
A New Molecular Detection System for Canine Distemper Virus Based on a Double-Check Strategy


Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. CDV-RT-qPCR Assays
2.3. Generation of a CDV Standard
2.4. Analytical Sensitivity
2.5. Analytical Specificity
2.6. Diagnostic Sensitivity and Specificity
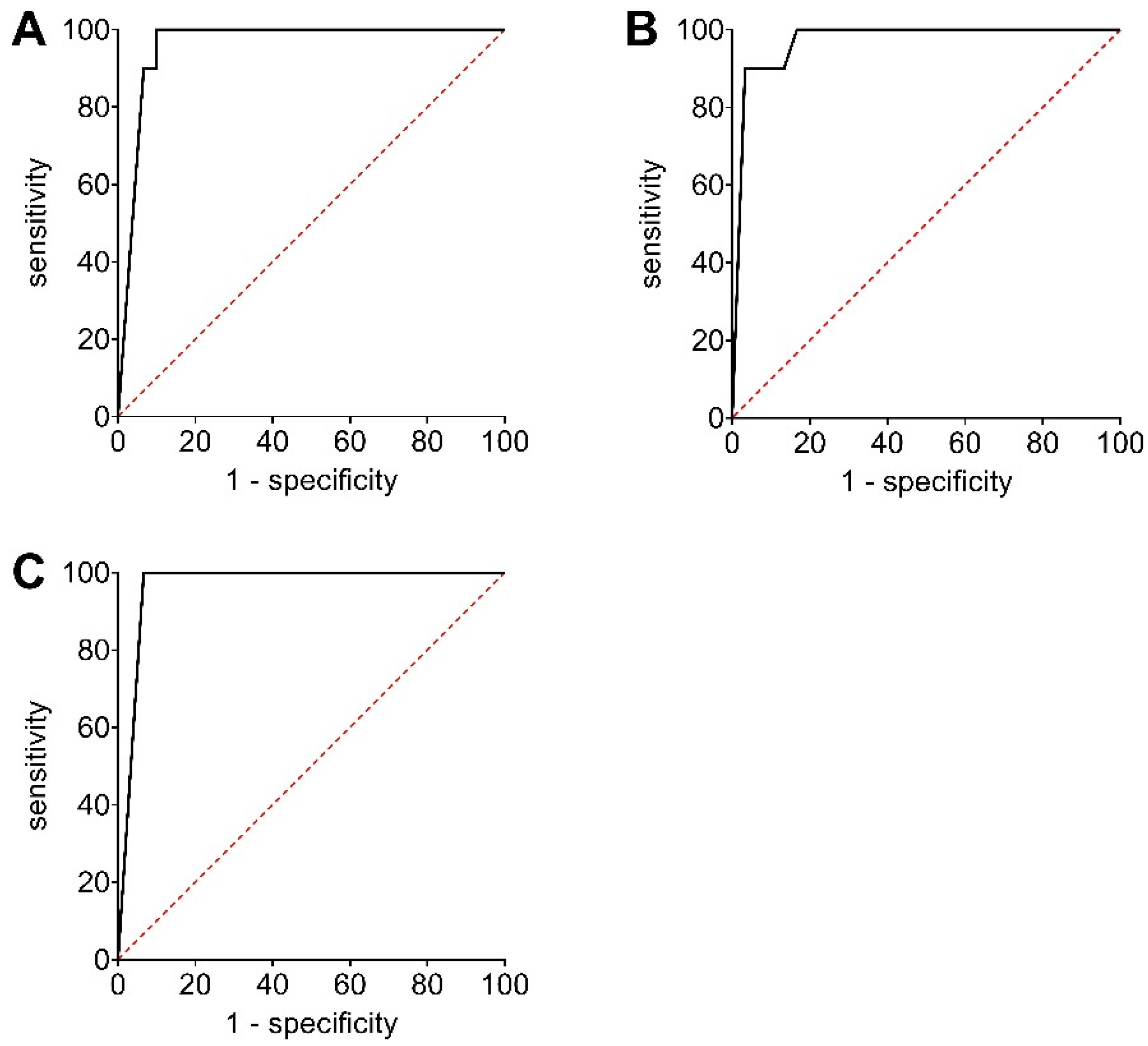
2.7. Receiver Operating Curve (ROC) Analyses
2.8. Quality Assessment and Bias Statement
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beineke, A.; Baumgartner, W.; Wohlsein, P. Cross-species transmission of canine distemper virus-an update. One Health 2015, 1, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Kapil, S.; Yeary, T.J. Canine distemper spillover in domestic dogs from urban wildlife. Vet. Clin. N. Am. Small Anim. Pract. 2011, 41, 1069–1086. [Google Scholar] [CrossRef]
- Martinez-Gutierrez, M.; Ruiz-Saenz, J. Diversity of susceptible hosts in canine distemper virus infection: A systematic review and data synthesis. BMC Vet. Res. 2016, 12, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barthold, S.W.; Bowen, R.A.; Hedrick, R.P.; Knowles, D.P.; Lairmore, M.D.; Parrish, C.R.; Saif, L.J.; Swayne, D.E. Fenner’s Veterinary Virology, 4th ed.; MacLachlan, N.J., Dubovi, E.J., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2011. [Google Scholar]
- ICTV. ICTV Taxonomy History: Canine Morbillivirus. Available online: https://talk.ictvonline.org//taxonomy/p/taxonomy-history?taxnode_id=201851613 (accessed on 3 January 2020).
- Rendon-Marin, S.; da Fontoura Budaszewski, R.; Canal, C.W.; Ruiz-Saenz, J. Tropism and molecular pathogenesis of canine distemper virus. Virol. J. 2019, 16, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duque-Valencia, J.; Sarute, N.; Olarte-Castillo, X.A.; Ruiz-Saenz, J. Evolution and Interspecies Transmission of Canine Distemper Virus-An Outlook of the Diverse Evolutionary Landscapes of a Multi-Host Virus. Viruses 2019, 11, 582. [Google Scholar] [CrossRef] [Green Version]
- Ricci, I.; Cersini, A.; Manna, G.; Marcario, G.A.; Conti, R.; Brocherel, G.; Grifoni, G.; Eleni, C.; Scicluna, M.T. A Canine Distemper Virus Retrospective Study Conducted from 2011 to 2019 in Central Italy (Latium and Tuscany Regions). Viruses 2021, 13, 272. [Google Scholar] [CrossRef] [PubMed]
