
Modulation of HIV Replication in Monocyte-Derived Macrophages (MDM) by Host Antiviral Factors Secretory Leukocyte Protease Inhibitor and Serpin Family C Member 1 Induced by Steroid Hormones

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Isolation and Culture of Monocyte-Derived Macrophages (MDMs)
2.3. Cell Lines and Culture Conditions
2.4. HIV-1 Infection
2.5. RNA Isolation and Real-Time Quantitative PCR Array
2.6. In Vitro HIV-1 Inhibition Assay
- (1)
- Pre-treatment treatment assay- MDMs (0.5 × 106 cells/well in a 24-well plate) were pre-treated with SERPIN C1 or SLPI and then infected with HIV-1 BaL (5 ng p24/mL/106 cells). After 2 h of virus adsorption, culture medium was replaced with SERPIN C1 (0.05 µg/mL and 0.1 µg/mL) or SLPI (1 µg/mL and 10 µg/mL) containing DMEM culture medium. MDMs were incubated for 7 days at 37 °C under 5% CO2.).
- (2)
- Post treatment assay-HIV-1 BaL (5 ng p24/mL/106 cells) infected MDMs (0.5 × 106 cells/well in a 24-well plate) were treated with SERPIN C1 (0.05 µg/mL and 0.1 µg/mL) or SLPI (1 µg/mL and 10 µg/mL) post infection and cultured for 7 days.
- (3)
- Inactivation of HIV-1- HIV-1 BaL (5 ng/mL p24 units) was incubated without or with SLPI (1 µg/mL and 10 µg/mL) or SERPIN C1 (0.05 µg/mL and 0.1 µg/mL) at 37 °C for 2 h in PBS containing 0.1% BSA. After that, MDMs (5 × 105 cells/well) were infected with HIV-1 BaL and incubated at 37 °C for 2 h for virus adsorption washed with phosphate-buffered saline (PBS) and cultured in 2 mL fresh media at 37 °C for 7 days.
2.7. HIV-1 Env Pseudo Typed Viral Production and Infection
2.8. Quantitation of HIV-1 Replication
2.9. SERPIN C1 and SLPI Assay
2.10. Detection of Integrated HIV-1 DNA, Early and Late Stage HIV-1 DNA Products
2.11. In Trans HIV-1 Transmission Assay
2.12. Statistical Analysis
3. Results and Discussion
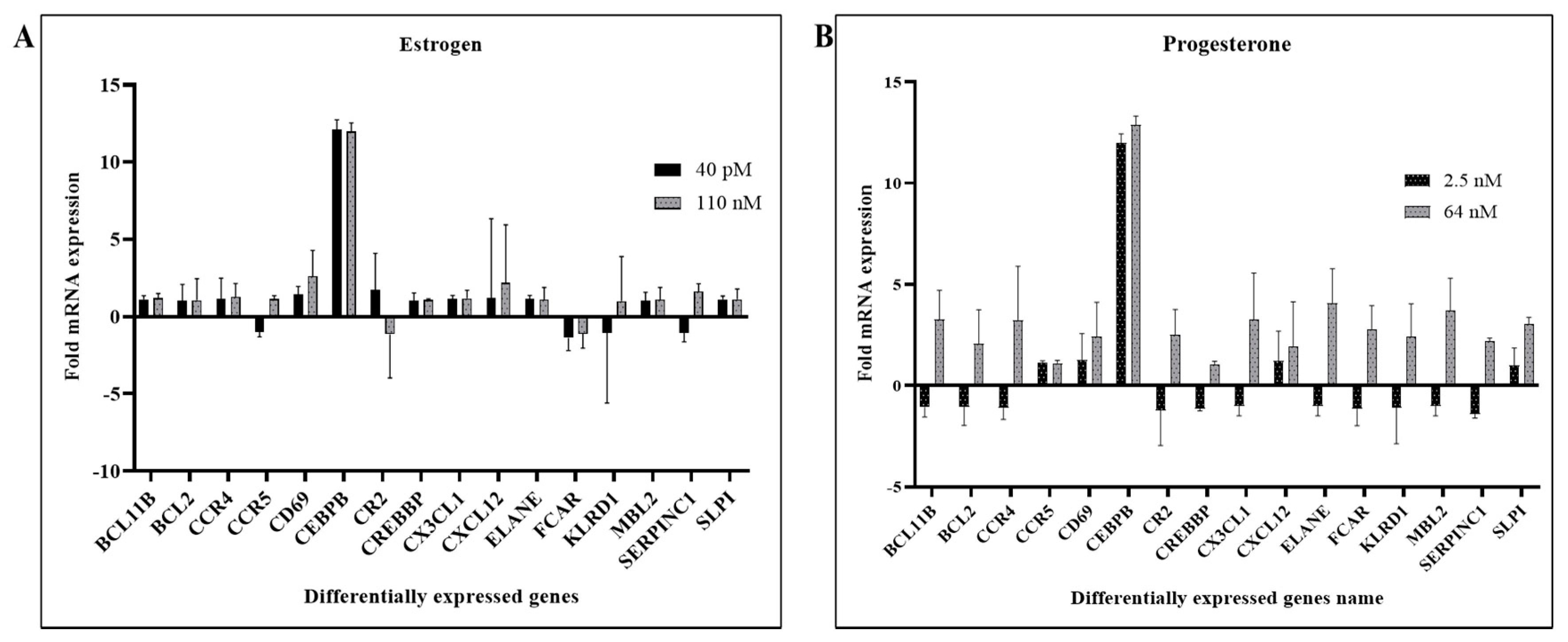
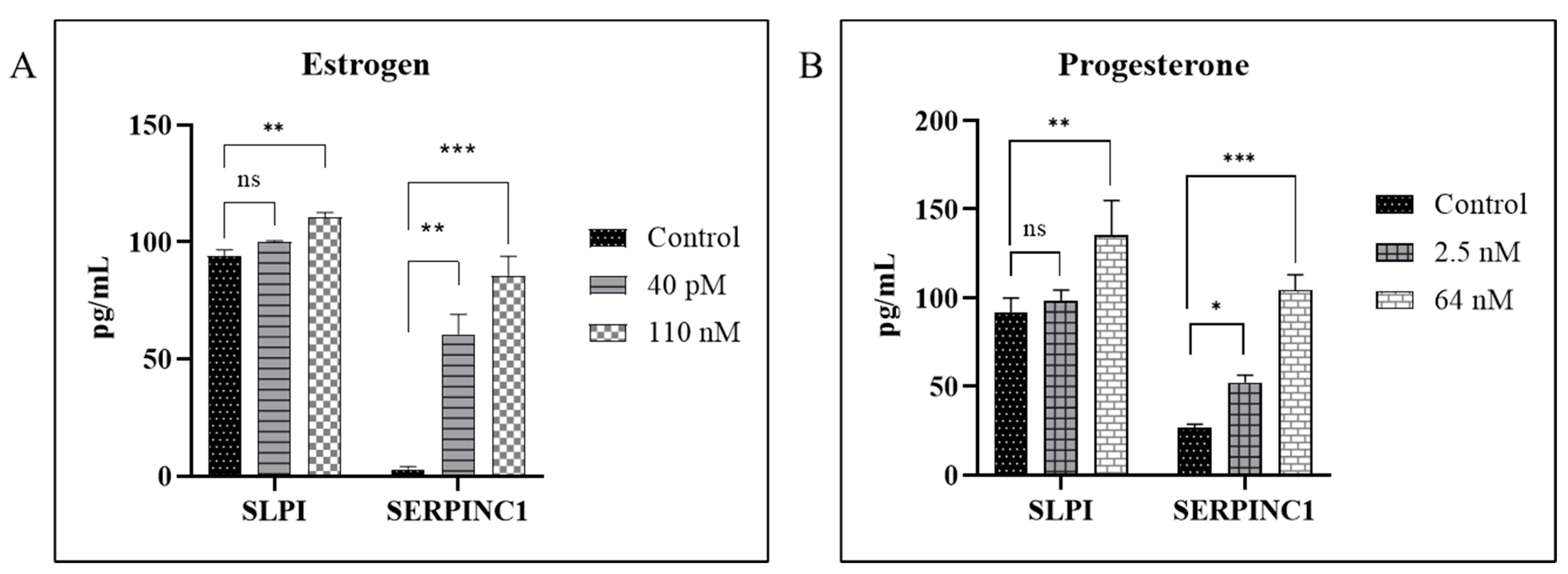
3.1. Hormones Induces SERPIN C1 and SLPI in HIV-1-Infected MDMs
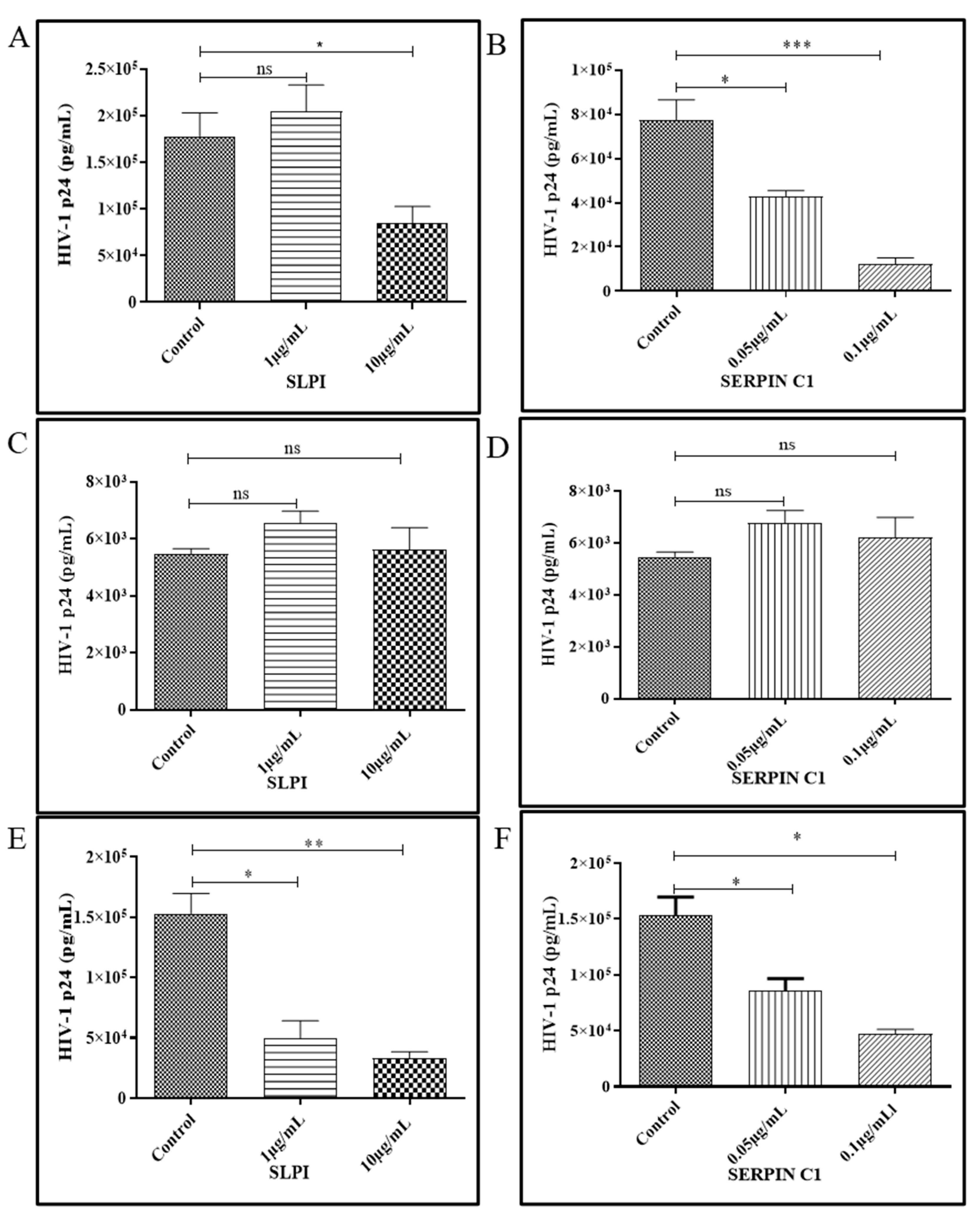
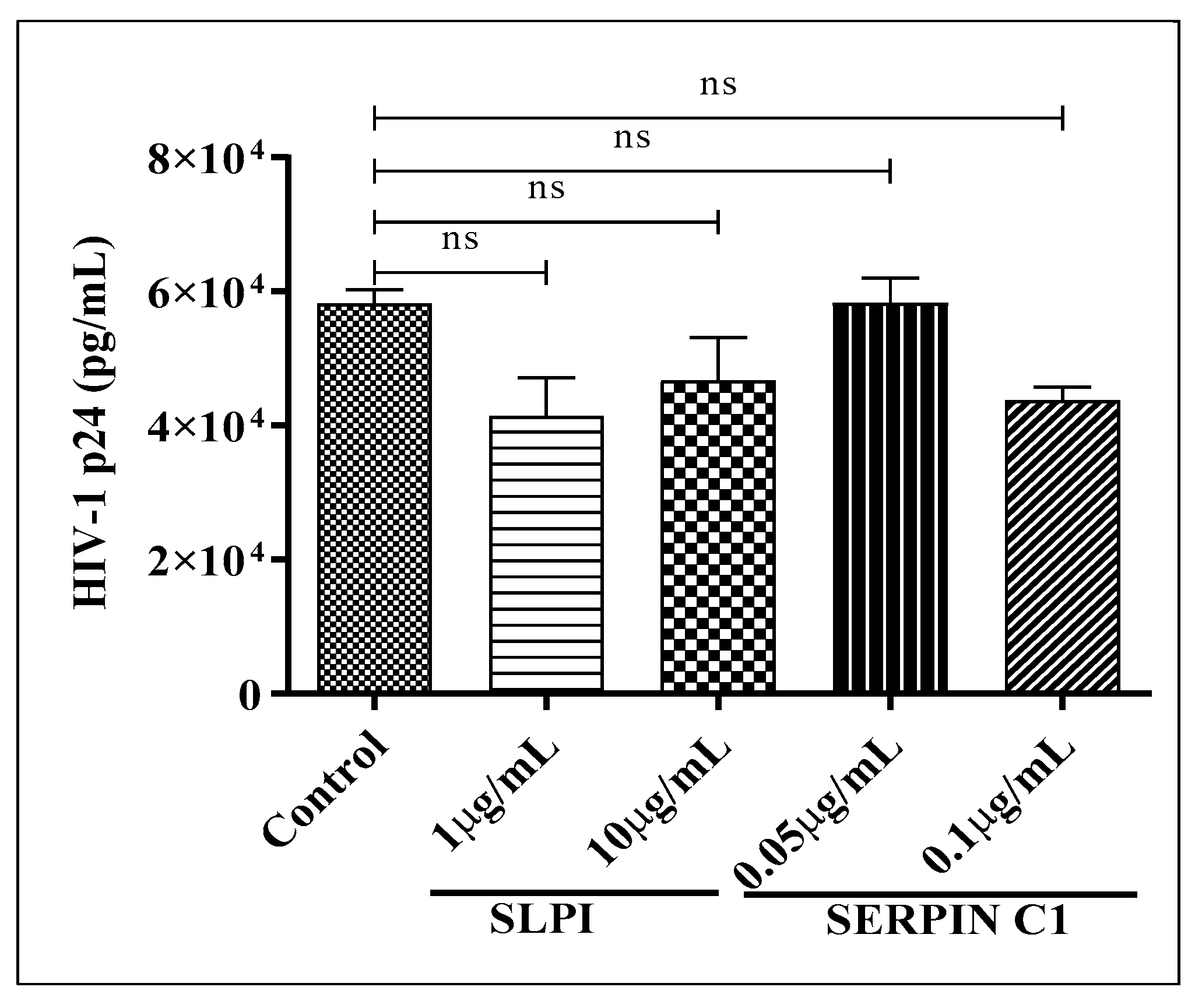
3.2. Human Recombinant Protein SERPIN C1 and SLPI Modulates HIV-1 Replication
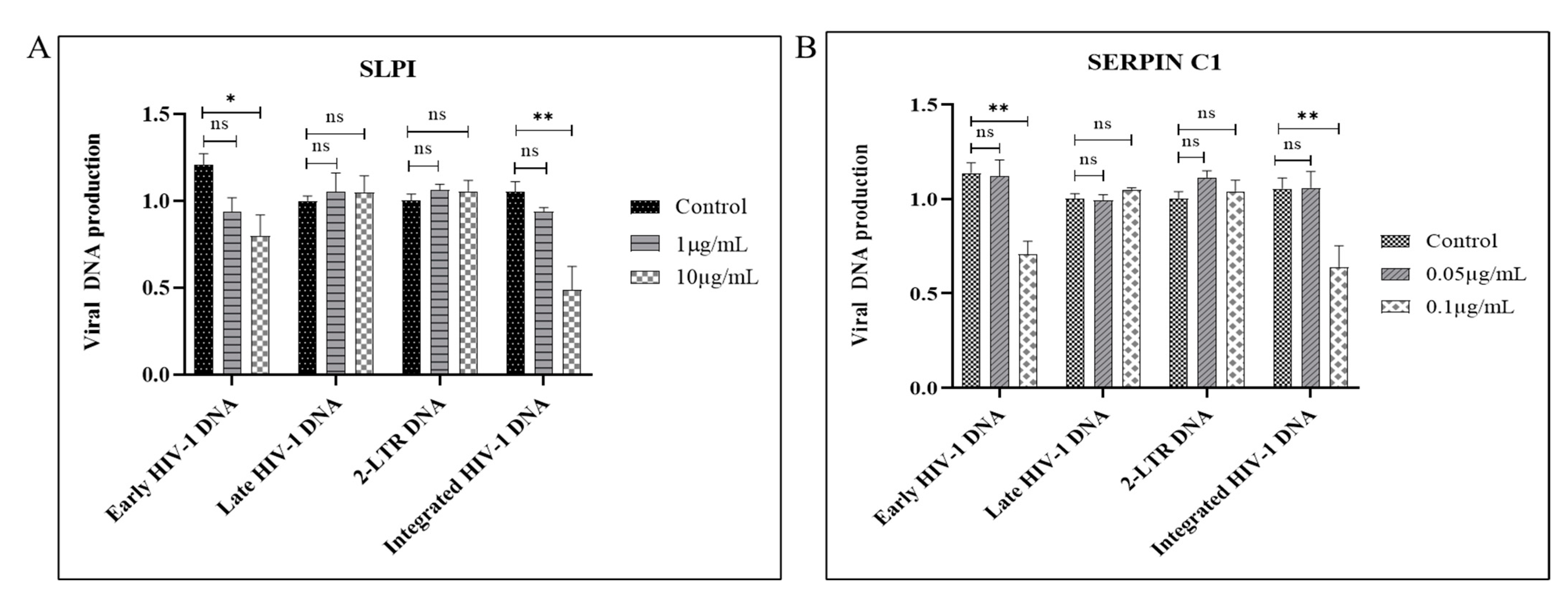
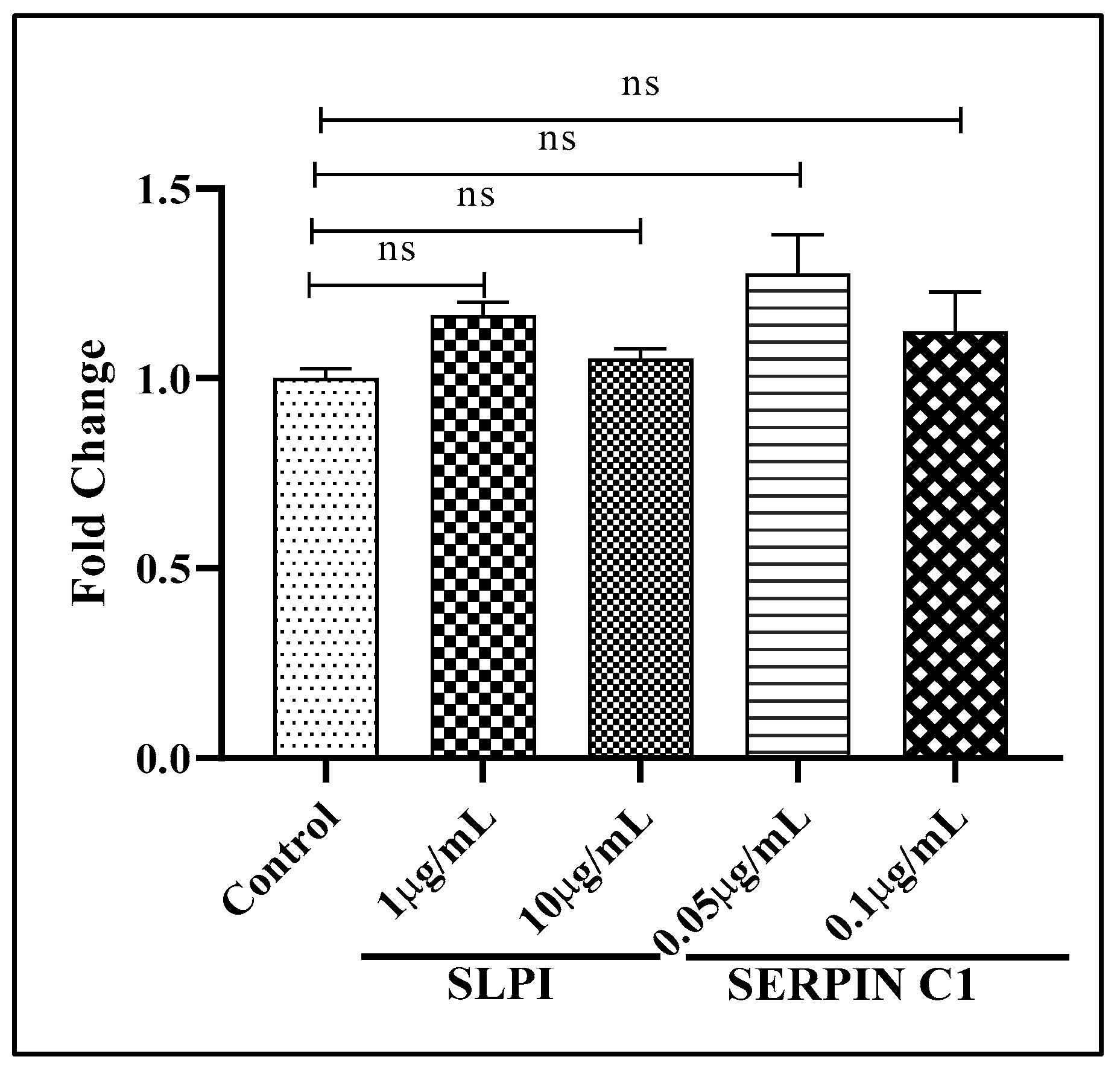
3.3. SERPIN C1 or SLPI Reduces the Level of Integrated HIV-1 DNA
3.4. Impact of SERPIN C1 or SLPI on HIV-1 Replication Post Integration
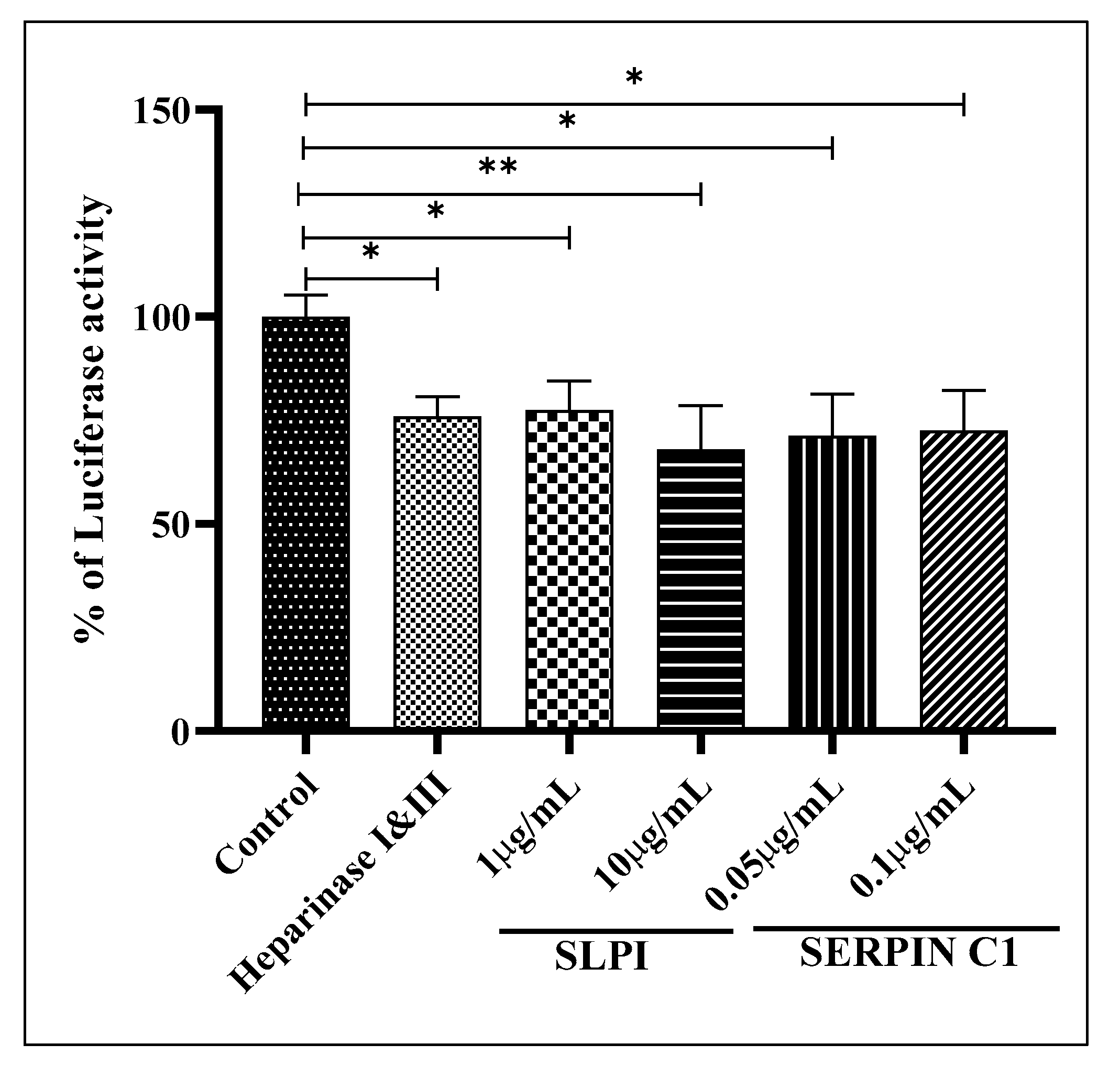
