
The Effect of Fluctuating Incubation Temperatures on West Nile Virus Infection in Culex Mosquitoes



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Insects
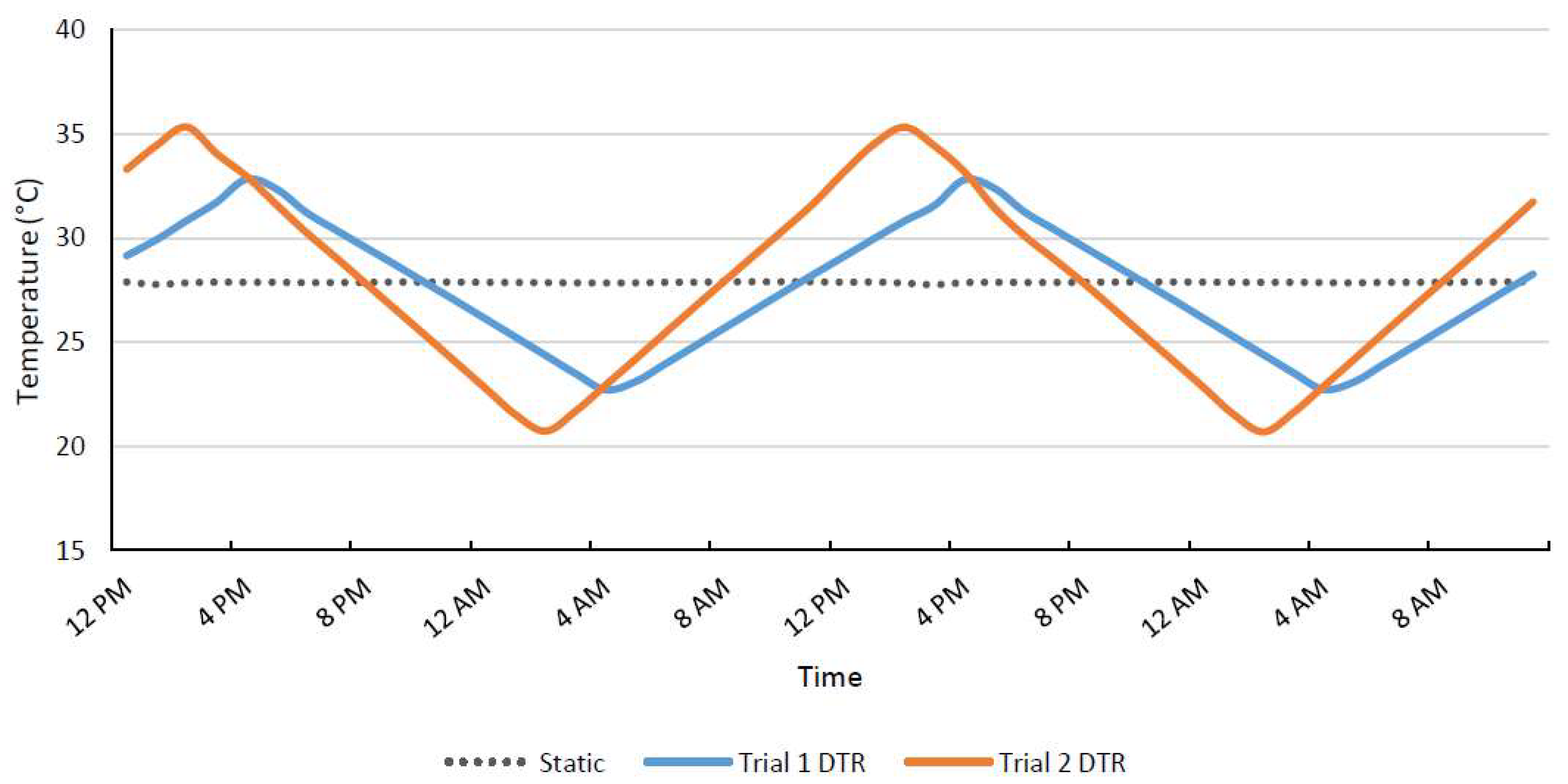
2.2. Fluctuating DTR Conditions
2.3. Infection Trials
2.4. Sample Testing
2.5. Statistical Analyses
3. Results
3.1. Trial 1: 11 °C DTR
3.2. Trial 2: 15 °C DTR
3.3. Trial Comparison—Culex quinquefasciatus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mordecai, E.A.; Caldwell, J.M.; Grossman, M.K.; Lippi, C.A.; Johnson, L.R.; Neira, M.; Rohr, J.R.; Ryan, S.J.; Savage, V.; Shocket, M.S.; et al. Thermal biology of mosquito-borne disease. Ecol. Lett. 2019, 22, 1690–1708. [Google Scholar] [CrossRef] [Green Version]
- Kramer, L.D.; Ciota, A.T. Dissecting vectorial capacity for mosquito-borne viruses. Current. Opin. Virol. 2015, 15, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, T.; Greischar, M.A.; Wadhawan, K.; Gilbert, B.; Paaijmans, K.; Mideo, N. Temperature-dependent variation in the extrinsic incubation period elevates the risk of vector-borne disease emergence. Epidemics 2019, 30, 100382. [Google Scholar] [CrossRef]
- Shelton, R.M. The effect of temperatures on development of eight mosquito species. Mosq. News 1973, 33, 1–12. [Google Scholar]
- Rueda, L.M.; Patel, K.J.; Axtell, R.C.; Stinner, R.E. Temperature-dependent development and survival rates of Culex quinquefasciatus and Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 1990, 27, 892–898. [Google Scholar] [CrossRef]
- Mohammed, A.; Chadee, D.D. Effects of different temperature regimens on the development of Aedes aegypti (L.) (Diptera: Culicidae) mosquitoes. Acta Tropica 2011, 119, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Reiskind, M.H.; Zarrabi, A.A. Is bigger really bigger? Differential responses to temperature in measures of body size of the mosquito, Aedes albopictus. J. Insect Physiol. 2012, 58, 911–917. [Google Scholar] [CrossRef]
- Ciota, A.T.; Matacchiero, A.C.; Kilpatrick, A.M.; Kramer, L.D. The effect of temperature on life history traits of Culex mosquitoes. J. Med. Entomol. 2014, 51, 55–62. [Google Scholar] [CrossRef]
- Rund, S.S.; Hou, T.Y.; Ward, S.M.; Collins, F.H.; Duffield, G.E. Genome-wide profiling of diel and circadian gene expression in the malaria vector Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2011, 108, E421–E430. [Google Scholar] [CrossRef] [Green Version]
- Rivas, G.B.S.; Teles-de-Freitas, R.; Pavan, M.G.; Lima, J.B.P.; Peixoto, A.A.; Bruno, R.V. Effects of Light and Temperature on Daily Activity and Clock Gene Expression in Two Mosquito Disease Vectors. J. Biol. Rhythm. 2018, 33, 272–288. [Google Scholar] [CrossRef]