- Riley, M.C.; Wilkes, R.P. Sequencing of emerging canine distemper virus strain reveals new distinct genetic lineage in the United States associated with disease in wildlife and domestic canine populations. Virol. J. 2015, 12, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anis, E.; Holford, A.L.; Galyon, G.D.; Wilkes, R.P. Antigenic analysis of genetic variants of Canine distemper virus. Vet. Microbiol. 2018, 219, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Loots, A.K.; Mitchell, E.; Dalton, D.L.; Kotze, A.; Venter, E.H. Advances in canine distemper virus pathogenesis research: A wildlife perspective. J. Gen. Virol. 2017, 98, 311–321. [Google Scholar] [CrossRef]
- Yon, L.; Duff, J.P.; Agren, E.O.; Erdelyi, K.; Ferroglio, E.; Godfroid, J.; Hars, J.; Hestvik, G.; Horton, D.; Kuiken, T.; et al. Recent Changes in Infectious Diseases in European Wildlife. J. Wildl. Dis. 2019, 55, 3–43. [Google Scholar] [CrossRef]
- Di Blasio, A.; Irico, L.; Caruso, C.; Miceli, I.; Robetto, S.; Peletto, S.; Varello, K.; Giorda, F.; Mignone, W.; Rubinetti, F.; et al. Canine Distemper Virus as an Emerging Multihost Pathogen in Wild Carnivores in Northwest Italy. J. Wildl. Dis. 2019, 55, 844–856. [Google Scholar] [CrossRef]
- Garigliany, M.; Sarlet, M.; Franssen, M.; Desmecht, D.; Volpe, R.; Lesenfants, C.; Paternostre, J.; Linden, A. Re-emergence of canine distemper in wildlife in Belgium. Vet. Rec. 2018, 182, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregers-Jensen, L.; Agger, J.F.; Hammer, A.S.; Andresen, L.; Chriel, M.; Hagberg, E.; Jensen, M.K.; Hansen, M.S.; Hjulsager, C.K.; Struve, T. Associations between biosecurity and outbreaks of canine distemper on Danish mink farms in 2012–2013. Acta Vet. Scand. 2015, 57, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molenaar, R.J.; Buter, R. Outbreaks of canine distemper in Dutch and Belgian mink farms. Vet. Q 2018, 38, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Monne, I.; Fusaro, A.; Valastro, V.; Citterio, C.; Dalla Pozza, M.; Obber, F.; Trevisiol, K.; Cova, M.; De Benedictis, P.; Bregoli, M.; et al. A distinct CDV genotype causing a major epidemic in Alpine wildlife. Vet. Microbiol. 2011, 150, 63–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renteria-Solis, Z.; Forster, C.; Aue, A.; Wittstatt, U.; Wibbelt, G.; Konig, M. Canine distemper outbreak in raccoons suggests pathogen interspecies transmission amongst alien and native carnivores in urban areas from Germany. Vet. Microbiol. 2014, 174, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Sekulin, K.; Hafner-Marx, A.; Kolodziejek, J.; Janik, D.; Schmidt, P.; Nowotny, N. Emergence of canine distemper in Bavarian wildlife associated with a specific amino acid exchange in the haemagglutinin protein. Vet. J. 2011, 187, 399–401. [Google Scholar] [CrossRef] [PubMed]
