

Identification of Three Viruses Infecting Mulberry Varieties

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Sequence Analyses
2.3. Phylogenetic Analyses
2.4. Completion of Genomes of Viruses
2.5. Mechanical Inoculation and Virus Detection by RT-PCR
3. Results
3.1. Discovery of Viruses and Sequencing Results
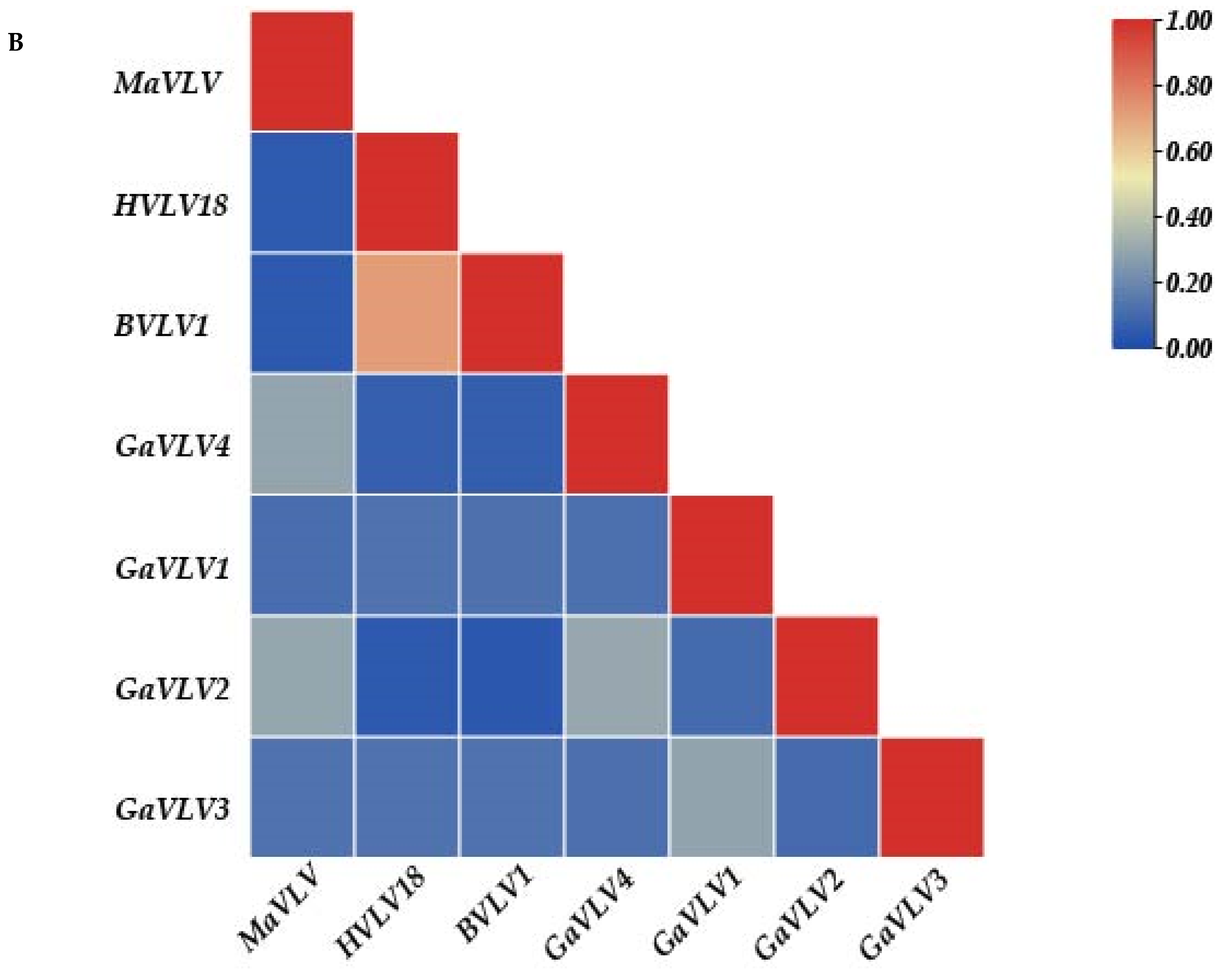
3.2. Phylogenetic Placement of Identified Viruses
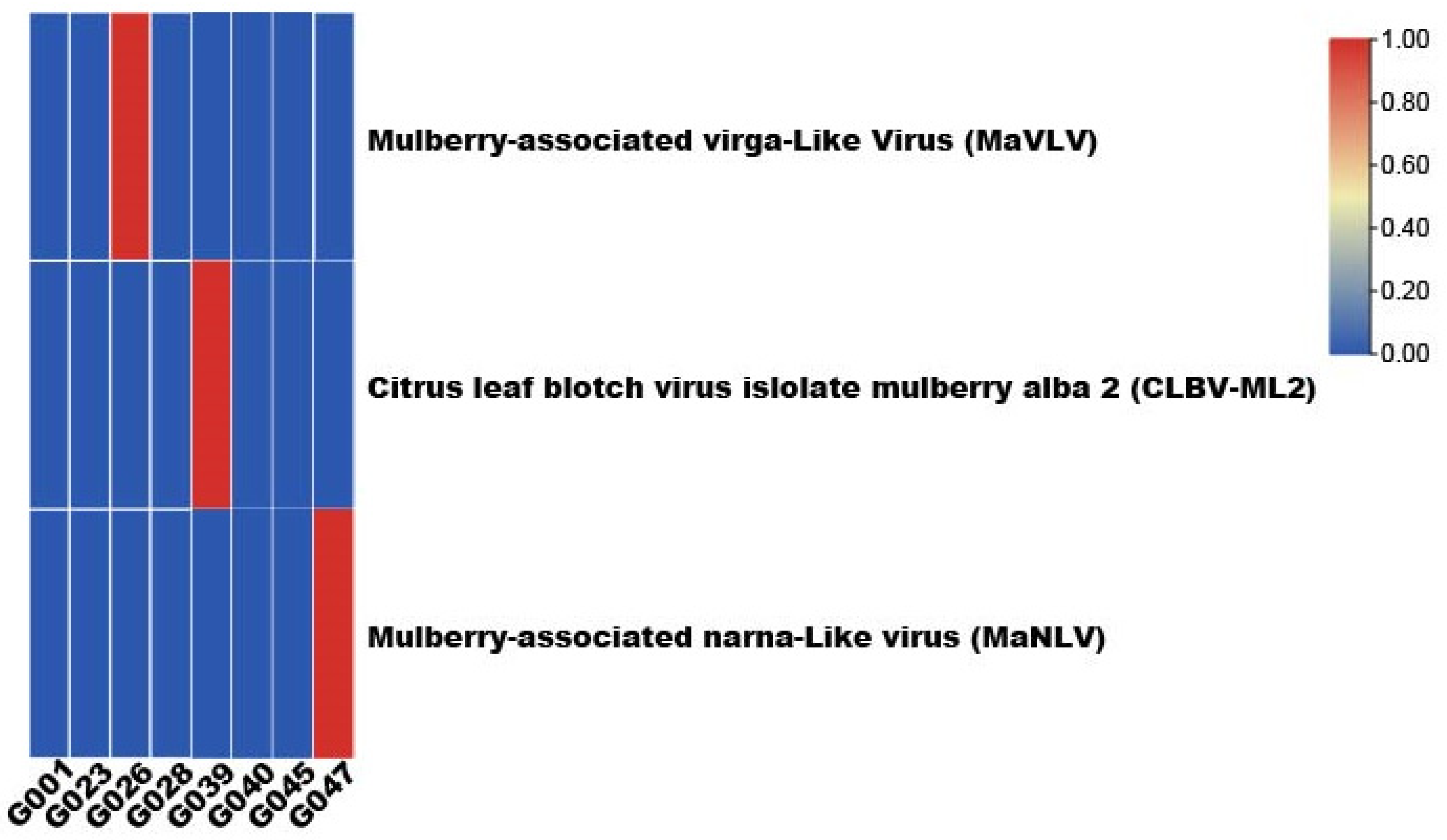
3.3. The Detection of Viruses from Samples and Complete Sequence of CLBV-ML2
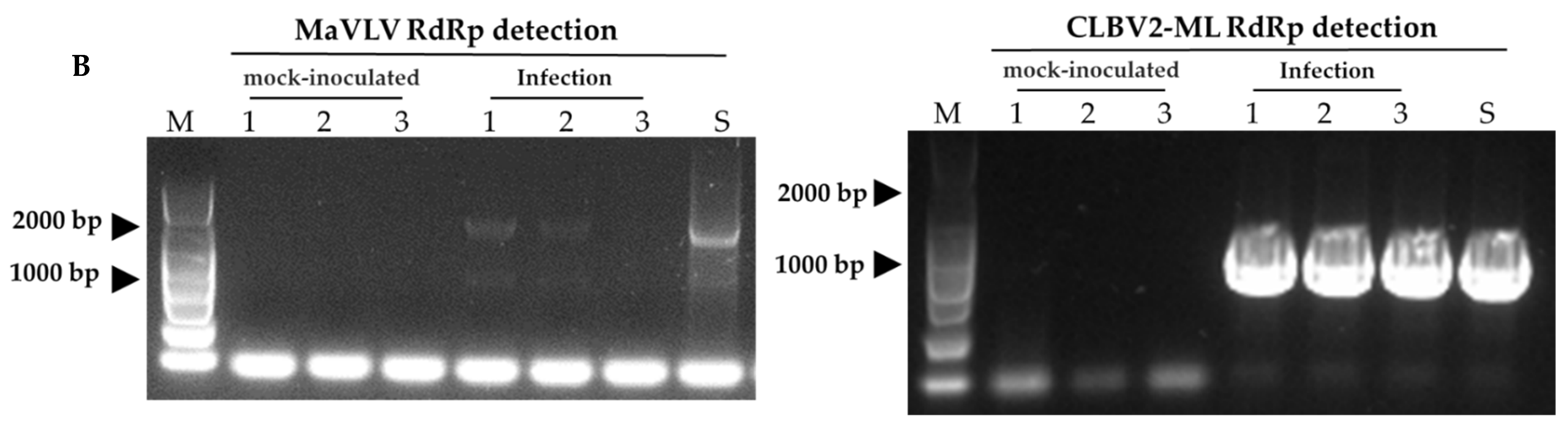
3.4. Viruses Infectivity in N. benthamiana and Morus alba
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mushtaq, M.; Dar, A.; Basu, U.; Bhat, B.; Mir, R.; Vats, S.; Dar, M.; Tyagi, A.; Ali, S.; Bansal, M.; et al. Integrating CRISPR-Cas and Next Generation Sequencing in Plant Virology. Front. Genet. 2021, 12, 735489. [Google Scholar] [CrossRef] [PubMed]
- Villamor, D.; Ho, T.; Rwahnih, M.A.; Martin, R.; Tzanetakis, I. High Throughput Sequencing For Plant Virus Detection and Discovery. Phytopathology 2019, 109, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Carino, E.; Bera, S.; Gao, F.; May, J.; Simon, A. Structural Analysis and Whole Genome Mapping of a New Type of Plant Virus Subviral RNA: Umbravirus-Like Associated RNAs. Viruses 2021, 13, 646. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Bodaghi, S.; Dang, T.; Gadhave, K.; Ho, T.; Osman, F.; Rwahnih, M.A.; Tzanetakis, I.; Simon, A.; Vidalakis, G. Complete Nucleotide Sequence, Genome Organization, and Comparative Genomic Analyses of Citrus Yellow-Vein Associated Virus (CYVaV). Front. Microbiol. 2021, 12, 683130. [Google Scholar] [CrossRef]
- Xie, H.H.; Wei, J.G.; Liu, F.; Pan, X.H.; Yang, X.B. First Report of Mulberry Root Rot Caused by Lasiodiplodia theobromae in China. Plant Dis. 2014, 98, 1581. [Google Scholar] [CrossRef]
- Jian, Q.; Ningjia, H.; Yong, W.; Zhonghuai, X. Ecological issues of mulberry and sustainable development. J. Resour. Ecol. 2012, 3, 330–339. [Google Scholar] [CrossRef]
