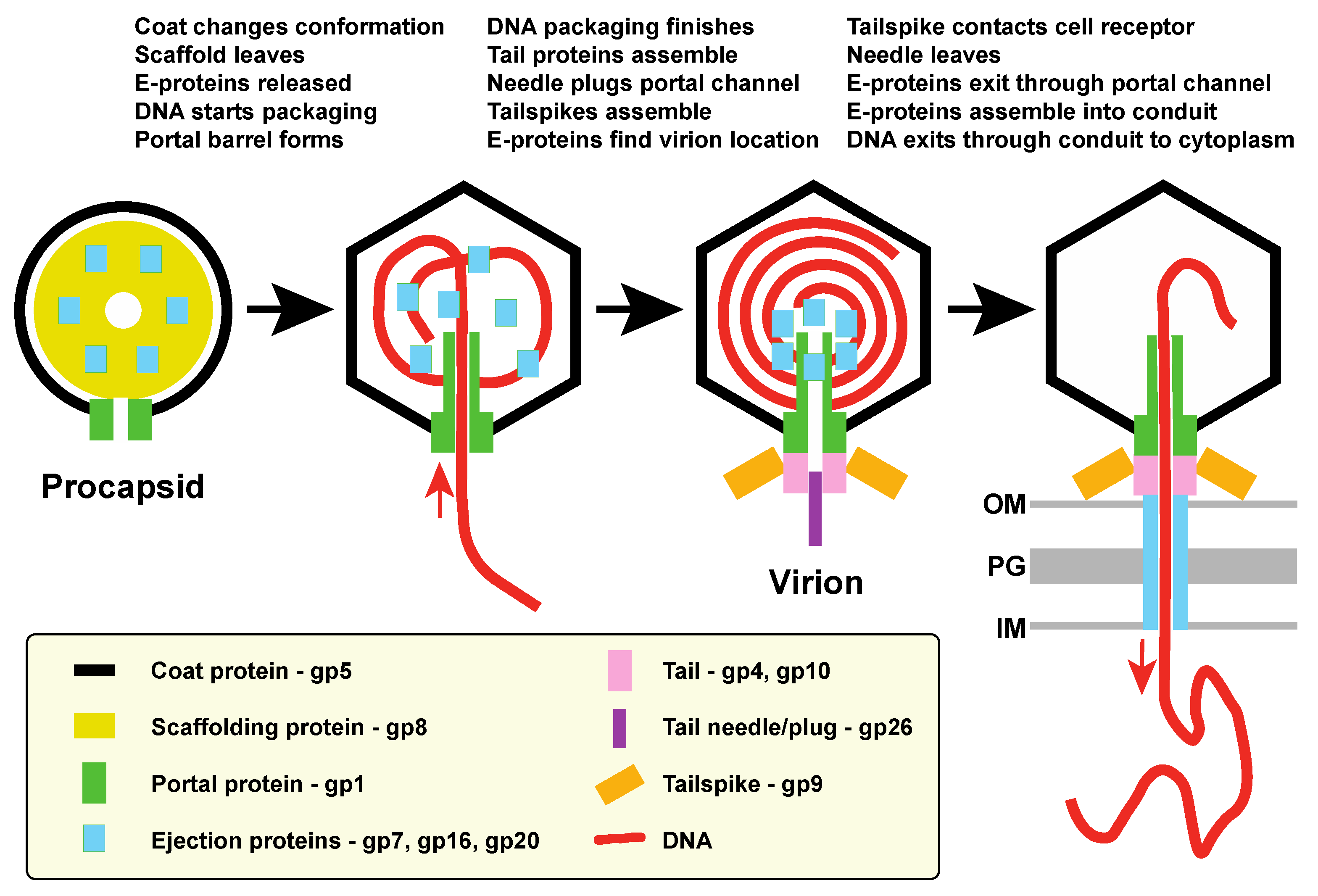
4.1. E-Protein Assembly and Intravirion Location
4.2. A Model for E-Protein Actions
- E-proteins are incorporated into procapsids by interactions with assembling scaffolding protein (and possibly coat protein?), and at least in the cases of gp7 and gp20 it is the N-terminal portion that is responsible for such interactions. Portal protein is not required for E-protein incorporation into virions.
