
A Transcriptomics-Based Bioinformatics Approach for Identification and In Vitro Screening of FDA-Approved Drugs for Repurposing against Dengue Virus-2

 , , , and
, , , and
Abstract
:1. Introduction
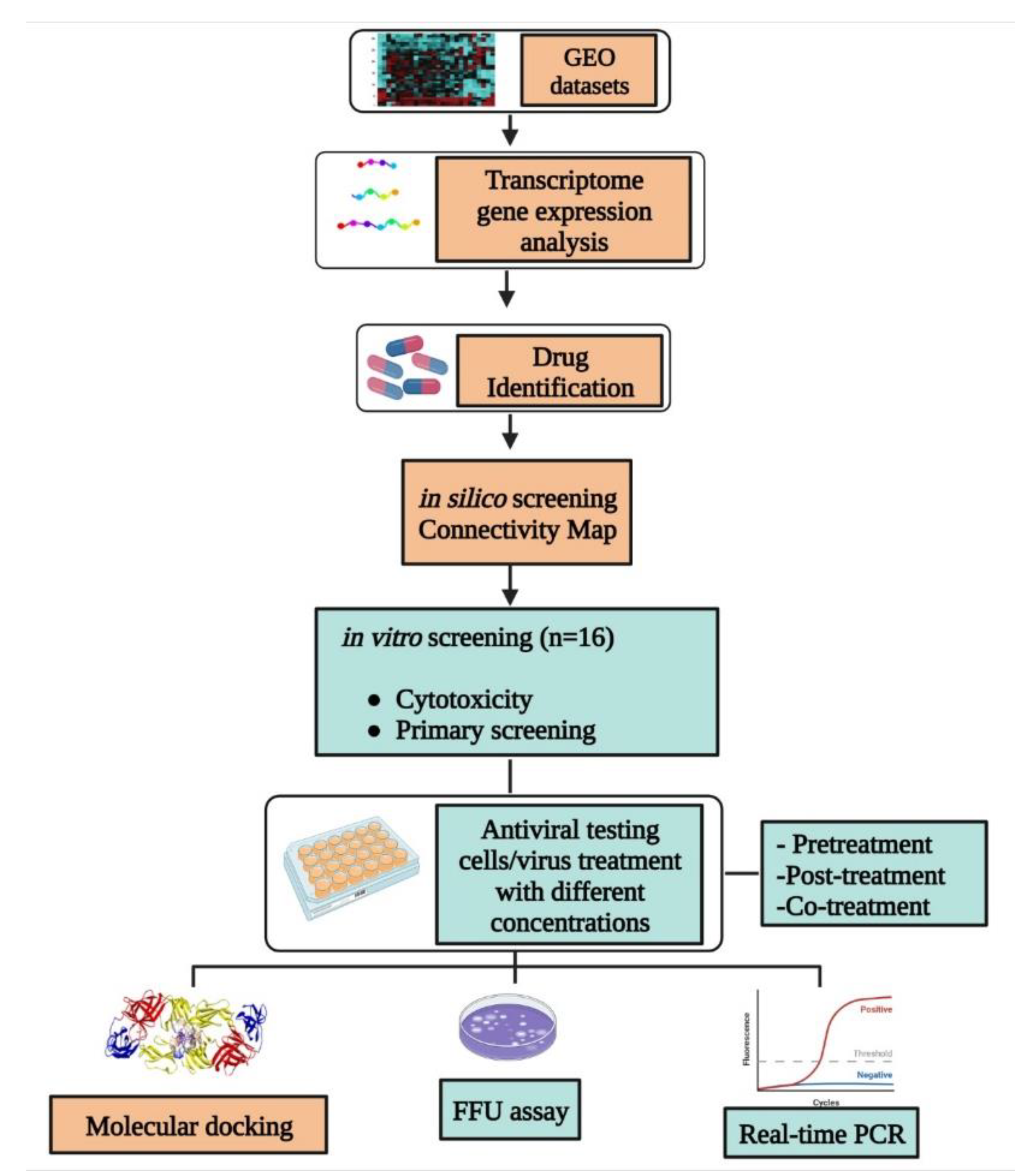
2. Materials and Methods
2.1. Transcriptome Data Collection and Analysis
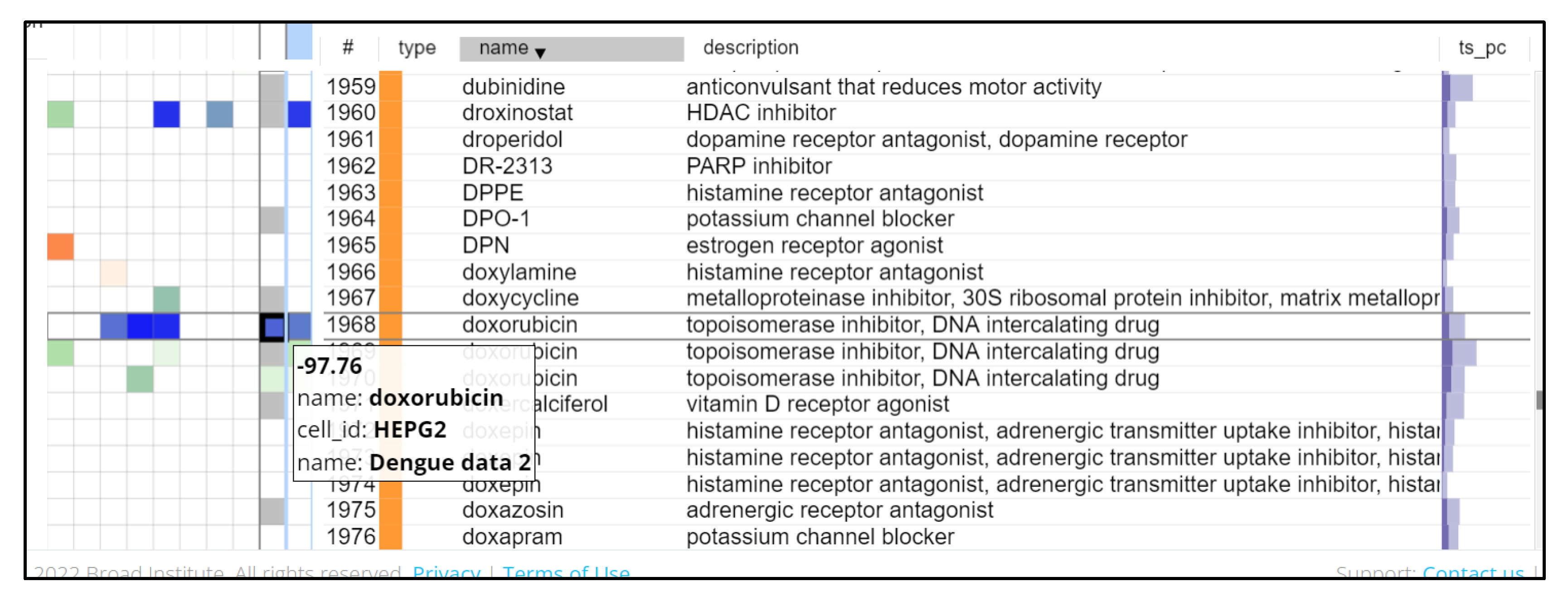
2.2. Identification of Drug Candidates Using CMap
2.3. Cells and Virus
2.4. Stock Preparation of Compounds
2.5. Cytotoxicity Screening of Compounds
2.6. Antiviral Assay
2.7. Focus Forming Unit Assay and Real-Time RT-PCR
2.8. Statistical Analysis
2.9. Molecular Docking Studies with DENV Protein Targets
3. Results
3.1. Transcriptomic Analysis
3.2. CMap Analysis
3.3. Viral Targeting Drugs Identified for Repurposing against DENV
3.4. Cytotoxicity Evaluation of Compounds
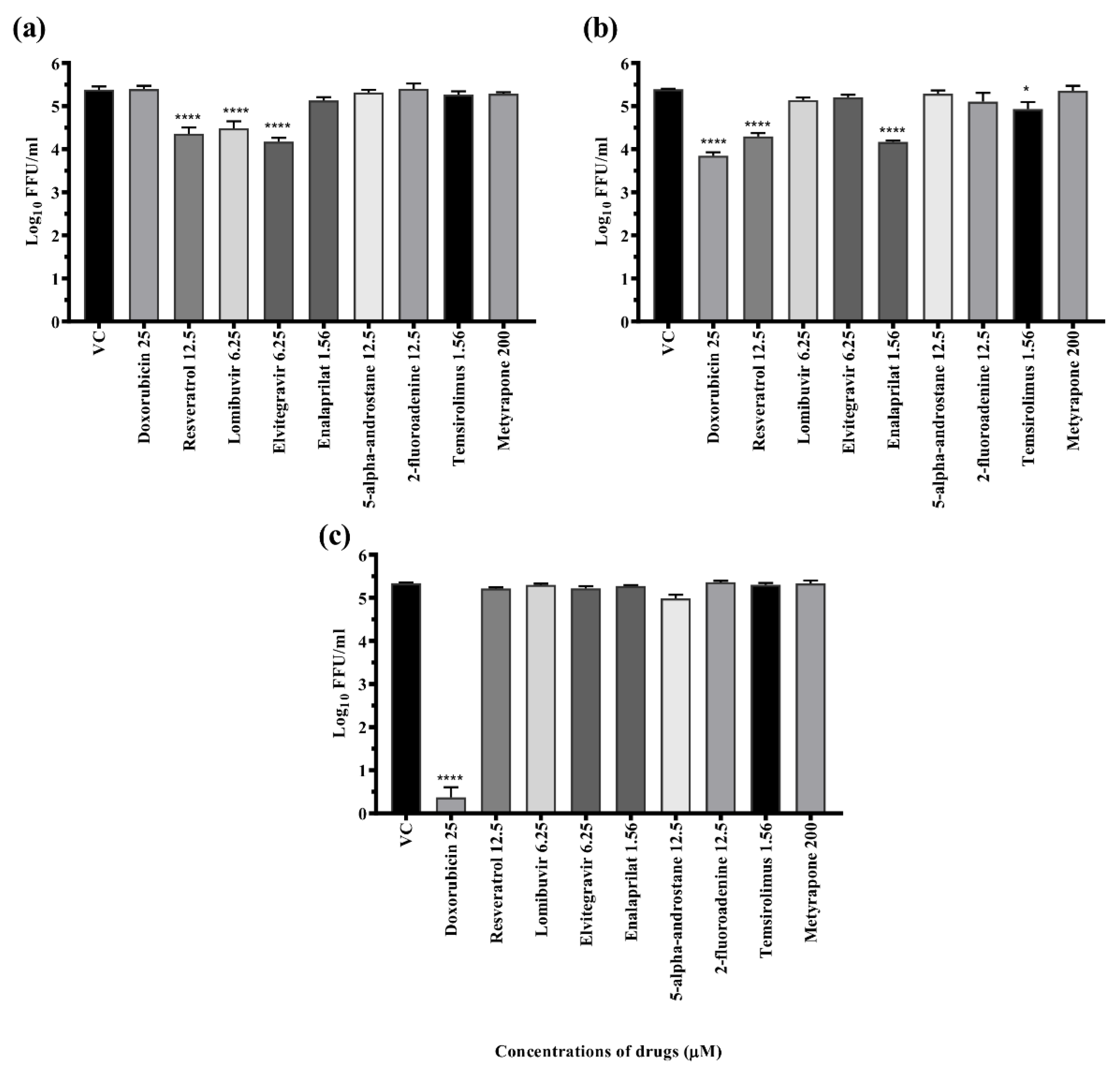
3.5. Primary Antiviral Screening of Compounds against DENV-2
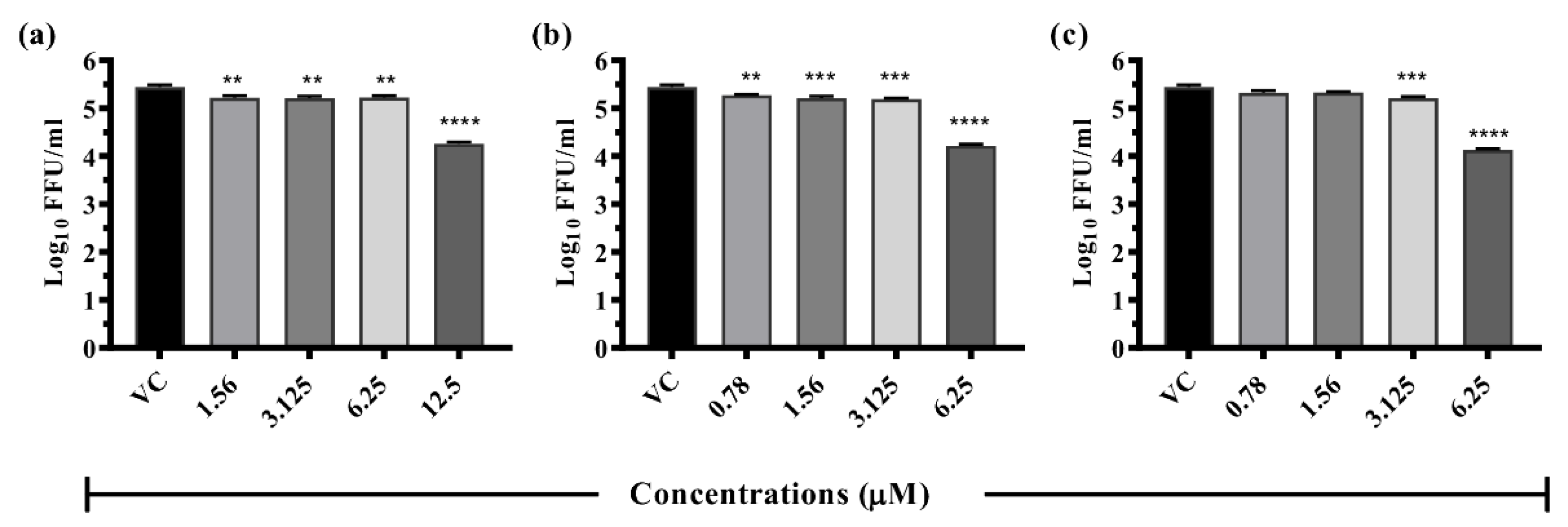
3.6. Dose-Dependent Antiviral Effect of Repurposed Drugs against DENV
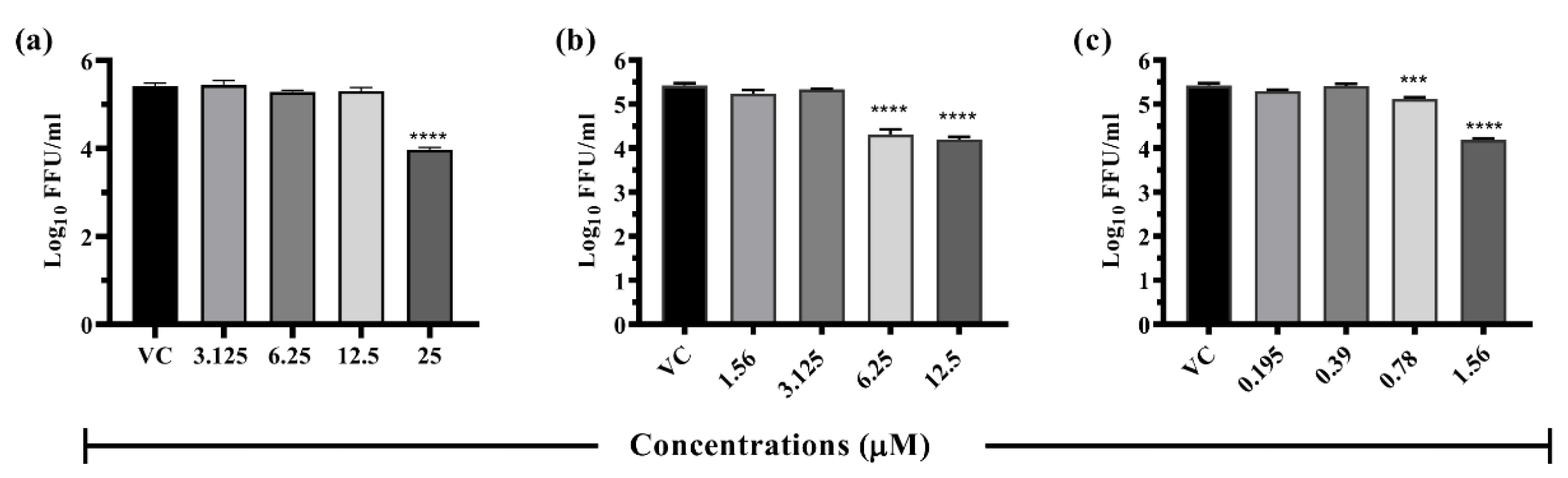
3.6.1. Drugs Exerting Prophylactic Effects
3.6.2. Drugs Exerting Therapeutic Effects
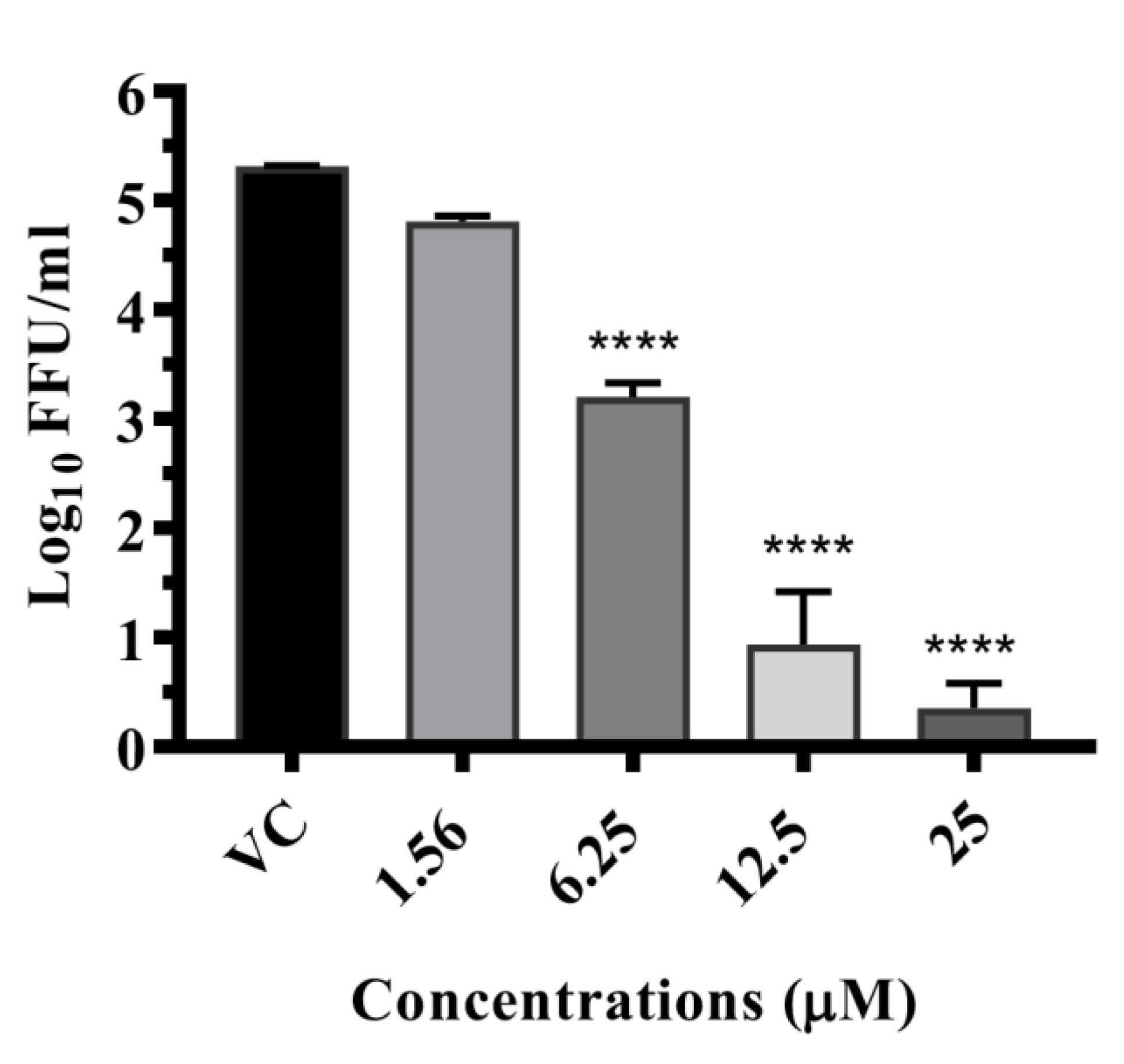
3.6.3. Drugs Exerting Virucidal Effects
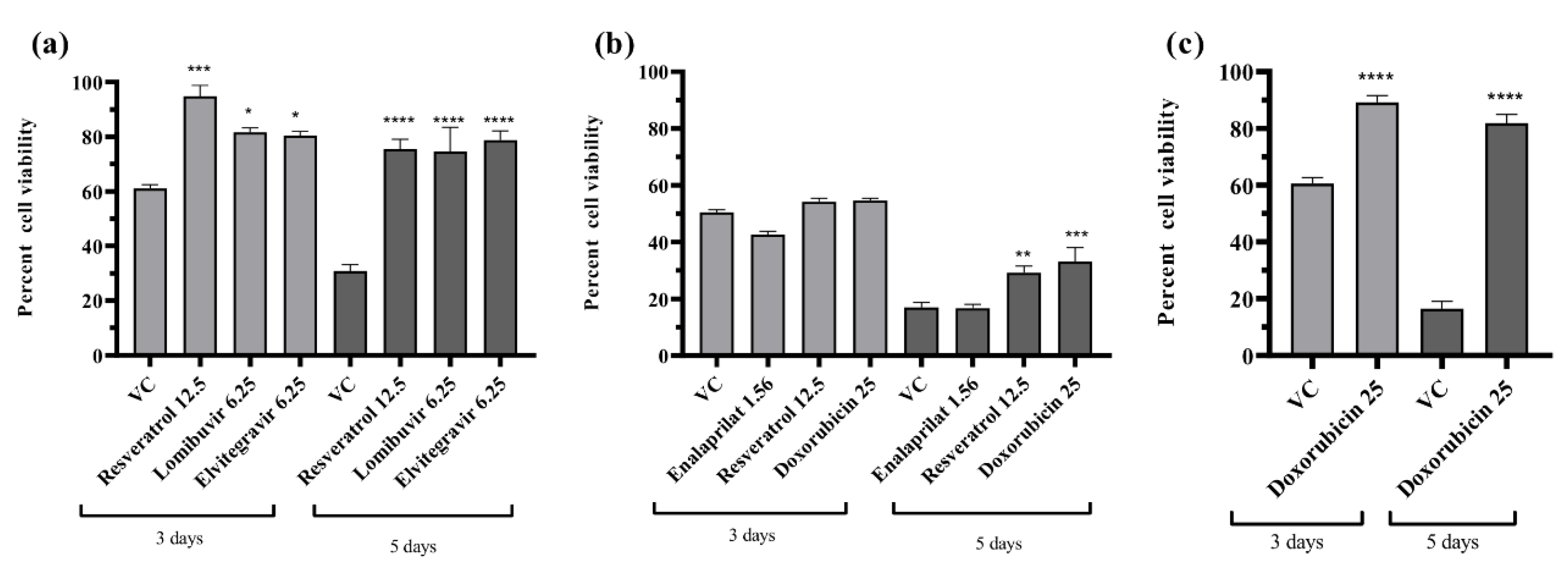
3.7. Effect of Different Drugs on the Percent Cell Viability of the Infected Cells
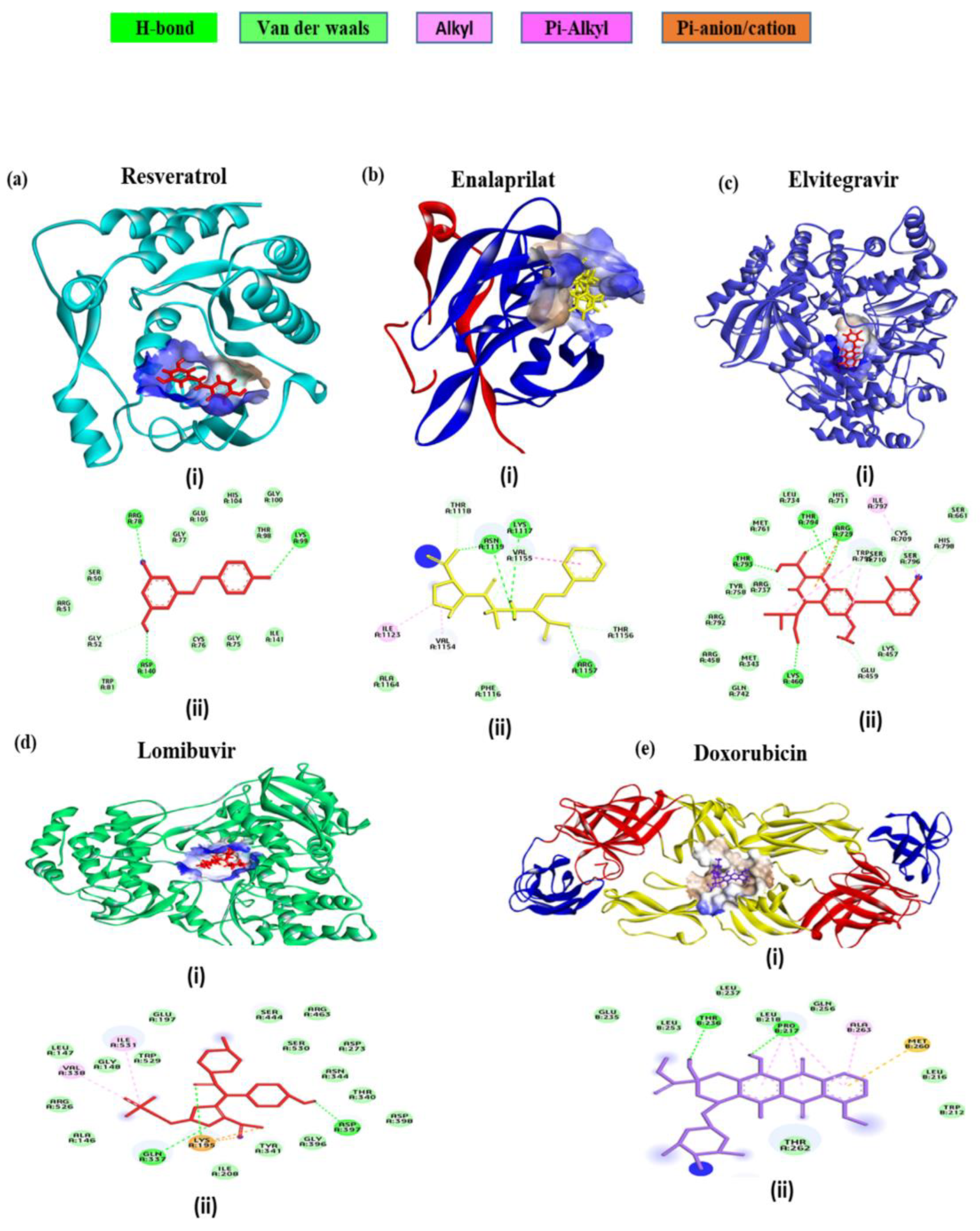
3.8. In Silico Interaction Studies of the Select Drugs with DENV Protein Targets
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gubler, D.J. Dengue and Dengue Hemorrhagic Fever. Clin. Microbiol. Rev. 1998, 11, 480–496. [Google Scholar] [CrossRef] [PubMed]
- Dengue and Severe Dengue. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 17 January 2022).
- Murugesan, A.; Manoharan, M. Dengue Virus. Emerg. Reemerging Viral Pathog. 2020, 1, 281–359. [Google Scholar] [CrossRef]
- De Almeida, R.R.; Paim, B.; de Oliveira, S.A.; Souza, A.S.; Gomes, A.C.P.; Escuissato, D.L.; Zanetti, G.; Marchiori, E. Dengue Hemorrhagic Fever: A State-of-the-Art Review Focused in Pulmonary Involvement. Lung 2017, 195, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Botta, L.; Rivara, M.; Zuliani, V.; Radi, M. Drug Repurposing Approaches to Fight Dengue Virus Infection and Related Diseases. Front. Biosci. Landmark Ed. 2018, 23, 997–1019. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Khairnar, S.J.; Jadhav, A.G. Drug Repurposing (DR): An Emerging Approach in Drug Discovery; IntechOpen: London, UK, 2020; ISBN 978-1-83968-521-7. [Google Scholar]
- Sultana, J.; Crisafulli, S.; Gabbay, F.; Lynn, E.; Shakir, S.; Trifirò, G. Challenges for Drug Repurposing in the COVID-19 Pandemic Era. Front. Pharmacol. 2020, 11, 588654. [Google Scholar] [CrossRef]
- Sleigh, S.H.; Barton, C.L. Repurposing Strategies for Therapeutics. Pharm. Med. 2010, 24, 151–159. [Google Scholar] [CrossRef]
- Low, Z.Y.; Farouk, I.A.; Lal, S.K. Drug Repositioning: New Approaches and Future Prospects for Life-Debilitating Diseases and the COVID-19 Pandemic Outbreak. Viruses 2020, 12, 1058. [Google Scholar] [CrossRef]
- Nascimento, E.J.M.; Braga-Neto, U.; Calzavara-Silva, C.E.; Gomes, A.L.V.; Abath, F.G.C.; Brito, C.A.A.; Cordeiro, M.T.; Silva, A.M.; Magalhães, C.; Andrade, R.; et al. Gene Expression Profiling during Early Acute Febrile Stage of Dengue Infection Can Predict the Disease Outcome. PLoS ONE 2009, 4, e7892. [Google Scholar] [CrossRef]