3.5. SERPIN C1 or SLPI Impede HIV-1 Entry into Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
Additional Information
References
- Marcus, J.L.; Leyden, W.A.; Alexeeff, S.E.; Anderson, A.N.; Hechter, R.C.; Hu, H.; Lam, J.O.; Towner, W.J.; Yuan, Q.; Horberg, M.A.; et al. Comparison of Overall and Comorbidity-Free Life Expectancy Between Insured Adults with and without HIV Infection, 2000-2016. JAMA Netw. Open 2020, 3, e207954. [Google Scholar] [CrossRef]
- CDC. NCHHSTP Atlas Plus. Available online: https://www.cdc.gov/nchhstp/atlas/index.htm (accessed on 31 March 2020).
- Long, E.M.; Martin, H.L., Jr.; Kreiss, J.K.; Rainwater, S.M.; Lavreys, L.; Jackson, D.J.; Rakwar, J.; Mandaliya, K.; Overbaugh, J. Gender differences in HIV-1 diversity at time of infection. Nat. Med. 2000, 6, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Anastos, K.; Gange, S.J.; Lau, B.; Weiser, B.; Detels, R.; Giorgi, J.V.; Margolick, J.B.; Cohen, M.; Phair, J.; Melnick, S.; et al. Association of race and gender with HIV-1 RNA levels and immunologic progression. J. Acquir. Immune Defic. Syndr. 2000, 24, 218–226. [Google Scholar] [CrossRef]
- Addo, M.M.; Altfeld, M. Sex-Based Differences in HIV Type 1 Pathogenesis. J. Infect. Dis. 2014, 209, S86–S92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griesbeck, M.; Scully, E.; Altfeld, M. Sex and gender differences in HIV-1 infection. Clin. Sci. 2016, 130, 1435–1451. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, M.; Bacchetti, P.; Miotti, P.; Quinn, T.C.; Veronese, F.; Greenblatt, R.M. Does Patient Sex Affect Human Immunodeficiency Virus Levels? Clin. Infect. Dis. 2002, 35, 313–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S. The effects of hormones on sex differences in infection: From genes to behavior. Neurosci. Biobehav. Rev. 2000, 24, 627–638. [Google Scholar] [CrossRef]
- Fish, E.N. The X-files in immunity: Sex-based differences predispose immune responses. Nat. Rev. Immunol. 2008, 8, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Asin, S.N.; Eszterhas, S.K.; Rollenhagen, C.; Heimberg, A.M.; Howell, A.L. HIV Type 1 Infection in Women: Increased Transcription of HIV Type 1 in Ectocervical Tissue Explants. J. Infect. Dis. 2009, 200, 965–972. [Google Scholar] [CrossRef]
- Asin, S.N.; Fanger, M.W.; Wildt-Perinic, D.; Ware, P.L.; Wira, C.R.; Howell, A.L. Transmission of HIV-1 by Primary Human Uterine Epithelial Cells and Stromal Fibroblasts. J. Infect. Dis. 2004, 190, 236–245. [Google Scholar] [CrossRef]