- During DNA packaging and procapsid maturation, scaffolding protein is released from the particles, coat protein undergoes a major conformational change as the shell expands, and the C-terminal portal barrel appears to form [16,25]. These changes likely release the E-proteins from their initial binding sites and may form new ones.
- The gp26 trimer needle binds to plug the portal channel and complete the formation of the infectious virion.
- Upon their release from scaffolding protein, E-proteins move to the near central position in the virion where the bubblegram analysis of virions places them and at least parts of them are in contact with DNA. They could either bind to the newly formed portal barrel or accumulate there because of possibly lower DNA density at the center of the particle. In DNA-filled particles that lack gp26 the E-proteins seem to not yet have reached the final poised-for-ejection location, since DNA can exit but the E-proteins remain associated with the particles. Some consequence of gp26 binding may allow the E-proteins to reach the positions from which they can exit before the DNA during injection. We suggest that this movement may place at least some portion of the E-proteins inside the portal channel. The order of these latter events is unclear, and cryoEM reconstruction of 26 minus particles suggests that the ejection proteins may begin to build the virion proximal portion of the conduit when DNA is spontaneously released under these conditions [69].The interior volume of the P22 virion portal vertex channel calculated from the virion structure of Tang et al. [16] (including the inside channel of the portal barrel and the ~120 Å long portion that extends beyond the coat protein shell (see [64])) is only about 0.3 × 106 Å3. Thus, at most only about 10% of the total E-protein mass could fit into the channel (making it very unlikely that any E-protein parts actually occupying the channel of the virion would significantly affect the above volume calculations). Nonetheless, it is interesting to note that the 12 gp7 molecules, whose volume is calculated to also be 0.3 × 106 Å3, would rather neatly fill the portal vertex channel. It has been suggested that gp7 builds the virion proximal portion of the periplasmic conduit [12], and if so, it seems sensible that it would leave first so as to build the conduit from the virion outward.
- Upon successful attachment of the virion’s tail to a target cell, the gp26 plug is removed, and E-proteins leave the virion via the portal channel before the DNA is injected and assemble into the trans-periplasm conduit. It is not known if every E-protein molecule in the virion participates in the assembly of the conduit; if E-proteins are present in excess in the virion, it may not be completely debilitating if fewer than the full complement of molecules are present in the virion.
- DNA passes out of the virion through the cell membranes, peptidoglycan and periplasm into the cytoplasm via this conduit. Curiously, the P22 E-protein conduit appears to be weakly attached to the virion, since it was not present when spent infecting virions were isolated after having released their DNA into cells [6].