- Soares-Schanoski, A.; Baptista Cruz, N.; de Castro-Jorge, L.A.; de Carvalho, R.V.H.; Santos, C.A.D.; da Rós, N.; Oliveira, Ú.; Costa, D.D.; Santos, C.L.S.D.; Cunha, M.D.P.; et al. Systems Analysis of Subjects Acutely Infected with the Chikungunya Virus. PLoS Pathog. 2019, 15, e1007880. [Google Scholar] [CrossRef]
- Chandele, A.; Sewatanon, J.; Gunisetty, S.; Singla, M.; Onlamoon, N.; Akondy, R.S.; Kissick, H.T.; Nayak, K.; Reddy, E.S.; Kalam, H.; et al. Characterization of Human CD8 T Cell Responses in Dengue Virus-Infected Patients from India. J. Virol. 2016, 90, 11259–11278. [Google Scholar] [CrossRef] [Green Version]
- Lamb, J.; Crawford, E.D.; Peck, D.; Modell, J.W.; Blat, I.C.; Wrobel, M.J.; Lerner, J.; Brunet, J.-P.; Subramanian, A.; Ross, K.N.; et al. The Connectivity Map: Using Gene-Expression Signatures to Connect Small Molecules, Genes, and Disease. Science 2006, 313, 1929–1935. [Google Scholar] [CrossRef] [PubMed]
- Lamb, J. The Connectivity Map: A New Tool for Biomedical Research. Nat. Rev. Cancer 2007, 7, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Bufalo, D.D.; Ciuffreda, L.; Trisciuoglio, D.; Desideri, M.; Cognetti, F.; Zupi, G.; Milella, M. Antiangiogenic Potential of the Mammalian Target of Rapamycin Inhibitor Temsirolimus. Cancer Res. 2006, 66, 5549–5554. [Google Scholar] [CrossRef] [PubMed]
- Bellmunt, J.; Szczylik, C.; Feingold, J.; Strahs, A.; Berkenblit, A. Temsirolimus Safety Profile and Management of Toxic Effects in Patients with Advanced Renal Cell Carcinoma and Poor Prognostic Features. Ann. Oncol. 2008, 19, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Mullen, P.J.; Garcia, G.; Purkayastha, A.; Matulionis, N.; Schmid, E.W.; Momcilovic, M.; Sen, C.; Langerman, J.; Ramaiah, A.; Shackelford, D.B.; et al. SARS-CoV-2 Infection Rewires Host Cell Metabolism and Is Potentially Susceptible to MTORC1 Inhibition. Nat. Commun. 2021, 12, 1876. [Google Scholar] [CrossRef]
- Saso, W.; Tsukuda, S.; Ohashi, H.; Fukano, K.; Morishita, R.; Matsunaga, S.; Ohki, M.; Ryo, A.; Park, S.-Y.; Suzuki, R.; et al. A New Strategy to Identify Hepatitis B Virus Entry Inhibitors by AlphaScreen Technology Targeting the Envelope-Receptor Interaction. Biochem. Biophys. Res. Commun. 2018, 501, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Tacar, O.; Sriamornsak, P.; Dass, C.R. Doxorubicin: An Update on Anticancer Molecular Action, Toxicity and Novel Drug Delivery Systems. J. Pharm. Pharmacol. 2012, 65, 157–170. [Google Scholar] [CrossRef]
- Coelmont, L.; Kaptein, S.; Paeshuyse, J.; Vliegen, I.; Dumont, J.-M.; Vuagniaux, G.; Neyts, J. Debio 025, a Cyclophilin Binding Molecule, Is Highly Efficient in Clearing Hepatitis C Virus (HCV) Replicon-Containing Cells when Used Alone or in Combination with Specifically Targeted Antiviral Therapy for HCV (STAT-C) Inhibitors. Antimicrob. Agents Chemother. 2009, 53, 967–976. [Google Scholar] [CrossRef]
- De Palma, A.M.; Heggermont, W.; Leyssen, P.; Pürstinger, G.; Wimmer, E.; De Clercq, E.; Rao, A.; Monforte, A.-M.; Chimirri, A.; Neyts, J. Anti-Enterovirus Activity and Structure-Activity Relationship of a Series of 2,6-Dihalophenyl-Substituted 1H,3H-Thiazolo[3,4-a]Benzimidazoles. Biochem. Biophys. Res. Commun. 2007, 353, 628–632. [Google Scholar] [CrossRef]
- Johansson, S.; Goldenberg, D.M.; Griffiths, G.L.; Wahren, B.; Hinkula, J. Elimination of HIV-1 Infection by Treatment with a Doxorubicin-Conjugated Anti-Envelope Antibody. AIDS 2006, 20, 1911–1915. [Google Scholar] [CrossRef]