- Yang, P.; Yang, F.; Tong, Z.; Guan, B.; Li, X.; Ming, M. Research and development of mulberry leaf tea. Seric. Tea Newsl. 2004, 28–30. [Google Scholar]
- Ren, J.; Mao, L.; WEI, T.; Feng, N.; Zheng, H. Study on technology of mulberry leaf juice beverage. Sci. Technol. Food Ind. 2001, 22, 2. [Google Scholar]
- Wang, H. Processing technology of mulberry juice beverage. Anhui Agric. Sci. 2005, 112. [Google Scholar]
- Tang, H.; Xie, Y. Study on processing technology of mulberry wine. Liquor.-Mak. Technol. 2004, 61–62. [Google Scholar]
- Lu, Q.Y.; Wu, Z.J.; Xia, Z.S.; Xie, L.H. Complete genome sequence of a novel monopartite geminivirus identified in mulberry (Morus alba L.). Arch. Virol. 2015, 160, 2135–2138. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zhang, S.; Jin, J.; Xie, J.; Cao, Y.; Cao, M. Molecular characterization of a novel cytorhabdovirus associated with paper mulberry mosaic disease. Arch. Virol. 2020, 165, 2703–2707. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Navarro, B.; Zhang, Z.; Lu, M.; Zhou, X.; Chi, S.; Di Serio, F.; Li, S. Identification and molecular characterization of a novel monopartite geminivirus associated with mulberry mosaic dwarf disease. J. Gen. Virol. 2015, 96, 2421–2434. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Peng, Y.; Kong, W. Identification and complete genome sequence of mulberry cryptic virus 1. Arch. Virol. 2022, 167, 687–690. [Google Scholar] [CrossRef]
- Gürcan, K.; Turan, S.; Teber, S.; Kılınçer, İ.; Uz, I.; Tamisier, L.; Massart, S.; Çağlayan, K. Molecular and biological characterization of a new mulberry idaeovirus. Virus Res. 2021, 298, 198411. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Z.; Xie, J.; Yu, H.; Zhang, S.; Li, R.; Cao, M. Mulberry (Morus alba) is a new natural host of Citrus leaf blotch virus in China. Plant Dis. 2020, 105, 716. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zhang, S.; Yu, H.; Xuan, Z.; Yang, L.; Zhan, B.; Murilo Zerbini, F.; Cao, M. Identification and characterization of two novel geminiviruses associated with paper mulberry (Broussonetia papyrifera) leaf curl disease. Plant Dis. 2020, 104, 3010–3018. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Nerva, L.; Varese, G.C.; Turina, M. Different Approaches to Discover Mycovirus Associated to Marine Organisms. Methods Mol. Biol. Clifton N.J. 2018, 1746, 97–114. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.J.; Adkins, S.; Bragard, C.; Gilmer, D.; Li, D.; MacFarlane, S.A.; Wong, S.M.; Melcher, U.; Ratti, C.; Ryu, K.H.; et al. ICTV Virus Taxonomy Profile: Virgaviridae. J. Gen. Virol. 2017, 98, 1999–2000. [Google Scholar] [CrossRef]
- Adams, M.J.; Antoniw, J.F.; Kreuze, J. Virgaviridae: A new family of rod-shaped plant viruses. Arch. Virol. 2009, 154, 1967–1972. [Google Scholar] [CrossRef] [Green Version]
- DeRisi, J.L.; Huber, G.; Kistler, A.; Retallack, H.; Wilkinson, M.; Yllanes, D. An exploration of ambigrammatic sequences in narnaviruses. Sci. Rep. 2019, 9, 17982. [Google Scholar] [CrossRef] [Green Version]
- Retallack, H.; Popova, K.D.; Laurie, M.T.; Sunshine, S.; DeRisi, J.L. Persistence of ambigrammatic narnaviruses requires translation of the reverse open reading frame. J. Virol. 2021, 95, e00109–e00121. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, D.; Tan, Y.; Zong, X.; Wei, H.; Liu, Q. First Report of Citrus leaf blotch virus in Sweet Cherry. Plant Dis. 2016, 100, 1027. [Google Scholar] [CrossRef]
- Chavan, R.R.; Blouin, A.G.; Cohen, D.; Pearson, M.N. Characterization of the complete genome of a novel citrivirus infecting Actinidia chinensis. Arch. Virol. 2013, 158, 1679–1686. [Google Scholar] [CrossRef]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Viral vectors for the expression of proteins in plants. Curr. Opin. Biotechnol. 2007, 18, 134–141. [Google Scholar] [CrossRef]