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Botstein, D.; Waddell, C.H.; King, J. Mechanism of head assembly and DNA encapsulation in Salmonella phage P22: I. Genes, proteins, structures and DNA maturation. J. Mol. Biol. 1973, 80, 669–695. [Google Scholar] [CrossRef]
- Hoffman, B.; Levine, M. Bacteriophage P22 virion protein which performs an essential early function. II. Characterization of the gene 16 function. J. Virol. 1975, 16, 1547–1559. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, B.; Levine, M. Bacteriophage P22 virion protein which performs an essential early function. I. Analysis of 16-ts mutants. J. Virol. 1975, 16, 1536–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, J.; Lenk, E.V.; Botstein, D. Mechanism of head assembly and DNA encapsulation in Salmonella phage P22: II. Morphogenetic pathway. J. Mol. Biol. 1973, 80, 697–731. [Google Scholar] [CrossRef]
- Poteete, A.R.; King, J. Functions of two new genes in Salmonella phage P22 assembly. Virology 1977, 76, 725–739. [Google Scholar] [CrossRef]
- Israel, V. E proteins of bacteriophage P22. I. Identification and ejection from wild-type and defective particles. J. Virol. 1977, 23, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Casjens, S.R.; Molineux, I.J. Short Noncontractile Tail Machines: Adsorption and DNA Delivery by Podoviruses. Adv. Exp. Med. Biol. 2012, 726, 143–179. [Google Scholar] [CrossRef]
- Molineux, I.J. No syringes please, ejection of phage T7 DNA from the virion is enzyme driven. Mol. Microbiol. 2001, 40, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Molineux, I.J.; Panja, D. Popping the cork: Mechanisms of phage genome ejection. Nat. Rev. Genet. 2013, 11, 194–204. [Google Scholar] [CrossRef]
- Perez, G.L.; Huynh, B.; Slater, M.; Maloy, S. Transport of Phage P22 DNA across the Cytoplasmic Membrane. J. Bacteriol. 2009, 191, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Speir, J.A.; Matsui, T.; Lin, Z.; Liang, L.; Lynn, A.; Varnado, B.; Weiss, T.M.; Tang, L. Structure of a Bacterial Virus DNA-Injection Protein Complex Reveals a Decameric Assembly with a Constricted Molecular Channel. PLoS ONE 2016, 11, e0149337. [Google Scholar] [CrossRef]
- Wang, C.; Tu, J.; Liu, J.; Molineux, I.J. Structural dynamics of bacteriophage P22 infection initiation revealed by cryo-electron tomography. Nat. Microbiol. 2019, 4, 1049–1056. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Margolin, W.; Molineux, I.J.; Liu, J. The Bacteriophage T7 Virion Undergoes Extensive Structural Remodeling During Infection. Science 2013, 339, 576–579. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-Y.; Kemp, P.; Molineux, I.J. Gp15 and gp16 cooperate in translocating bacteriophage T7 DNA into the infected cell. Virology 2010, 398, 176–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp, P.; Garcia, L.R.; Molineux, I.J. Changes in bacteriophage T7 virion structure at the initiation of infection. Virology 2005, 340, 307–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Lander, G.C.; Olia, A.S.; Li, R.; Casjens, S.; Prevelige, P.; Cingolani, G.; Baker, T.S.; Johnson, J.E. Peering Down the Barrel of a Bacteriophage Portal: The Genome Packaging and Release Valve in P22. Structure 2011, 19, 496–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olia, A.S.; Casjens, S.; Cingolani, G. Structure of phage P22 cell envelope–penetrating needle. Nat. Struct. Mol. Biol. 2007, 14, 1221–1226. [Google Scholar] [CrossRef] [PubMed]