- Al-Motawa, M.S.; Abbas, H.; Wijten, P.; de la Fuente, A.; Xue, M.; Rabbani, N.; Thornalley, P.J. Vulnerabilities of the SARS-CoV-2 Virus to Proteotoxicity—Opportunity for Repurposed Chemotherapy of COVID-19 Infection. Front. Pharmacol. 2020, 11, 585408. [Google Scholar] [CrossRef] [PubMed]
- Kaptein, S.J.F.; Burghgraeve, T.D.; Froeyen, M.; Pastorino, B.; Alen, M.M.F.; Mondotte, J.A.; Herdewijn, P.; Jacobs, M.; de Lamballerie, X.; Schols, D.; et al. A Derivate of the Antibiotic Doxorubicin Is a Selective Inhibitor of Dengue and Yellow Fever Virus Replication In Vitro. Antimicrob. Agents Chemother. 2010, 54, 5269–5280. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.R.; Peterson, B.L.; Appelbaum, F.R.; Kolitz, J.; Elias, L.; Shepherd, L.; Hines, J.; Threatte, G.A.; Larson, R.A.; Cheson, B.D.; et al. Fludarabine Compared with Chlorambucil as Primary Therapy for Chronic Lymphocytic Leukemia. N. Engl. J. Med. 2000, 343, 1750–1757. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.A.; Mahajan, S.; Ritz, J. Fludarabine-Induced Immunosuppression Is Associated with Inhibition of STAT1 Signaling. Nat. Med. 1999, 5, 444–447. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Liu, Y.-F.; Wu, C.-F.; Xu, F.; Shen, Z.-X.; Zhu, Y.-M.; Li, J.-M.; Tang, W.; Zhao, W.-L.; Wu, W.; et al. Long-Term Efficacy and Safety of All-Trans Retinoic Acid/Arsenic Trioxide-Based Therapy in Newly Diagnosed Acute Promyelocytic Leukemia. Proc. Natl. Acad. Sci. USA 2009, 106, 3342–3347. [Google Scholar] [CrossRef]
- Engels, F.K.; ten Tije, A.J.; Baker, S.D.; Lee, C.K.K.; Loos, W.J.; Vulto, A.G.; Verweij, J.; Sparreboom, A. Effect of Cytochrome P450 3A4 Inhibition on the Pharmacokinetics of Docetaxel. Clin. Pharmacol. Ther. 2004, 75, 448–454. [Google Scholar] [CrossRef]
- Engels, F.K.; Sparreboom, A.; Mathot, R.A.A.; Verweij, J. Potential for Improvement of Docetaxel-Based Chemotherapy: A Pharmacological Review. Br. J. Cancer 2005, 93, 173–177. [Google Scholar] [CrossRef]
- Shin, H.J.; Kim, D.N.; Lee, S.K. Association between Epstein-Barr Virus Infection and Chemoresistance to Docetaxel in Gastric Carcinoma. Mol. Cells 2011, 32, 173–179. [Google Scholar] [CrossRef]
- Fei, X.F.; Wang, B.X.; Li, T.J.; Tashiro, S.; Minami, M.; Xing, D.J.; Ikejima, T. Evodiamine, a Constituent of Evodiae Fructus, Induces Anti-Proliferating Effects in Tumor Cells. Cancer Sci. 2003, 94, 92–98. [Google Scholar] [CrossRef]
- Jiang, J.; Hu, C. Evodiamine: A Novel Anti-Cancer Alkaloid from Evodia Rutaecarpa. Molecules 2009, 14, 1852–1859. [Google Scholar] [CrossRef]
- Choi, Y.H.; Shin, E.M.; Kim, Y.S.; Cai, X.F.; Lee, J.J.; Kim, H.P. Anti-Inflammatory Principles from the Fruits of Evodia Rutaecarpa and Their Cellular Action Mechanisms. Arch. Pharm. Res. 2006, 29, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.-P.; Li, W.-Z.; Zhao, X.-F.; Wang, G.-F.; Yang, J.-C.; Zhang, L.; Chen, X.-X.; Xu, Y.-X.; Li, K.-S. A Drug Screening Method Based on the Autophagy Pathway and Studies of the Mechanism of Evodiamine against Influenza A Virus. PLoS ONE 2012, 7, e42706. [Google Scholar] [CrossRef] [PubMed]
- Majnooni, M.B.; Fakhri, S.; Bahrami, G.; Naseri, M.; Farzaei, M.H.; Echeverría, J. Alkaloids as Potential Phytochemicals against SARS-CoV-2: Approaches to the Associated Pivotal Mechanisms. Evid. Based Complement. Altern. Med. 2021, 2021, e6632623. [Google Scholar] [CrossRef] [PubMed]