- Reeves, W.C.; Hardy, J.L.; Reisen, W.K.; Milby, M.M. Potential effect of global warming on mosquito-borne arboviruses. J. Med. Entomol. 1994, 31, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Sternberg, E.D.; Thomas, M.B. Local adaptation to temperature and the implications for vector-borne diseases. Trends Parasitol. 2014, 30, 115–122. [Google Scholar] [CrossRef]
- Elbers, A.R.W.; Koenraadt, C.J.M.; Meiswinkel, R. Mosquitoes and Culicoides biting midges: Vector range and the influence of climate change. Rev. Sci. Tech. Off. Int. Epiz. 2015, 34, 123–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, S.P.; Keeling, M.J. The impact of temperature changes on vector-borne disease transmission: Culicoides midges and bluetongue virus. J. R. Soc. Interface 2017, 14, 20160481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turell, M.J.; O’Guinn, M.L.; Dohm, D.J.; Jones, J.W. Vector competence of North American mosquitoes (Diptera: Culicidae) for West Nile virus. J. Med. Entomol. 2001, 38, 130–134. [Google Scholar] [CrossRef]
- Anderson, S.L.; Richards, S.L.; Tabachnick, W.J.; Smartt, C.T. Effects of West Nile virus dose and extrinsic incubation temperature on temporal progression of vector competence in Culex pipiens quinquefasciatus. J. Am. Mosq. Control. Assoc. 2010, 26, 103–107. [Google Scholar] [CrossRef]
- Vogels, C.B.; Goertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector competence of European mosquitoes for West Nile virus. Emerg. Microbes Infect. 2017, 6, e96. [Google Scholar] [CrossRef] [Green Version]
- Heitmann, A.; Jansen, S.; Luhken, R.; Helms, M.; Pluskota, B.; Becker, N.; Kuhn, C.; Schmidt-Chanasit, J.; Tannich, E. Experimental risk assessment for chikungunya virus transmission based on vector competence, distribution and temperature suitability in Europe, 2018. Eurosurveillance 2018, 23, 1800033. [Google Scholar] [CrossRef]
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect. Genet. Evol. 2019, 67, 191–209. [Google Scholar] [CrossRef]
- Meyer, R.P.; Hardy, J.L.; Reisen, W.K. Diel changes in adult mosquito microhabitat temperatures and their relationship to the extrinsic incubation of arboviruses in mosquitoes in Kern County, California. J. Med. Entomol. 1990, 27, 607–614. [Google Scholar] [CrossRef]
- Alto, B.W.; Wiggins, K.; Eastmond, B.; Ortiz, S.; Zirbel, K.; Lounibos, L.P. Diurnal Temperature Range and Chikungunya Virus Infection in Invasive Mosquito Vectors. J. Med. Entomol. 2018, 55, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Paaijmans, K.P.; Fansiri, T.; Carrington, L.B.; Kramer, L.D.; Thomas, M.B.; Scott, T.W. Impact of daily temperature fluctuations on dengue virus transmission by Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 108, 7460–7465. [Google Scholar] [CrossRef] [Green Version]