- Olia, A.S.; Prevelige, P.E.; Johnson, J.E.; Cingolani, G. Three-dimensional structure of a viral genome-delivery portal vertex. Nat. Struct. Mol. Biol. 2011, 18, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Steinbacher, S.; Seckler, R.; Miller, S.; Steipe, B.; Huber, R.; Reinemer, P. Crystal structure of P22 tailspike protein: Interdigi-tated subunits in a thermostable trimer. Science 1994, 265, 383–386. [Google Scholar] [CrossRef]
- Steinbacher, S.; Miller, S.; Baxa, U.; Budisa, N.; Weintraub, A.; Seckler, R.; Huber, R. Phage P22 tailspike protein: Crystal structure of the head-binding domain at 2.3 Å, fully refined structure of the endorhamnosidase at 1.56 Å resolution, and the molecular basis of O-antigen recognition and cleavage. J. Mol. Biol. 1997, 267, 865–880. [Google Scholar] [CrossRef]
- Teschke, C.M.; Parent, K.N. ‘Let the phage do the work’: Using the phage P22 coat protein structures as a framework to understand its folding and assembly mutants. Virology 2010, 401, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Hryc, C.F.; Chen, D.-H.; Afonine, P.V.; Jakana, J.; Wang, Z.; Haase-Pettingell, C.; Jiang, W.; Adams, P.; King, J.A.; Schmid, M.F.; et al. Accurate model annotation of a near-atomic resolution cryo-EM map. Proc. Natl. Acad. Sci. USA 2017, 114, 3103–3108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olia, A.S.; Bhardwaj, A.; Joss, L.; Casjens, S.; Cingolani, G. Role of Gene 10 Protein in the Hierarchical Assembly of the Bacteriophage P22 Portal Vertex Structure. Biochemistry 2007, 46, 8776–8784. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Weigele, P.; King, J.; Chiu, W.; Jiang, W. Cryo-EM asymmetric reconstruction of bacteriophage P22 reveals organi-zation of its DNA packaging and infecting machinery. Structure 2006, 14, 1073–1082. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.-H.; Baker, M.L.; Hryc, C.; DiMaio, F.; Jakana, J.; Wu, W.; Dougherty, M.; Haase-Pettingell, C.; Schmid, M.F.; Jiang, W.; et al. Structural basis for scaffolding-mediated assembly and maturation of a dsDNA virus. Proc. Natl. Acad. Sci. USA 2011, 108, 1355–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Leavitt, J.C.; Cheng, N.; Gilcrease, E.B.; Motwani, T.; Teschke, C.M.; Casjens, S.R.; Steven, A.C. Localization of the Houdinisome (Ejection Proteins) inside the Bacteriophage P22 Virion by Bubblegram Imaging. mBio 2016, 7, e01152-16. [Google Scholar] [CrossRef] [Green Version]
- Bryant, J.L.; King, J. DNA injection proteins are targets of acridine-sensitized photoinactivation of bacteriophage P22. J. Mol. Biol. 1984, 180, 837–863. [Google Scholar] [CrossRef]
- Mullaney, J.; Black, L.W. Activity of foreign proteins targeted within the bacteriophage T4 head and prohead: Implications for packaged DNA structure. J. Mol. Biol. 1998, 283, 913–929. [Google Scholar] [CrossRef]
- Hong, Y.R.; Black, L.W. An expression-packaging-processing vector which selects and maintains 7-kb DNA inserts in the blue T4 phage genome. Gene 1993, 136, 193–198. [Google Scholar]
- Abremski, K.; Black, L.W. The function of bacteriophage T4 internal protein I in a restrictive strain of Escherichia coli. Virology 1979, 97, 439–447. [Google Scholar] [CrossRef]