- Vidoni, C.; Fuzimoto, A.; Ferraresi, A.; Isidoro, C. Targeting Autophagy with Natural Products to Prevent SARS-CoV-2 Infection. J. Tradit. Complement. Med. 2021. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, S.; Li, H. Evodiamine Alleviates Severe Pneumonia Induced by Methicillin-Susceptible Staphylococcus Aureus Following Cytomegalovirus Reactivation through Suppressing NF-ΚB and MAPKs. Int. J. Mol. Med. 2018, 42, 3247–3255. [Google Scholar] [CrossRef]
- Cho, K.-J.; Park, J.-H.; Hancock, J.F. Staurosporine. Commun. Integr. Biol. 2013, 6, e24746. [Google Scholar] [CrossRef]
- Malsy, M.; Bitzinger, D.; Graf, B.; Bundscherer, A. Staurosporine Induces Apoptosis in Pancreatic Carcinoma Cells PaTu 8988t and Panc-1 via the Intrinsic Signaling Pathway. Eur. J. Med. Res. 2019, 24, 5. [Google Scholar] [CrossRef]
- Omura, S.; Sasaki, Y.; Iwai, Y.; Takeshima, H. Staurosporine, a Potentially Important Gift from a Microorganism. J. Antibiot. 1995, 48, 535–548. [Google Scholar] [CrossRef]
- Szkudelska, K.; Szkudelski, T. Resveratrol, Obesity and Diabetes. Eur. J. Pharmacol. 2010, 635, 1–8. [Google Scholar] [CrossRef]
- Das, S.; Das, D.K. Anti-Inflammatory Responses of Resveratrol. Inflamm. Allergy Drug Targets Discontin. 2022, 6, 168–173. [Google Scholar] [CrossRef]
- Elmali, N.; Baysal, O.; Harma, A.; Esenkaya, I.; Mizrak, B. Effects of Resveratrol in Inflammatory Arthritis. Inflammation 2007, 30, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The Therapeutic Potential of Resveratrol: A Review of Clinical Trials. Npj Precis. Oncol. 2017, 1, 1–9. [Google Scholar] [CrossRef]
- Anekonda, T.S. Resveratrol—A Boon for Treating Alzheimer’s Disease? Brain Res. Rev. 2006, 52, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Filip, V.; Plocková, M.; Šmidrkal, J.; Špičková, Z.; Melzoch, K.; Schmidt, Š. Resveratrol and Its Antioxidant and Antimicrobial Effectiveness. Food Chem. 2003, 83, 585–593. [Google Scholar] [CrossRef]
- Campagna, M.; Rivas, C. Antiviral Activity of Resveratrol. Biochem. Soc. Trans. 2010, 38, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Bradamante, S.; Barenghi, L.; Villa, A. Cardiovascular Protective Effects of Resveratrol. Cardiovasc. Drug Rev. 2004, 22, 169–188. [Google Scholar] [CrossRef]
- Daniel, E.; Aylwin, S.; Mustafa, O.; Ball, S.; Munir, A.; Boelaert, K.; Chortis, V.; Cuthbertson, D.J.; Daousi, C.; Rajeev, S.P.; et al. Effectiveness of Metyrapone in Treating Cushing’s Syndrome: A Retrospective Multicenter Study in 195 Patients. J. Clin. Endocrinol. Metab. 2015, 100, 4146–4154. [Google Scholar] [CrossRef]
- Jahn, H.; Schick, M.; Kiefer, F.; Kellner, M.; Yassouridis, A.; Wiedemann, K. Metyrapone as Additive Treatment in Major Depression: A Double-Blind and Placebo-Controlled Trial. Arch. Gen. Psychiatry 2004, 61, 1235–1244. [Google Scholar] [CrossRef]
- Filippakopoulos, P.; Qi, J.; Picaud, S.; Shen, Y.; Smith, W.B.; Fedorov, O.; Morse, E.M.; Keates, T.; Hickman, T.T.; Felletar, I.; et al. Selective Inhibition of BET Bromodomains. Nature 2010, 468, 1067–1073. [Google Scholar] [CrossRef]
- Gibbons, H.R.; Mi, D.J.; Farley, V.M.; Esmond, T.; Kaood, M.B.; Aune, T.M. Bromodomain Inhibitor JQ1 Reversibly Blocks IFN-γ Production. Sci. Rep. 2019, 9, 10280. [Google Scholar] [CrossRef] [Green Version]
- Delmore, J.E.; Issa, G.C.; Lemieux, M.E.; Rahl, P.B.; Shi, J.; Jacobs, H.M.; Kastritis, E.; Gilpatrick, T.; Paranal, R.M.; Qi, J.; et al. BET Bromodomain Inhibition as a Therapeutic Strategy to Target C-Myc. Cell 2011, 146, 904–917. [Google Scholar] [CrossRef] [PubMed]