- Carrington, L.B.; Seifert, S.N.; Armijos, M.V.; Lambrechts, L.; Scott, T.W. Reduction of Aedes aegypti vector competence for dengue virus under large temperature fluctuations. Am. J. Trop. Med. Hyg. 2013, 88, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paaijmans, K.P.; Blanford, S.; Bell, A.S.; Blanford, J.I.; Read, A.F.; Thomas, M.B. Influence of climate on malaria transmission depends on daily temperature variation. Proc. Natl. Acad. Sci. USA 2010, 107, 15135–15139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention. West Nile Virus-Final Cumulative Maps & Data for 1999–2019. Available online: https://www.cdc.gov/westnile/statsmaps/cumMapsData.html (accessed on 23 March 2021).
- U.S. Climate Data. Available online: https://www.usclimatedata.com/climate/united-states/us (accessed on 18 October 2019).
- Shocket, M.S.; Verwillow, A.B.; Numazu, M.G.; Slamani, H.; Cohen, J.M.; El Moustaid, F.; Rohr, J.; Johnson, L.R.; Mordecai, E.A. Transmission of West Nile and five other temperate mosquito-borne viruses peaks at temperatures between 23 degrees C and 26 degrees C. Elife 2020, 9, e58511. [Google Scholar] [CrossRef]
- Anderson, S.L.; Richards, S.L.; Smartt, C.T. A simple method for determining arbovirus transmission in mosquitoes. J. Am. Mosq. Control. Assoc. 2010, 26, 108–111. [Google Scholar] [CrossRef] [Green Version]
- McGregor, B.L.; Connelly, C.R.; Kenney, J.L. Infection, Dissemination, and Transmission Potential of North American Culex quinquefasciatus, Culex tarsalis, and Culicoides sonorensis for Oropouche Virus. Viruses 2021, 13, 226. [Google Scholar] [CrossRef]
- Girard, Y.A.; Klingler, K.A.; Higgs, S. West Nile virus dissemination and tissue tropisms in orally infected Culex pipiens quinquefasciatus. Vector-Borne Zoonotic Dis. 2004, 4, 109–122. [Google Scholar] [CrossRef]
- Goddard, L.B.; Roth, A.E.; Reisen, W.K.; Scott, T.W. Vector competence of California mosquitoes for West Nile virus. Emerg. Infect. Dis. 2002, 8, 1385–1391. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K.; Fang, Y.; Martinez, V.M. Effects of temperature on the transmission of West Nile virus by Culex tarsalis (Diptera: Culicidae). J. Med. Entomol. 2014, 43, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Cornel, A.J.; Jupp, P.G.; Blackburn, N.K. Environmental temperature on the vector competence of Culex univittatus (Diptera: Culicidae) for West Nile Virus. J. Med. Entomol. 1993, 30, 449–456. [Google Scholar] [CrossRef]