- Black, L.W.; Abremski, K. Restriction of phage T4 internal protein I mutants by a strain of Escherichia coli. Virology 1974, 60, 180–191. [Google Scholar] [CrossRef]
- Cortines, J.; Weigele, P.R.; Gilcrease, E.B.; Casjens, S.R.; Teschke, C.M. Decoding bacteriophage P22 assembly: Identification of two charged residues in scaffolding protein responsible for coat protein interaction. Virology 2011, 421, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leavitt, J.C.; Gilcrease, E.B.; Wilson, K.; Casjens, S.R. Function and horizontal transfer of the small terminase subunit of the tailed bacteriophage Sf6 DNA packaging nanomotor. Virology 2013, 440, 117–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla-Meier, G.P.; Gilcrease, E.B.; Weigele, P.R.; Cortines, J.; Siegel, M.; Leavitt, J.C.; Teschke, C.M.; Casjens, S.R. Unraveling the Role of the C-terminal Helix Turn Helix of the Coat-binding Domain of Bacteriophage P22 Scaffolding Protein. J. Biol. Chem. 2012, 287, 33766–33780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [Green Version]
- Earnshaw, W.; Casjens, S.; Harrison, S.C. Assembly of the head of bacteriophage P22: X-ray diffraction from heads, proheads and related structures. J. Mol. Biol. 1976, 104, 387–410. [Google Scholar] [CrossRef]
- Casjens, S.; Hayden, M. Analysis in vivo of the bacteriophage P22 headful nuclease. J. Mol. Biol. 1988, 199, 467–474. [Google Scholar] [CrossRef]
- Adhikari, P.; Berget, P.B. Sequence of a DNA injection gene from Salmonella typhimurium phage P22. Nucleic Acids Res. 1993, 21, 1499. [Google Scholar] [CrossRef] [Green Version]
- Conlin, C.A.; Vimr, E.R.; Miller, C.G. Oligopeptidase A is required for normal phage P22 development. J. Bacteriol. 1992, 174, 5869–5880. [Google Scholar] [CrossRef] [Green Version]
- Eppler, K.; Wyckoff, E.; Goates, J.; Parr, R.; Casjens, S. Nucleotide sequence of the bacteriophage P22 genes required for DNA packaging. Virology 1991, 183, 519–538. [Google Scholar] [CrossRef]
- Jackson, E.N.; Jackson, D.A.; Deans, R.J. EcoRI analysis of bacteriophage P22 DNA packaging. J. Mol. Biol. 1978, 118, 365–388. [Google Scholar] [CrossRef] [Green Version]
- Tye, B.-K.; Huberman, J.A.; Botstein, D. Non-random circular permutation of phage P22 DNA. J. Mol. Biol. 1974, 85, 501–527. [Google Scholar] [CrossRef]
- Tye, B.-K.; Chan, R.K.; Botstein, D. Packaging of an oversize transducing genome by Salmonella phage P22. J. Mol. Biol. 1974, 85, 485–500. [Google Scholar] [CrossRef]
- Moore, S.D.; Prevelige, P.E., Jr. Bacteriophage P22 portal vertex formation in vivo. J. Mol. Biol. 2002, 315, 975–994. [Google Scholar] [CrossRef] [Green Version]
- Weigele, P.R.; Sampson, L.; Winn-Stapley, D.; Casjens, S.R. Molecular Genetics of Bacteriophage P22 Scaffolding Protein’s Functional Domains. J. Mol. Biol. 2005, 348, 831–844. [Google Scholar] [CrossRef]
- Berndsen, Z.T.; Keller, N.; Grimes, S.; Jardine, P.; Smith, D.E. Nonequilibrium dynamics and ultraslow relaxation of confined DNA during viral packaging. Proc. Natl. Acad. Sci. USA 2014, 111, 8345–8350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earnshaw, W.; Hendrix, R.W.; King, J. Structural studies of bacteriophage lambda heads and proheads by small angle X-ray diffraction. J. Mol. Biol. 1979, 134, 575–594. [Google Scholar] [CrossRef]