- Baratta, M.G.; Schinzel, A.C.; Zwang, Y.; Bandopadhayay, P.; Bowman-Colin, C.; Kutt, J.; Curtis, J.; Piao, H.; Wong, L.C.; Kung, A.L.; et al. An In-Tumor Genetic Screen Reveals That the BET Bromodomain Protein, BRD4, Is a Potential Therapeutic Target in Ovarian Carcinoma. Proc. Natl. Acad. Sci. USA 2015, 112, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Zhang, D.; Tian, Y.; Xie, Z.; Zou, M.-H. BRD4 Inhibition by JQ1 Prevents High-Fat Diet-Induced Diabetic Cardiomyopathy by Activating PINK1/Parkin-Mediated Mitophagy In Vivo. J. Mol. Cell. Cardiol. 2020, 149, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Stratton, M.S.; Bagchi, R.A.; Felisbino, M.B.; Hirsch, R.A.; Smith, H.E.; Riching, A.S.; Enyart, B.Y.; Koch, K.A.; Cavasin, M.A.; Alexanian, M.; et al. Dynamic Chromatin Targeting of BRD4 Stimulates Cardiac Fibroblast Activation. Circ. Res. 2019, 125, 662–677. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Zhang, L.; Tang, Y.; Tu, Q.; Zheng, L.; Yu, L.; Murray, D.; Cheng, J.; Kim, S.H.; Zhou, X.; et al. BET Inhibitor JQ1 Blocks Inflammation and Bone Destruction. J. Dent. Res. 2014, 93, 657–662. [Google Scholar] [CrossRef]
- Leoni, F.; Fossati, G.; Lewis, E.C.; Lee, J.-K.; Porro, G.; Pagani, P.; Modena, D.; Moras, M.L.; Pozzi, P.; Reznikov, L.L.; et al. The Histone Deacetylase Inhibitor ITF2357 Reduces Production of Pro-Inflammatory Cytokines In Vitro and Systemic Inflammation In Vivo. Mol. Med. 2005, 11, 1–15. [Google Scholar] [CrossRef]
- Milan, M.; Pace, V.; Maiullari, F.; Chirivì, M.; Baci, D.; Maiullari, S.; Madaro, L.; Maccari, S.; Stati, T.; Marano, G.; et al. Givinostat Reduces Adverse Cardiac Remodeling through Regulating Fibroblasts Activation. Cell Death Dis. 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Tambaro, F.P.; Dell’Aversana, C.; Carafa, V.; Nebbioso, A.; Radic, B.; Ferrara, F.; Altucci, L. Histone Deacetylase Inhibitors: Clinical Implications for Hematological Malignancies. Clin. Epigenet. 2010, 1, 25–44. [Google Scholar] [CrossRef]
- Jaskulska, A.; Janecka, A.E.; Gach-Janczak, K. Thapsigargin—From Traditional Medicine to Anticancer Drug. Int. J. Mol. Sci. 2020, 22, 4. [Google Scholar] [CrossRef]
- Sehgal, P.; Szalai, P.; Olesen, C.; Praetorius, H.A.; Nissen, P.; Christensen, S.B.; Engedal, N.; Møller, J.V. Inhibition of the Sarco/Endoplasmic Reticulum (ER) Ca2+-ATPase by Thapsigargin Analogs Induces Cell Death via ER Ca2+ Depletion and the Unfolded Protein Response. J. Biol. Chem. 2017, 292, 19656–19673. [Google Scholar] [CrossRef] [Green Version]
- Al-Beltagi, S.; Preda, C.A.; Goulding, L.V.; James, J.; Pu, J.; Skinner, P.; Jiang, Z.; Wang, B.L.; Yang, J.; Banyard, A.C.; et al. Thapsigargin Is a Broad-Spectrum Inhibitor of Major Human Respiratory Viruses: Coronavirus, Respiratory Syncytial Virus and Influenza A Virus. Viruses 2021, 13, 234. [Google Scholar] [CrossRef]
- Goulding, L.V.; Yang, J.; Jiang, Z.; Zhang, H.; Lea, D.; Emes, R.D.; Dottorini, T.; Pu, J.; Liu, J.; Chang, K.-C. Thapsigargin at Non-Cytotoxic Levels Induces a Potent Host Antiviral Response That Blocks Influenza A Virus Replication. Viruses 2020, 12, 1093. [Google Scholar] [CrossRef]
- Ayaz, S.I.; Sharkey, C.M.; Kwiatkowski, G.M.; Wilson, S.S.; John, R.S.; Tolomello, R.; Mahajan, A.; Millis, S.; Levy, P.D. Intravenous Enalaprilat for Treatment of Acute Hypertensive Heart Failure in the Emergency Department. Int. J. Emerg. Med. 2016, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.K. Critical Appraisal of Elvitegravir in the Treatment of HIV-1/AIDS. HIVAIDS 2014, 6, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Schafer, J.J.; Squires, K.E. Integrase Inhibitors: A Novel Class of Antiretroviral Agents. Ann. Pharmacother. 2010, 44, 145–156. [Google Scholar] [CrossRef]