- Zhang, M.; Huang, Q.; Huang, Y.; Wood, O.; Yuan, W.; Chancey, C.; Daniel, S.; Rios, M.; Hewlett, I.; Clouse, A.K.; et al. beta-estradiol attenuates the anti-HIV-1 efficacy of Stavudine (D4T) in primary PBL. Retrovirology 2008, 5, 82. [Google Scholar] [CrossRef] [Green Version]
- Vishwanathan, A.S.; Guenthner, P.C.; Lin, C.Y.; Dobard, C.; Sharma, S.; Adams, D.R.; Otten, A.R.; Heneine, W.; Hendry, R.M.; McNicholl, J.M.; et al. High Susceptibility to Repeated, Low-Dose, Vaginal SHIV Exposure Late in the Luteal Phase of the Menstrual Cycle of Pigtail Macaques. JAIDS J. Acquir. Immune Defic. Syndr. 2011, 57, 261–264. [Google Scholar] [CrossRef]
- Ragupathy, V.; Devadas, K.; Tang, S.; Wood, O.; Lee, S.; Dastyer, A.; Wang, X.; Dayton, A.; Hewlett, I. Effect of sex steroid hormones on replication and transmission of major HIV subtypes. J. Steroid Biochem. Mol. Biol. 2013, 138, 63–71. [Google Scholar] [CrossRef]
- Rodríguez-García, M.; Biswas, N.; Patel, M.V.; Barr, F.D.; Crist, S.G.; Ochsenbauer, C.; Fahey, J.V.; Wira, C.R. Estradiol Reduces Susceptibility of CD4+ T Cells and Macrophages to HIV-Infection. PLoS ONE 2013, 8, e62069. [Google Scholar] [CrossRef] [Green Version]
- Hagen, S.; Altfeld, M. The X awakens: Multifactorial ramifications of sex-specific differences in HIV-1 infection. J. Virus Erad. 2016, 2, 78–81. [Google Scholar] [CrossRef]
- Ragupathy, V.; Xue, W.; Tan, J.; Devadas, K.; Gao, Y.; Hewlett, I. Progesterone augments cell susceptibility to HIV-1 and HIV-1/HSV-2 co-infections. J. Mol. Endocrinol. 2016, 57, 185–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngcapu, S.; Masson, L.; Sibeko, S.; Werner, L.; McKinnon, L.; Mlisana, K.; Shey, M.; Samsunder, N.; Karim, S.A.; Karim, Q.A.; et al. Lower concentrations of chemotactic cytokines and soluble innate factors in the lower female genital tract associated with the use of injectable hormonal contraceptive. J. Reprod. Immunol. 2015, 110, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannoni, E.; Guignard, L.; Reymond, M.K.; Perreau, M.; Roth-Kleiner, M.; Calandra, T.; Roger, T. Estradiol and Progesterone Strongly Inhibit the Innate Immune Response of Mononuclear Cells in Newborns. Infect. Immun. 2011, 79, 2690–2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verthelyi, D. Sex hormones as immunomodulators in health and disease. Int. Immunopharmacol. 2001, 1, 983–993. [Google Scholar] [CrossRef]