- Evilevitch, A. The mobility of packaged phage genome controls ejection dynamics. eLife 2018, 7, e37345. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.; Wyckoff, E.; Hayden, M.; Sampson, L.; Eppler, K.; Randall, S.; Moreno, E.T.; Serwer, P. Bacteriophage P22 portal protein is part of the gauge that regulates packing density of intravirion DNA. J. Mol. Biol. 1992, 224, 1055–1074. [Google Scholar] [CrossRef]
- Harpaz, Y.; Gerstein, M.; Chothia, C. Volume changes on protein folding. Structure 1994, 2, 641–649. [Google Scholar] [CrossRef] [Green Version]
- Erickson, H.P. Size and Shape of Protein Molecules at the Nanometer Level Determined by Sedimentation, Gel Filtration, and Electron Microscopy. Biol. Proced. Online 2009, 11, 32–51. [Google Scholar] [CrossRef] [Green Version]
- Keifer, D.Z.; Motwani, T.; Teschke, C.M.; Jarrold, M.F. Measurement of the accurate mass of a 50 MDa infectious virus. Rapid Commun. Mass Spectrom. 2016, 30, 1957–1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earnshaw, W.; Casjens, S.R. DNA packaging by the double-stranded DNA bacteriophages. Cell 1980, 21, 319–331. [Google Scholar] [CrossRef]
- Serwer, P. Buoyant density sedimentation of macromolecules in sodium iothalamate density gradients. J. Mol. Biol. 1975, 92, 433–448. [Google Scholar] [CrossRef]
- Casjens, S.; King, J. P22 morphogenesis I: Catalytic scaffolding protein in capsid assembly. J. Supramol. Struct. 1974, 2, 202–224. [Google Scholar] [CrossRef] [PubMed]
- Mullaney, J.M.; Black, L.W. Capsid Targeting Sequence Targets Foreign Proteins into Bacteriophage T4 and Permits Proteolytic Processing. J. Mol. Biol. 1996, 261, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Black, L.W. Bacteriophage T4 internal protein mutants: Isolation and properties. Virology 1974, 60, 166–179. [Google Scholar] [CrossRef]
- Earnshaw, W.; King, J. Structure of phage P22 coat protein aggregates formed in the absence of the scaffolding protein. J. Mol. Biol. 1978, 126, 721–747. [Google Scholar] [CrossRef]
- Greene, B.; King, J. Scaffolding Mutants Identifying Domains Required for P22 Procapsid Assembly and Maturation. Virology 1996, 225, 82–96. [Google Scholar] [CrossRef] [Green Version]
- King, J.; Casjens, S. Catalytic head assembling protein in virus morphogenesis. Nature 1974, 251, 112–119. [Google Scholar] [CrossRef]
- Lander, G.C.; Tang, L.; Casjens, S.R.; Gilcrease, E.B.; Prevelige, P.; Poliakov, A.; Potter, C.S.; Carragher, B.; Johnson, J.E. The Structure of an Infectious P22 Virion Shows the Signal for Headful DNA Packaging. Science 2006, 312, 1791–1795. [Google Scholar] [CrossRef] [Green Version]
- Leavitt, J.C.; Gogokhia, L.; Gilcrease, E.B.; Bhardwaj, A.; Cingolani, G.; Casjens, S.R. The Tip of the Tail Needle Affects the Rate of DNA Delivery by Bacteriophage P22. PLoS ONE 2013, 8, e70936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Sdao, S.M.; Dover, J.A.; Porcek, N.B.; Knobler, C.M.; Gelbart, W.M.; Parent, K.N. Bacteriophage P22 ejects all of its internal proteins before its genome. Virology 2015, 485, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhardwaj, A.; Sankhala, R.S.; Olia, A.S.; Brooke, D.; Casjens, S.R.; Taylor, D.J.; Cingolani, G. Structural Plasticity of the Protein Plug That Traps Newly Packaged Genomes in Podoviridae Virions. J. Biol. Chem. 2016, 291, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Bauer, D.W.; Li, D.; Huffman, J.B.; Homa, F.L.; Wilson, K.T.; Leavitt, J.C.; Casjens, S.; Baines, J.D.; Evilevitch, A. Exploring the Balance between DNA Pressure and Capsid Stability in Herpesviruses and Phages. J. Virol. 2015, 89, 9288–9298. [Google Scholar] [CrossRef] [Green Version]