- Kramer, L.D.; Hardy, J.L.; Presser, S.B. Effect of temperature of extrinsic incubation on the vector competence of Culex tarsalis for Western equine encephalomyelitis virus. Am. J. Trop. Med. Hyg. 1983, 32, 1130–1139. [Google Scholar] [CrossRef]
- Paz, S. Climate change impacts on West Nile virus transmission in a global context. Philos. Trans. R. Soc. B 2015, 370, 20130561. [Google Scholar] [CrossRef]
- Braganza, K.; Karoly, D.J.; Arblaster, J.M. Diurnal temperature range as an index of global climate change during the twentieth century. Geophys. Res. Lett. 2004, 31, 1–4. [Google Scholar] [CrossRef]
- Lobell, D.B.; Bonfils, C.; Duffy, P.B. Climate change uncertainty for daily minimum and maximum temperatures: A model inter-comparison. Geophys. Res. Lett. 2007, 34. [Google Scholar] [CrossRef] [Green Version]
- Yee, W.L.; Foster, W.A. Diel sugar-feeding and host-seeking rhythms in mosquitoes (Diptera: Culicidae) under laboratory conditions. J. Med. Entomol. 1992, 29, 784–791. [Google Scholar] [CrossRef]
- Reisen, W.K.; Lothrop, H.D.; Meyer, R.P. Time of host-seeking by Culex tarsalis (Diptera: Culicidae) in California. J. Med. Entomol. 1997, 34, 430–437. [Google Scholar] [CrossRef]
- Godsey, M.S.G., Jr.; Burkhalter, K.; Delorey, M.; Savage, H.M. Seasonality and time of host-seeking activity of Culex tarsalis and floodwater Aedes in northern Colorado, 2006–2007. J. Am. Mosq. Control. Assoc. 2010, 26, 148–159. [Google Scholar]
- Savage, H.M.; Anderson, M.; Gordon, E.; McMillen, L.; Colton, L.; Delorey, M.; Sutherland, G.; Aspen, S.; Charnetzky, D.; Burkhalter, K.; et al. Host-seeking heights, host-seeking activity patterns, and West Nile virus infection rates for members of the Culex pipiens complex at different habitat types within the Hybrid Zone, Shelby County, TN, 2002 (Diptera: Culicidae). J. Med. Entomol. 2008, 45, 276–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Culex Species | Trial | Time (DPI) | Treatment | Bodies † | Bodies p | Legs † | Legs p | Saliva † | Saliva p |
|---|---|---|---|---|---|---|---|---|---|
| quinquefasciatus | 1 | 4 | Static | 29/31 | 0.255 | 2/29 | 1 | 1/2 | 1 |
| Fluctuating | 25/31 | 2/25 | 2/2 | ||||||
| 8 | Static | 28/32 | 1 | 9/28 | 0.781 | 1/9 | 0.450 | ||
| Fluctuating | 28/31 | 11/28 | 0/11 | ||||||
| 2 | 4 | Static | 69/89 | 0.319 | 4/69 | 1 | 0/4 | 1 | |
| Fluctuating | 63/90 | 3/63 | 0/3 | ||||||
| 8 | Static | 62/90 | 0.622 | 10/62 | 0.622 | 2/10 | 1 | ||
| Fluctuating | 66/90 | 10/66 | 1/10 | ||||||