- Boucher, A.; Mourad, W.; Mailloux, J.; Lemay, A.; Akoum, A. Ovarian hormones modulate monocyte chemotactic protein-1 expression in endometrial cells of women with endometriosis. Mol. Hum. Reprod. 2000, 6, 618–626. [Google Scholar] [CrossRef] [Green Version]
- Tasker, C.; Ding, J.; Schmolke, M.; Rivera-Medina, A.; Garcia-Sastre, A.; Chang, T.L. 17β-Estradiol Protects Primary Macrophages Against HIV Infection Through Induction of Interferon-Alpha. Viral Immunol. 2014, 27, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Rollenhage, C.; Macura, S.L.; Lathrop, M.J.; MacKenzie, T.A.; Doncel, G.F.; Asin, S.N. Enhancing Interferon Regulatory Factor 7 Mediated Antiviral Responses and Decreasing Nuclear Factor Kappa B Expression Limit HIV-1 Replication in Cervical Tissues. PLoS ONE 2015, 10, e0131919. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Sun, Y.; Ma, F.; Lü, P.; Huang, H.; Zhou, J. Progesterone inhibits Toll-like receptor 4-mediated innate immune response in macrophages by suppressing NF-κB activation and enhancing SOCS1 expression. Immunol. Lett. 2009, 125, 151–155. [Google Scholar] [CrossRef]
- Sun, Y.; Cai, J.; Ma, F.; Lü, P.; Huang, H.; Zhou, J. miR-155 mediates suppressive effect of progesterone on TLR3, TLR4-triggered immune response. Immunol. Lett. 2012, 146, 25–30. [Google Scholar] [CrossRef]
- Asin, S.N.; Heimberg, A.M.; Eszterhas, S.K.; Rollenhagen, C.; Howell, A.L. Estradiol and Progesterone Regulate HIV Type 1 Replication in Peripheral Blood Cells. AIDS Res. Hum. Retroviruses 2008, 24, 701–716. [Google Scholar] [CrossRef] [PubMed]
- Andany, N.; Kennedy, V.L.; Aden, M.; Loutfy, M. Perspectives on menopause and women with HIV. Int. J. Women’s Health 2016, 8, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Wira, C.R.; Rodriguez-Garcia, M.; Patel, M.V. The role of sex hormones in immune protection of the female reproductive tract. Nat. Rev. Immunol. 2015, 15, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Rollenhagen, C.; Asin, S.N. Enhanced HIV-1 replication in ex vivo ectocervical tissues from post-menopausal women correlates with increased inflammatory responses. Mucosal Immunol. 2011, 4, 671–681. [Google Scholar] [CrossRef]
- Ghosh, M.; Jais, M.; Delisle, J.; Younes, N.; Benyeogor, I.; Biswas, R.; Mohamed, H.; Daniels, J.; Wang, C.; Young, M.; et al. Dysregulation in Genital Tract Soluble Immune Mediators in Postmenopausal Women Is Distinct by HIV Status. AIDS Res. Hum. Retroviruses 2019, 35, 251–259. [Google Scholar] [CrossRef]