- Ek-Kommonen, C.; Sihvonen, L.; Pekkanen, K.; Rikula, U.; Nuotio, L. Outbreak off canine distemper in vaccinated dogs in Finland. Vet. Rec. 1997, 141, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Rikula, U.; Nuotio, L.; Sihvonen, L. Vaccine coverage, herd immunity and occurrence of canine distemper from 1990–1996 in Finland. Vaccine 2007, 25, 7994–7998. [Google Scholar] [CrossRef]
- Bhatt, M.; Rajak, K.K.; Chakravarti, S.; Yadav, A.K.; Kumar, A.; Gupta, V.; Chander, V.; Mathesh, K.; Chandramohan, S.; Sharma, A.K.; et al. Phylogenetic analysis of haemagglutinin gene deciphering a new genetically distinct lineage of canine distemper virus circulating among domestic dogs in India. Transbound Emerg. Dis. 2019, 66, 1252–1267. [Google Scholar] [CrossRef]
- Vergara-Wilson, V.; Hidalgo-Hermoso, E.; Sanchez, C.R.; Abarca, M.J.; Navarro, C.; Celis-Diez, S.; Soto-Guerrero, P.; Diaz-Ayala, N.; Zordan, M.; Cifuentes-Ramos, F.; et al. Canine Distemper Outbreak by Natural Infection in a Group of Vaccinated Maned Wolves in Captivity. Pathogens 2021, 10, 51. [Google Scholar] [CrossRef]
- Almuna, R.; Lopez-Perez, A.M.; Sarmiento, R.E.; Suzan, G. Drivers of canine distemper virus exposure in dogs at a wildlife interface in Janos, Mexico. Vet. Rec. Open 2021, 8, e7. [Google Scholar] [CrossRef]
- Viana, M.; Cleaveland, S.; Matthiopoulos, J.; Halliday, J.; Packer, C.; Craft, M.E.; Hampson, K.; Czupryna, A.; Dobson, A.P.; Dubovi, E.J.; et al. Dynamics of a morbillivirus at the domestic-wildlife interface: Canine distemper virus in domestic dogs and lions. Proc. Natl. Acad. Sci. USA 2015, 112, 1464–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista Linhares, M.; Whiteley, H.E.; Samuelson, J.P.; Hsiao, S.H.; Stern, A.W.; Sprandel, I.T.; Roady, P.J.; Coleman, D.A.; Rizzo, R.; Froderman, S.F.; et al. Sylvatic Canine Morbillivirus in Captive Panthera Highlights Viral Promiscuity and the Need for Better Prevention Strategies. Pathogens 2021, 10, 544. [Google Scholar] [CrossRef] [PubMed]
- Elia, G.; Decaro, N.; Martella, V.; Cirone, F.; Lucente, M.S.; Lorusso, E.; Di Trani, L.; Buonavoglia, C. Detection of canine distemper virus in dogs by real-time RT-PCR. J. Virol. Methods 2006, 136, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Scagliarini, A.; Dal Pozzo, F.; Gallina, L.; Vaccari, F.; Morganti, L. TaqMan based real time PCR for the quantification of canine distemper virus. Vet. Res. Commun. 2007, 31 (Suppl. 1), 261–263. [Google Scholar] [CrossRef] [PubMed]
- Wernike, K.; Beer, M.; Freuling, C.M.; Klupp, B.; Mettenleiter, T.C.; Muller, T.; Hoffmann, B. Molecular double-check strategy for the identification and characterization of Suid herpesvirus 1. J. Virol. Methods 2014, 209, 110–115. [Google Scholar] [CrossRef]