- Li, P.; Dorsch, W.; Lauffer, D.J.; Bilimoria, D.; Chauret, N.; Court, J.J.; Das, S.K.; Denis, F.; Mani, N.; Nanthakumar, S.; et al. Discovery of Novel Allosteric HCV NS5B Inhibitors. 2. Lactam-Containing Thiophene Carboxylates. ACS Med. Chem. Lett. 2017, 8, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Abdurakhmanov, E.; Øie Solbak, S.; Danielson, U.H. Biophysical Mode-of-Action and Selectivity Analysis of Allosteric Inhibitors of Hepatitis C Virus (HCV) Polymerase. Viruses 2017, 9, 151. [Google Scholar] [CrossRef]
- Mohamed, K.; Yazdanpanah, N.; Saghazadeh, A.; Rezaei, N. Computational Drug Discovery and Repurposing for the Treatment of COVID-19: A Systematic Review. Bioorg. Chem. 2021, 106, 104490. [Google Scholar] [CrossRef]
- Parashar, D.; Paingankar, M.S.; Kumar, S.; Gokhale, M.D.; Sudeep, A.B.; Shinde, S.B.; Arankalle, V.A. Administration of E2 and NS1 SiRNAs Inhibit Chikungunya Virus Replication in Vitro and Protects Mice Infected with the Virus. PLoS Negl. Trop. Dis. 2013, 7, e2405. [Google Scholar] [CrossRef]
- Patil, P.; Agrawal, M.; Almelkar, S.; Jeengar, M.K.; More, A.; Alagarasu, K.; Kumar, N.V.; Mainkar, P.S.; Parashar, D.; Cherian, S. In Vitro and In Vivo Studies Reveal α-Mangostin, a Xanthonoid from Garcinia Mangostana, as a Promising Natural Antiviral Compound against Chikungunya Virus. Virol. J. 2021, 18, 47. [Google Scholar] [CrossRef]
- Panda, K.; Alagarasu, K.; Patil, P.; Agrawal, M.; More, A.; Kumar, N.V.; Mainkar, P.S.; Parashar, D.; Cherian, S. In Vitro Antiviral Activity of α-Mangostin against Dengue Virus Serotype-2 (DENV-2). Molecules 2021, 26, 3016. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound Databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 1. Method and Assessment of Docking Accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- Halgren, T.A.; Murphy, R.B.; Friesner, R.A.; Beard, H.S.; Frye, L.L.; Pollard, W.T.; Banks, J.L. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 2. Enrichment Factors in Database Screening. J. Med. Chem. 2004, 47, 1750–1759. [Google Scholar] [CrossRef] [PubMed]
- Modis, Y.; Ogata, S.; Clements, D.; Harrison, S.C. A Ligand-Binding Pocket in the Dengue Virus Envelope Glycoprotein. Proc. Natl. Acad. Sci. USA 2003, 100, 6986–6991. [Google Scholar] [CrossRef]
- Yildiz, M.; Ghosh, S.; Bell, J.A.; Sherman, W.; Hardy, J.A. Allosteric Inhibition of the NS2B-NS3 Protease from Dengue Virus. ACS Chem. Biol. 2013, 8, 2744–2752. [Google Scholar] [CrossRef]
- El Sahili, A.; Soh, T.S.; Schiltz, J.; Gharbi-Ayachi, A.; Seh, C.C.; Shi, P.-Y.; Lim, S.P.; Lescar, J. NS5 from Dengue Virus Serotype 2 Can Adopt a Conformation Analogous to That of Its Zika Virus and Japanese Encephalitis Virus Homologues. J. Virol. 2019, 94, e01294-19. [Google Scholar] [CrossRef]
- Ayala-Nuñez, N.V.; Jarupathirun, P.; Kaptein, S.J.F.; Neyts, J.; Smit, J.M. Antibody-Dependent Enhancement of Dengue Virus Infection Is Inhibited by SA-17, a Doxorubicin Derivative. Antiviral Res. 2013, 100, 238–245. [Google Scholar] [CrossRef]
- Guzman, M.G.; Vazquez, S. The Complexity of Antibody-Dependent Enhancement of Dengue Virus Infection. Viruses 2010, 2, 2649–2662. [Google Scholar] [CrossRef] [Green Version]