- Devadas, K.; Biswas, S.; Ragupathy, V.; Lee, S.; Dayton, A.; Hewlett, I. Modulation of HIV replication in monocyte derived macrophages (MDM) by steroid hormones. PLoS ONE 2018, 13, e0191916. [Google Scholar] [CrossRef] [Green Version]
- Feistritzer, C.; Wiedermann, C.J. Effects of anticoagulant strategies on activation of inflammation and coagulation. Expert Opin. Biol. Ther. 2007, 7, 855–870. [Google Scholar] [CrossRef]
- Opal, S.M.; Esmon, C.T. Bench-to-bedside review: Functional relationships between coagulation and the innate immune response and their respective roles in the pathogenesis of sepsis. Crit. Care 2002, 7, 23–38. [Google Scholar] [CrossRef] [Green Version]
- McNeely, T.B.; Dealy, M.; Dripps, D.J.; Orenstein, J.M.; Eisenberg, S.P.; Wahl, S.M. Secretory leukocyte protease inhibitor: A human saliva protein exhibiting anti-human immunodeficiency virus 1 activity in vitro. J. Clin. Investig. 1995, 96, 456–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgener, A.; Boutilier, J.; Wachihi, C.; Kimani, J.; Carpenter, M.; Westmacott, G.; Cheng, K.; Ball, T.B.; Plummer, F. Identification of Differentially Expressed Proteins in the Cervical Mucosa of HIV-1-Resistant Sex Workers. J. Proteome Res. 2008, 7, 4446–4454. [Google Scholar] [CrossRef]
- Geiben-Lynn, R.; Brown, N.; Walker, B.D.; Luster, A.D. Purification of a Modified Form of Bovine Antithrombin III as an HIV-1 CD8+ T-cell Antiviral Factor. J. Biol. Chem. 2002, 277, 42352–42357. [Google Scholar] [CrossRef] [Green Version]
- Wahl, S.M.; McNeely, T.B.; Janoff, E.N.; Shugars, D.; Worley, P.; Tucker, C.; Orenstein, J.M. Secretory leukocyte protease inhibitor (SLPI) in mucosal fluids inhibits HIV-1. Oral Dis. 1997, 3, S64–S69. [Google Scholar] [CrossRef]
- Wahl, S.M.; Worley, P.; Jin, W.; McNeely, T.B.; Eisenberg, S.; Fasching, C.; Orenstein, J.M.; Janoff, E.N. Anatomic dissociation between HIV-1 and its endogenous inhibitor in mucosal tissues. Am. J. Pathol. 1997, 150, 1275–1284. [Google Scholar]
- Zhang, Y.; DeWitt, D.L.; McNeely, T.B.; Wahl, S.M.; Wahl, L.M. Secretory leukocyte protease inhibitor suppresses the production of monocyte prostaglandin H synthase-2, prostaglandin E2, and matrix metalloproteinases. J. Clin. Investig. 1997, 99, 894–900. [Google Scholar] [CrossRef] [Green Version]