| 12 | Static | 76/113 | 0.241 | 39/76 | 1 | 10/39 | 0.219 | ||
| Fluctuating | 51/87 | 25/51 | 2/25 | ||||||
| tarsalis | 1 | 4 | Static | 19/23 | 0.109 | 11/19 | 0.213 | 4/11 | 0.338 |
| Fluctuating | 23/23 | 8/23 | 1/8 | ||||||
| 8 | Static | 24/25 | 1 | 22/24 | 0.489 | 20/22 | 0.223 | ||
| Fluctuating | 24/26 | 24/24 | 24/24 | ||||||
| 2 | 4 | Static | 42/63 | 0.714 | 7/42 | 0.757 | 1/7 | 1 | |
| Fluctuating | 41/65 | 5/41 | 1/5 | ||||||
| 8 | Static | 19/41 | 0.691 | 15/19 | 0.456 | 10/15 | 0.716 | ||
| Fluctuating | 28/68 | 25/28 | 19/25 |
| Culex Species | Trial | Time (DPI) | Treatment | Body Titer | Body p | Leg Titer | Leg p | Saliva Titer | Saliva p |
|---|---|---|---|---|---|---|---|---|---|
| quinquefasciatus | 1 | 4 | Static | 5.24 | 0.574 | 1.48 | 0.333 | 0.82 | n/a |
| Fluctuating | 5.20 | 0.92 | 0.82 | ||||||
| 8 | Static | 6.18 | 0.054 | 4.17 | 0.619 | 4.69 | n/a | ||
| Fluctuating | 5.82 | 3.90 | n/a | ||||||
| 2 | 4 | Static | 6.49 | 0.421 | 1.18 | 0.270 | n/a | n/a | |
| Fluctuating | 5.55 | 0.82 | n/a | ||||||
| 8 | Static | 7.02 | <0.001 | 4.20 | 0.791 | 2.15 | 0.667 | ||
| Fluctuating | 6.39 | 4.17 | 1.00 | ||||||
| 12 | Static | 7.62 | 0.007 | 4.60 | 0.821 | 2.49 | 0.075 | ||
| Fluctuating | 6.92 | 4.18 | 1.00 | ||||||
| tarsalis | 1 | 4 | Static | 6.71 | 0.062 | 3.47 | 0.342 | 1.00 | n/a |
| Fluctuating | 6.84 | 3.19 | 0.82 | ||||||
| 8 | Static | 7.81 | <0.001 | 5.01 | 0.008 | 3.67 | 0.191 | ||
| Fluctuating | 8.15 | 5.44 | 4.11 | ||||||
| 2 | 4 | Static | 5.46 | 0.068 | 4.25 | 0.222 | 1.70 | n/a | |
| Fluctuating | 6.97 | 4.66 | 2.99 | ||||||
| 8 | Static | 7.95 | 0.025 | 5.07 | 0.665 | 3.18 | 0.854 | ||
| Fluctuating | 8.23 | 5.06 | 4.79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McGregor, B.L.; Kenney, J.L.; Connelly, C.R. The Effect of Fluctuating Incubation Temperatures on West Nile Virus Infection in Culex Mosquitoes. Viruses 2021, 13, 1822. https://doi.org/10.3390/v13091822
McGregor BL, Kenney JL, Connelly CR. The Effect of Fluctuating Incubation Temperatures on West Nile Virus Infection in Culex Mosquitoes. Viruses. 2021; 13(9):1822. https://doi.org/10.3390/v13091822
Chicago/Turabian StyleMcGregor, Bethany L., Joan L. Kenney, and C. Roxanne Connelly. 2021. "The Effect of Fluctuating Incubation Temperatures on West Nile Virus Infection in Culex Mosquitoes" Viruses 13, no. 9: 1822. https://doi.org/10.3390/v13091822
APA StyleMcGregor, B. L., Kenney, J. L., & Connelly, C. R. (2021). The Effect of Fluctuating Incubation Temperatures on West Nile Virus Infection in Culex Mosquitoes. Viruses, 13(9), 1822. https://doi.org/10.3390/v13091822