- Fischer, M.; Freuling, C.M.; Muller, T.; Wegelt, A.; Kooi, E.A.; Rasmussen, T.B.; Voller, K.; Marston, D.A.; Fooks, A.R.; Beer, M.; et al. Molecular double-check strategy for the identification and characterization of European Lyssaviruses. J. Virol. Methods 2014, 203, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef]
- Toussaint, J.F.; Sailleau, C.; Breard, E.; Zientara, S.; De Clercq, K. Bluetongue virus detection by two real-time RT-qPCRs targeting two different genomic segments. J. Virol. Methods 2007, 140, 115–123. [Google Scholar] [CrossRef]
- Burns, M.; Valdivia, H. Modelling the limit of detection in real-time quantitative PCR. Eur. Food Res. Technol. 2008, 226, 1513–1524. [Google Scholar] [CrossRef]
- Forootan, A.; Sjoback, R.; Bjorkman, J.; Sjogreen, B.; Linz, L.; Kubista, M. Methods to determine limit of detection and limit of quantification in quantitative real-time PCR (qPCR). Biomol. Detect. Quantif. 2017, 12, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Vandemeulebroucke, E.; De Clercq, K.; Van der Stede, Y.; Vandenbussche, F. A proposed validation method for automated nucleic acid extraction and RT-qPCR analysis: An example using Bluetongue virus. J. Virol. Methods 2010, 165, 76–82. [Google Scholar] [CrossRef] [PubMed]
- OIE. Chapter 1.1.5. Principles and methods of validation of diagnostic assays for infectious diseases. In OIE Terrestrial Manual 2013; OIE: Paris, France, 2019; Volume 2013, pp. 1–16. [Google Scholar]
- Parikh, R.; Mathai, A.; Parikh, S.; Chandra Sekhar, G.; Thomas, R. Understanding and using sensitivity, specificity and predictive values. Indian J. Ophthalmol. 2008, 56, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Akobeng, A.K. Understanding diagnostic tests 1: Sensitivity, specificity and predictive values. Acta Paediatr. 2007, 96, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Akobeng, A.K. Understanding diagnostic tests 3: Receiver operating characteristic curves. Acta Paediatr. 2007, 96, 644–647. [Google Scholar] [CrossRef]
- Whiting, P.; Rutjes, A.W.; Reitsma, J.B.; Bossuyt, P.M.; Kleijnen, J. The development of QUADAS: A tool for the quality assessment of studies of diagnostic accuracy included in systematic reviews. BMC Med. Res. Methodol. 2003, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Whiting, P.F.; Weswood, M.E.; Rutjes, A.W.; Reitsma, J.B.; Bossuyt, P.N.; Kleijnen, J. Evaluation of QUADAS, a tool for the quality assessment of diagnostic accuracy studies. BMC Med. Res. Methodol. 2006, 6, 9. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| PCR Assay | Primer/Probe | Sequence 5′–3′ | Amplicon Size (bp) | Location | Reference |