- McNeely, T.B.; Shugars, D.C.; Rosendahl, M.; Tucker, C.; Eisenberg, S.P.; Wahl, S.M. Inhibition of human immunodeficiency virus type 1 infectivity by secretory leukocyte protease inhibitor occurs prior to viral reverse transcription. Blood 1997, 90, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, C.; VanCott, T.C.; Mbori-Ngacha, R.A.; Horani, L.; Bosire, R.K.; Kreiss, J.K.; Richardson, B.A.; John-Stewart, G.C. Salivary Secretory Leukocyte Protease Inhibitor Is Associated with Reduced Transmission of Human Immunodeficiency Virus Type 1 through Breast Milk. J. Infect. Dis. 2002, 186, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Geiben-Lynn, R.; Kursar, M.; Brown, N.V.; Kerr, E.L.; Luster, A.D.; Walker, B.D. Noncytolytic Inhibition of X4 Virus by Bulk CD8 + Cells from Human Immunodeficiency Virus Type 1 (HIV-1)-Infected Persons and HIV-1-Specific Cytotoxic T Lymphocytes Is Not Mediated by β-Chemokines. J. Virol. 2001, 75, 8306–8316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitney, J.B.; Asmal, M.; Geiben-Lynn, R. Serpin Induced Antiviral Activity of Prostaglandin Synthetase-2 against HIV-1 Replication. PLoS ONE 2011, 6, e18589. [Google Scholar] [CrossRef] [Green Version]
- Brabin, L. Interactions of the Female Hormonal Environment, Susceptibility to Viral Infections, and Disease Progression. AIDS Patient Care STDs 2002, 16, 211–221. [Google Scholar] [CrossRef]
- Hel, Z.; Stringer, E.; Mestecky, J. Sex Steroid Hormones, Hormonal Contraception, and the Immunobiology of Human Immunodeficiency Virus-1 Infection. Endocr. Rev. 2009, 31, 79–97. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.P.; Klein, S.L. Pregnancy and pregnancy-associated hormones alter immune responses and disease pathogenesis. Horm. Behav. 2012, 62, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Brotman, R.M.; Ravel, J.; Bavoil, P.M.; Gravitt, P.E.; Ghanem, K.G. Microbiome, sex hormones, and immune responses in the reproductive tract: Challenges for vaccine development against sexually transmitted infections. Vaccine 2013, 32, 1543–1552. [Google Scholar] [CrossRef] [Green Version]
- Ochiel, D.O.; Fahey, J.V.; Ghosh, M.; Haddad, S.N.; Wira, C.R. Innate Immunity in the Female Reproductive Tract: Role of Sex Hormones in Regulating Uterine Epithelial Cell Protection Against Pathogens. Curr. Womens Health Rev. 2008, 4, 102–117. [Google Scholar] [CrossRef] [Green Version]