- Berget, P.; Poteete, A.R. Structure and Functions of the Bacteriophage P22 Tail Protein. J. Virol. 1980, 34, 234. [Google Scholar] [CrossRef] [Green Version]
- Lenk, E.; Casjens, S.; Weeks, J.; King, J. Intracellular visualization of precursor capsids in phage P22 mutant infected cells. Virology 1975, 68, 182–199. [Google Scholar] [CrossRef]
- Strauss, H.; King, J. Steps in the stabilization of newly packaged DNA during phage P22 Morphogenesis. J. Mol. Biol. 1984, 172, 523–543. [Google Scholar] [CrossRef]
- McNulty, R.; Cardone, G.; Gilcrease, E.; Baker, T.; Casjens, S.; Johnson, J. Cryo-EM elucidation of the structure of bacterio-phage P22 virions after genome release. Biophys. J. 2018, 114, 1295–1301. [Google Scholar] [CrossRef] [Green Version]
- Kazmierczak, K.; Davydova, E.; Mustaev, A.; Rothman-Denes, L. The phage N4 virion RNA polymerase catalytic domain is related to single-subunit RNA polymerases. EMBO J. 2002, 21, 5815–5823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceyssens, P.-J.; Minakhin, L.; Bossche, A.V.D.; Yakunina, M.; Klimuk, E.; Blasdel, B.; De Smet, J.; Noben, J.-P.; Bläsi, U.; Severinov, K.; et al. Development of Giant Bacteriophage KZ Is Independent of the Host Transcription Apparatus. J. Virol. 2014, 88, 10501–10510. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.S.; Kutter, E.; Mosig, G.; Arisaka, F.; Kunisawa, T.; Ruger, W. Bacteriophage T4 genome. Microbiol. Mol. Biol. Rev. 2003, 67, 86–156. [Google Scholar] [CrossRef] [Green Version]
- Rifat, D.; Wright, N.T.; Varney, K.M.; Weber, D.J.; Black, L.W. Restriction Endonuclease Inhibitor IPI* of Bacteriophage T4: A Novel Structure for a Dedicated Target. J. Mol. Biol. 2008, 375, 720–734. [Google Scholar] [CrossRef] [Green Version]
- Piya, D.; Vara, L.; Russell, W.K.; Young, R.; Gill, J.J. The multicomponent antirestriction system of phage P1 is linked to capsid morphogenesis. Mol. Microbiol. 2017, 105, 399–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, L.W.; Thomas, J.A. Condensed Genome Structure. Adv. Exp. Med. Biol. 2011, 726, 469–487. [Google Scholar] [CrossRef] [Green Version]
- Ali, B.; Desmond, M.I.; Mallory, S.A.; Benítez, A.D.; Buckley, L.J.; Weintraub, S.T.; Osier, M.V.; Black, L.W.; Thomas, J.A. To Be or Not To Be T4: Evidence of a Complex Evolutionary Pathway of Head Structure and Assembly in Giant Salmonella Virus SPN3US. Front. Microbiol. 2017, 8, 2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, J.; Summer, E.J.; Russell, W.K.; Cologna, S.M.; Carlile, T.M.; Fuller, A.C.; Kitsopoulos, K.; Mebane, L.M.; Parkinson, B.N.; Sullivan, D.; et al. Genomes and Characterization of Phages Bcep22 and BcepIL02, Founders of a Novel Phage Type in Burkholderia cenocepacia. J. Bacteriol. 2011, 193, 5300–5313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, D.W.; Casjens, S.R.; Sharma, R.; Grose, J.H. Genomic comparison of 60 completely sequenced bacteriophages that infect Erwinia and/or Pantoea bacteria. Virology 2019, 535, 59–73. [Google Scholar] [CrossRef]
- Leptihn, S.; Gottschalk, J.; Kuhn, A. T7 ejectosome assembly: A story unfolds. Bacteriophage 2016, 6, e1128513. [Google Scholar] [CrossRef] [Green Version]