- Fried, J.R.; Gibbons, R.V.; Kalayanarooj, S.; Thomas, S.J.; Srikiatkhachorn, A.; Yoon, I.-K.; Jarman, R.G.; Green, S.; Rothman, A.L.; Cummings, D.A.T. Serotype-Specific Differences in the Risk of Dengue Hemorrhagic Fever: An Analysis of Data Collected in Bangkok, Thailand from 1994 to 2006. PLoS Negl. Trop. Dis. 2010, 4, e617. [Google Scholar] [CrossRef] [PubMed]
- Malakar, S.; Sreelatha, L.; Dechtawewat, T.; Noisakran, S.; Yenchitsomanus, P.; Chu, J.J.H.; Limjindaporn, T. Drug Repurposing of Quinine as Antiviral against Dengue Virus Infection. Virus Res. 2018, 255, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Paemanee, A.; Hitakarun, A.; Roytrakul, S.; Smith, D.R. Screening of Melatonin, α-Tocopherol, Folic Acid, Acetyl-l-Carnitine and Resveratrol for Anti-Dengue 2 Virus Activity. BMC Res. Notes 2018, 11, 307. [Google Scholar] [CrossRef] [PubMed]
- Kato, F.; Ishida, Y.; Oishi, S.; Fujii, N.; Watanabe, S.; Vasudevan, S.G.; Tajima, S.; Takasaki, T.; Suzuki, Y.; Ichiyama, K.; et al. Novel Antiviral Activity of Bromocriptine against Dengue Virus Replication. Antiviral Res. 2016, 131, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.-J.; Hanh, V.T.; Nguyen, T.Q.; Jhan, M.-K.; Ho, M.-R.; Lin, C.-F. Repurposing the Antiemetic Metoclopramide as an Antiviral against Dengue Virus Infection in Neuronal Cells. Front. Cell. Infect. Microbiol. 2021, 10, 606743. [Google Scholar] [CrossRef]
- Zainal, N.; Chang, C.-P.; Cheng, Y.-L.; Wu, Y.-W.; Anderson, R.; Wan, S.-W.; Chen, C.-L.; Ho, T.-S.; AbuBakar, S.; Lin, Y.-S. Resveratrol Treatment Reveals a Novel Role for HMGB1 in Regulation of the Type 1 Interferon Response in Dengue Virus Infection. Sci. Rep. 2017, 7, 42998. [Google Scholar] [CrossRef]
- Chatterjee, K.; Zhang, J.; Honbo, N.; Karliner, J.S. Doxorubicin Cardiomyopathy. Cardiology 2010, 115, 155–162. [Google Scholar] [CrossRef]
- Smeets, N.J.L.; Schreuder, M.F.; Dalinghaus, M.; Male, C.; Lagler, F.B.; Walsh, J.; Laer, S.; de Wildt, S.N. Pharmacology of Enalapril in Children: A Review. Drug Discov. Today 2020, 25, 1957–1970. [Google Scholar] [CrossRef]
- Hernández-Fonseca, J.P.; Durán, A.; Valero, N.; Mosquera, J. Losartan and Enalapril Decrease Viral Absorption and Interleukin 1 Beta Production by Macrophages in an Experimental Dengue Virus Infection. Arch. Virol. 2015, 160, 2861–2865. [Google Scholar] [CrossRef]
- Loe, M.W.C.; Lee, R.C.H.; Chu, J.J.H. Antiviral Activity of the FDA-Approved Drug Candesartan Cilexetil against Zika Virus Infection. Antiviral Res. 2019, 172, 104637. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Compound Name | Pharmacological Class | CC50 Value (µM) | Reported Activity |
|---|---|---|---|---|
| 1. | Temsirolimus | Antineoplastic | 12.24 | 1. Antineoplastic [15,16] 2. Antiviral, anti-SARS-CoV-2, and anti-HBV [17,18] |
| 2. | Doxorubicin hydrochloride | Antineoplastic | 116.9 | 1. Antineoplastic [19] 2. Antiviral [20,21,22,23,24] |
| 3. | 2-Fluoroadenine-9-β-D-arabinofuranoside | Antineoplastic | 42.24 | 1. Antineoplastic [25] 2. Immunosuppressant [26] |
| 4. | Retinoic acid p-hydroxyanilide | Antineoplastic | 2.38 | 1. Antineoplastic [27] |
| 5. | Docetaxel | Antineoplastic | 2.26 | 1. Antineoplastic [28,29] 2. Antiviral-EBV [30] |
| 6. | Evodiamine | Antineoplastic | 10.24 | 1. Antineoplastic [31,32] 2. Anti-inflammatory [33] 3. Antiviral [34,35,36], viral, and bacterial pneumonia [37] |
| 7. | Staurosporine from Streptomyces sp. | Antibiotic | 0.08 | 1. Antineoplastic [38,39] 2. Antimicrobial [40] |