|---|---|---|---|---|---|
| ELIA-Mix | CDV-F | AGC TAG TTT CAT CTT AAC TAT CAA ATT | 87 | N gene | Elia et al., 2006 [27] |
| CDV-R | TTA ACT CTC CAG AAA ACT CAT GC | ||||
| CDV-Pb | FAM-ACC CAA GAG CCG GAT ACA TAG TTT CAA TGC-TAMRA | ||||
| CDV-Mix 3 | CDV4.1-F | CTG TCR GTA ATC GAG RAT TCG A | 116 | P gene | Scagliarini et al., 2007 |
| CDV3-R | GCC GAA AGA ATA TCC CCA GTT AG | [28], modified | |||
| CD3.1-FAMas | FAM-ATC TTC GCC AGA RTC YTC AGT GCT-BHQ1 | ||||
| CDV-Mix 7 | CDV-1808-F | AGG ARC AGG CCT AYC ATG TCA | 96 | P gene | in this study |
| CDV-1903-R | TRC TGC TGA CCT CTT GAA TCT C | ||||
| CDV-1842-FAM | FAM-ATG CCT CAA ARC CCT CAG AGA GAA TCC-BHQ1 | ||||
| EGFP-Mix 2-HEX | EGFP-1-F | GAC CAC TAC CAG CAG AAC AC | 177 | 637–794 | Hoffmann et al., 2006 |
| EGFP-10-R | CTT GTA CAG CTC GTC CAT GC | [31] | |||
| EGFP-HEX | HEX-AGC ACC CAG TCC GCC CTG AGC A-BHQ1 | ||||
| β-Actin-Mix 2-HEX | ACT-1005-F | CAG CAC AAT GAA GAT CAA GAT CAT C | 130 | 1005–1114 | Toussaint et al., 2007 |
| ACT-1135-R | CGG ACT CAT CGT ACT CCT GCT T | [32] | |||
| ACT-1081-HEX | HEX-TCG CTG TCC ACC TTC CAG CAG ATG T-BHQ1 |
| Concentration of the Template (in GE/μL) | CDV-Mix 3 | CDV-Mix 7 | ||||||
|---|---|---|---|---|---|---|---|---|
| No of Positive Detectable Replicates | Mean Cq | SD | CV% | No of Positive Detectable Replicates | Mean Cq | SD | CV% | |
| 104 | 36 | 24.2 | 0.29 | 1.18 | 36 | 23.6 | 0.17 | 0.70 |
| 103 | 36 | 27.2 | 0.14 | 0.52 | 36 | 26.7 | 0.23 | 0.87 |
| 102 | 36 | 30.2 | 0.16 | 0.54 | 36 | 29.9 | 0.22 | 0.74 |
| 101 | 36 | 33.4 | 0.31 | 0.93 | 36 | 33.5 | 0.43 | 1.29 |
| 100 | 31 | 37.0 | 1.11 | 2.99 | 13 | 38.1 | 1.15 | 3.02 |
| 10−1 | 5 | 38.7 | 0.48 | 1.23 | 1 | 38.0 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halecker, S.; Bock, S.; Beer, M.; Hoffmann, B. A New Molecular Detection System for Canine Distemper Virus Based on a Double-Check Strategy. Viruses 2021, 13, 1632. https://doi.org/10.3390/v13081632
Halecker S, Bock S, Beer M, Hoffmann B. A New Molecular Detection System for Canine Distemper Virus Based on a Double-Check Strategy. Viruses. 2021; 13(8):1632. https://doi.org/10.3390/v13081632
Chicago/Turabian StyleHalecker, Sabrina, Sabine Bock, Martin Beer, and Bernd Hoffmann. 2021. "A New Molecular Detection System for Canine Distemper Virus Based on a Double-Check Strategy" Viruses 13, no. 8: 1632. https://doi.org/10.3390/v13081632
APA StyleHalecker, S., Bock, S., Beer, M., & Hoffmann, B. (2021). A New Molecular Detection System for Canine Distemper Virus Based on a Double-Check Strategy. Viruses, 13(8), 1632. https://doi.org/10.3390/v13081632