- Solar, P.; Velásquez, L. Consequences of nongenomic actions of estradiol on pathogenic genital tract response. J. Mol. Signal. 2013, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gartner, S.; Markovits, P.; Markovitz, D.M.; Kaplan, M.H.; Gallo, R.C.; Popovic, M. The Role of Mononuclear Phagocytes in HTLV-III/LAV Infection. Science 1986, 233, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Nonnemacher, M.R.; Wigdahl, B. CCAAT/enhancer-binding proteins and the pathogenesis of retrovirus infection. Futur. Microbiol. 2009, 4, 299–321. [Google Scholar] [CrossRef] [Green Version]
- Wamel, J.L.van; Berkhout, B. The First Strand Transfer during HIV-1 Reverse Transcription Can Occur either Intramolecularly or Intermolecularly. Virology 1998, 244, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Peliska, J.A.; Benkovic, S.J. Mechanism of DNA strand transfer reactions catalyzed by HIV-1 reverse transcriptase. Science 1992, 258, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Bukrinsky, M.I.; Sharova, N.; Dempsey, M.P.; Stanwick, T.L.; Bukrinskaya, A.G.; Haggerty, S.; Stevenson, M. Active nuclear import of human immunodeficiency virus type 1 preintegration complexes. Proc. Natl. Acad. Sci. USA 1992, 89, 6580–6584. [Google Scholar] [CrossRef] [Green Version]
- Shugars, D.; Sauls, D.; Weinberg, J. Secretory leukocyte protease inhibitor blocks infectivity of primary monocytes and mononuclear cells with both monocytotropic and lymphocytotropic strains of human immunodeficiency virus type 1. Oral Dis. 1997, 3, S70–S72. [Google Scholar] [CrossRef] [PubMed]
- Afratis, N.; Nikitovic, D.; Multhaupt, H.A.B.; Theocharis, A.D.; Couchman, J.R.; Karamanos, N.K. Syndecans—key regulators of cell signaling and biological functions. FEBS J. 2017, 284, 27–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saphire, A.C.S.; Bobardt, M.D.; Zhang, Z.; David, G.; Gallay, P.A. Syndecans Serve as Attachment Receptors for Human Immunodeficiency Virus Type 1 on Macrophages. J. Virol. 2001, 75, 9187–9200. [Google Scholar] [CrossRef] [Green Version]
- Bobardt, M.D.; Saphire, A.C.; Hung, H.-C.; Yu, X.; Van der Schueren, B.; Zhang, Z.; David, G.; Gallay, A.P. Syndecan Captures, Protects, and Transmits HIV to T Lymphocytes. Immun. 2003, 18, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Bugatti, A.; Paiardi, G.; Urbinati, C.E.; Chiodelli, P.; Orro, A.; Uggeri, M.; Milanesi, L.; Caruso, A.; Caccuri, F.; D’Ursi, P.; et al. Heparin and heparan sulfate proteoglycans promote HIV-1 p17 matrix protein oligomerization: Computational, biochemical and biological implications. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Hamimi, C.; David, A.; Versmisse, P.; Weiss, L.; Bruel, T.; Zucman, D.; Appay, V.; Moris, A.; Ungeheuer, M.-N.; Lascoux-Combe, C.; et al. Dendritic Cells from HIV Controllers Have Low Susceptibility to HIV-1 Infection In Vitro but High Capacity to Capture HIV-1 Particles. PLoS ONE 2016, 11, e0160251. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Greenwell-Wild, T.; Lei, K.; Jin, W.; Swisher, J.; Hardegen, N.; Wild, C.T.; Wahl, S.M. Secretory Leukocyte Protease Inhibitor Binds to Annexin II, a Cofactor for Macrophage HIV-1 Infection. J. Exp. Med. 2004, 200, 1337–1346. [Google Scholar] [CrossRef] [Green Version]
- Bhakuni, T.; Ali, M.F.; Ahmad, I.; Bano, S.; Ansari, S.; Jairajpuri, M.A. Role of heparin and non heparin binding serpins in coagulation and angiogenesis: A complex interplay. Arch. Biochem. Biophys. 2016, 604, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Rezaie, A.R.; Giri, H. Anticoagulant and signaling functions of antithrombin. J. Thromb. Haemost. 2020, 18, 3142–3153. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biswas, S.; Chen, E.; Gao, Y.; Lee, S.; Hewlett, I.; Devadas, K. Modulation of HIV Replication in Monocyte-Derived Macrophages (MDM) by Host Antiviral Factors Secretory Leukocyte Protease Inhibitor and Serpin Family C Member 1 Induced by Steroid Hormones. Viruses 2022, 14, 95. https://doi.org/10.3390/v14010095
Biswas S, Chen E, Gao Y, Lee S, Hewlett I, Devadas K. Modulation of HIV Replication in Monocyte-Derived Macrophages (MDM) by Host Antiviral Factors Secretory Leukocyte Protease Inhibitor and Serpin Family C Member 1 Induced by Steroid Hormones. Viruses. 2022; 14(1):95. https://doi.org/10.3390/v14010095
Chicago/Turabian StyleBiswas, Santanu, Emily Chen, Yamei Gao, Sherwin Lee, Indira Hewlett, and Krishnakumar Devadas. 2022. "Modulation of HIV Replication in Monocyte-Derived Macrophages (MDM) by Host Antiviral Factors Secretory Leukocyte Protease Inhibitor and Serpin Family C Member 1 Induced by Steroid Hormones" Viruses 14, no. 1: 95. https://doi.org/10.3390/v14010095
APA StyleBiswas, S., Chen, E., Gao, Y., Lee, S., Hewlett, I., & Devadas, K. (2022). Modulation of HIV Replication in Monocyte-Derived Macrophages (MDM) by Host Antiviral Factors Secretory Leukocyte Protease Inhibitor and Serpin Family C Member 1 Induced by Steroid Hormones. Viruses, 14(1), 95. https://doi.org/10.3390/v14010095