| 8. | Resveratrol | Anti-inflammatory | 40.82 | 1. Antilipemic and antidiabetic [41] 2. Anti-inflammatory [42,43] 3. Antineoplastic [44] 4. Alzheimer’s disease pathomechanism modulator [45] 5. Antioxidant and antimicrobial [46] 6. Antiviral [47] 7. Cardioprotective [48] |
| 9. | Metyrapone | Adrenal steroid synthesis inhibitor | 19,554 | 1. 11β-hydroxylase enzyme inhibitor [49] 2. Antidepressant [50] |
| 10. | (+)-JQ1 | Bromodomain inhibitor (Thienotriazolodiazepine) | 0.82 | 1. Bromodomain inhibitor [51] 2. Reduces IFN-γ expression [52] 3. Antineoplastic [53,54] 4. Cardioprotective [55,56] 5. Anti-inflammatory [57] |
| 11. | Givinostat hydrochloride hydrate | Hydroxamate inhibitor | 1.61 | 1. Anti-inflammatory [58] 2. Cardioprotective [59] 3. Antineoplastic/antiangiogenic [60] |
| 12. | Thapsigargin | Calcium channel blocker | 1.38 | 1. Antineoplastic [61] 2. Endoplasmic reticulum Ca 2+ inhibitor [62] 3. Antiviral [63,64] |
| 13. | Enalaprilat | Angiotensin-converting enzyme (ACE) inhibitor | 80.83 | 1. Treatment of hypertension and hypertensive heart failure [65] |
| 14. | 5α-Androstan-3β-ol | Steroid | 23.13 | No reported activity |
| 15. | Elvitegravir | Antiviral | 12.7 | 1. Antiretroviral [66,67] |
| 16. | Lomibuvir | Antiviral | 38.54 | 1. Anti-HCV activity [68,69] 2. SARS-CoV-2 [70] |
| Sr. No. | Compound Name | Chemical Structure | CC50 (µM) | Maximum Concentration (µM) | Log Difference Effectiveness against DENV-2 | EC50 (µM) | Selectivity Index (SI) |
|---|---|---|---|---|---|---|---|
| 1 | Doxorubicin |  | 116.9 | 25 | Post-treatment—1.453 Co-treatment—4.958 | 19.99 6.573 | 5.848 17.785 |
| 12.5 | Co-treatment—4.377 | 6.573 | 17.785 | ||||
| 6.25 | Co-treatment—2.107 | 6.573 | 16.115 | ||||
| 2 | Resveratrol |  | 40.54 | 12.5 | Post-treatment—1.226 Pre-treatment—1.188 | 4.013 7.592 | 10.102 5.340 |
| 6.25 | Post-treatment—1.115 | 4.013 | 10.102 | ||||
| 3 | Enalaprilat |  | 80.83 | 1.56 | Post-treatment—1.239 | 1.079 | 74.911 |
| 4 | Elvitegravir |  | 12.7 | 6.25 | Pre-treatment—1.316 | 4.405 | 2.883 |
| 5 | Lomibuvir |  | 38.54 | 6.25 | Pre-treatment—1.224 | 3.740 | 10.305 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Punekar, M.; Kasabe, B.; Patil, P.; Kakade, M.B.; Parashar, D.; Alagarasu, K.; Cherian, S. A Transcriptomics-Based Bioinformatics Approach for Identification and In Vitro Screening of FDA-Approved Drugs for Repurposing against Dengue Virus-2. Viruses 2022, 14, 2150. https://doi.org/10.3390/v14102150
Punekar M, Kasabe B, Patil P, Kakade MB, Parashar D, Alagarasu K, Cherian S. A Transcriptomics-Based Bioinformatics Approach for Identification and In Vitro Screening of FDA-Approved Drugs for Repurposing against Dengue Virus-2. Viruses. 2022; 14(10):2150. https://doi.org/10.3390/v14102150
Chicago/Turabian StylePunekar, Madhura, Bhagyashri Kasabe, Poonam Patil, Mahadeo B. Kakade, Deepti Parashar, Kalichamy Alagarasu, and Sarah Cherian. 2022. "A Transcriptomics-Based Bioinformatics Approach for Identification and In Vitro Screening of FDA-Approved Drugs for Repurposing against Dengue Virus-2" Viruses 14, no. 10: 2150. https://doi.org/10.3390/v14102150
APA StylePunekar, M., Kasabe, B., Patil, P., Kakade, M. B., Parashar, D., Alagarasu, K., & Cherian, S. (2022). A Transcriptomics-Based Bioinformatics Approach for Identification and In Vitro Screening of FDA-Approved Drugs for Repurposing against Dengue Virus-2. Viruses, 14(10), 2150. https://doi.org/10.3390/v14102150