- Lupo, D.; Leptihn, S.; Nagler, G.; Haase, M.; Molineux, I.J.; Kuhn, A. The T7 ejection nanomachine components gp15–gp16 form a spiral ring complex that binds DNA and a lipid membrane. Virology 2015, 486, 263–271. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Name | Genotype a | Source |
|---|---|---|
| Salmonella enterica serovar Typhimurium LT2 | ||
| UB-2235 | galK::TetRA-1,∆Fels-1,∆Fels-2,∆Gifsy-1,∆Gifsy-2 (P22 UC-0937) | b |
| UB-2272 | UB-2235 (P22 UC-0937 with galK::7) c | [26] |
| UB-2274 | UB-2235 (P22 UC-0937 with galK::16) c | [26] |
| UB-2276 | UB-2235 (P22 UC-0937 with galK::20) c | [26] |
| UB-2278 | UB-2235 (P22 UC-0937 with tri∆::galK) | [26] |
| UB-2285 | UB-2235 (P22 UC-0937 with 7−∆-1) d | [26] |
| UB-2288 | UB-2235 (P22 UC-0937 with 16−∆-1) d | [26] |
| UB-2289 | UB-2235 (P22 UC-0937 with 20−∆-1) d | [26] |
| UB-2350 | UB-2235 (P22 UC-0937 with 7−amH1375) | this report |
| UB-2366 | UB-2235 (P22 UC-0937 with 16−amN121) | this report |
| UB-2387 | UB-2235 (P22 UC-0937 with 20−amN20) | this report |
| P22 bacteriophages | ||
| UC-0008 | P22 c1-7, 13−amH101, 2−amN20 | [1] |
| UC-0009 | P22 c1-7, 13−amH101, 16−amN121 | [1] |
| UC-0100 | P22 c1-7, 13−amH101, 7−amH1375 | [5] |
| UC-0937 | P22 c1-7, 13−amH101, orf25::CamR-EG1, sieA-∆1 | b |
| Right-End | |||||
|---|---|---|---|---|---|
| Genome | Restriction | Fragment | Packaged | DNA Length | |
| P22 Phage | Length a | Site Location b | Length c | DNA Length d | Increase e |
| WT (UC-0937) | 39,521 | –769 (SpeI) | 4650 | 43,400 | – |
| 20−∆-1 | 38,161 | +257 (XhoI) | 5500 | 43,900 | 500 |
| 20−amN20 | 39,521 | +257 (XhoI) | 4200 | 44,000 | 600 |
| 16−∆-1 | 37,751 | +257 (XhoI) | 6100 | 44,100 | 700 |
| 16−amN121 | 39,521 | +257 (XhoI) | 4400 | 44,200 | 800 |
| 7−∆-1 | 38,890 | –769(SpeI) | 7000 | 45,100 | 1700 |
| 7−amH1375 | 39,521 | +257 (XhoI) | 5000 | 44,800 | 1400 |
| tri∆::galK | 36,867 | +257 (XhoI) | 8000 | 45,100 | 1700 |
| Average | Volume b | Volume c | ||
|---|---|---|---|---|
| bp Length | of “Extra” | of “Missing” E- | ||
| P22 Phage | Increase a | DNA (Å3 × 10−6) | Proteins (Å3 × 10−6) | Volume Ratio d |
| 20−∆-1 | 500 | 1.1 | 2.1 | 0.52 |
| 20−amN20 | 600 | 1.3 | 2.4 (2.1) e | 0.54 (0.62) e |
| 16−∆-1 | 700 | 1.5 | 1.5 | 1.0 |
| 16−amN121 | 800 | 1.6 | 1.3 | 0.8 |
| 7−∆-1 | 1700 | 3.6 | 2.3 | 1.6 |
| 7−amH1375 | 1400 | 2.9 | 1.5 (1.8) e | 2.0 (1.6) e |
| tri∆::galK | 1700 | 3.6 | 3.0 | 1.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leavitt, J.C.; Gilcrease, E.B.; Woodbury, B.M.; Teschke, C.M.; Casjens, S.R. Intravirion DNA Can Access the Space Occupied by the Bacteriophage P22 Ejection Proteins. Viruses 2021, 13, 1504. https://doi.org/10.3390/v13081504
Leavitt JC, Gilcrease EB, Woodbury BM, Teschke CM, Casjens SR. Intravirion DNA Can Access the Space Occupied by the Bacteriophage P22 Ejection Proteins. Viruses. 2021; 13(8):1504. https://doi.org/10.3390/v13081504
Chicago/Turabian StyleLeavitt, Justin C., Eddie B. Gilcrease, Brianna M. Woodbury, Carolyn M. Teschke, and Sherwood R. Casjens. 2021. "Intravirion DNA Can Access the Space Occupied by the Bacteriophage P22 Ejection Proteins" Viruses 13, no. 8: 1504. https://doi.org/10.3390/v13081504
APA StyleLeavitt, J. C., Gilcrease, E. B., Woodbury, B. M., Teschke, C. M., & Casjens, S. R. (2021). Intravirion DNA Can Access the Space Occupied by the Bacteriophage P22 Ejection Proteins. Viruses, 13(8), 1504. https://doi.org/10.3390/v13081504