- Guerri, J.; Pina, J.A.; Vives, M.C.; Navarro, L.; Moreno, P. Seed Transmission of Citrus leaf botch virus: Implications in Quarantine and Certification Programs. Plant Dis. 2004, 88, 906. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, K.; Agüero, J.; Vives, M.C.; Aleza, P.; Pina, J.A.; Moreno, P.; Navarro, L.; Guerri, J. Precocious flowering of juvenile citrus induced by a viral vector based on Citrus leaf blotch virus: A new tool for genetics and breeding. Plant Biotechnol. J. 2016, 14, 1976–1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agüero, J.; Ruiz-Ruiz, S.; Del-Carmen, M.V.; Velázquez, K.; Navarro, L.; Peña, L.; Moreno, P.; Guerri, J. Development of viral vectors based on Citrus leaf blotch virus to express foreign proteins or analyze gene function in citrus plants. Mol. Plant-Microbe Interact. 2012, 25, 1326–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agüero, J.; Vives, M.C.; Velázquez, K.; Pina, J.A.; Navarro, L.; Moreno, P.; Guerri, J. Effectiveness of gene silencing induced by viral vectors based on Citrus leaf blotch virus is different in Nicotiana benthamiana and citrus plants. Virology 2014, 460–461, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Dawson, W.O.; Bar-Joseph, M.; Garnsey, S.M.; Moreno, P. Citrus tristeza virus: Making an ally from an enemy. Annu. Rev. Phytopathol. 2015, 53, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Kamencayová, M.; Košík, I.; Hunková, J.; Subr, Z.W. Transient expression of the influenza A virus PB1-F2 protein using a plum pox virus-based vector in Nicotiana benthamiana. Acta Virol. 2014, 58, 274–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Sasaki, N.; Isogai, M.; Yoshikawa, N. Stable expression of foreign proteins in herbaceous and apple plants using Apple latent spherical virus RNA2 vectors. Arch. Virol. 2004, 149, 1541–1558. [Google Scholar] [CrossRef] [PubMed]
- Maliogka, V.I.; Minafra, A.; Saldarelli, P.; Ruiz-García, A.B.; Glasa, M.; Katis, N.; Olmos, A. Recent Advances on Detection and Characterization of Fruit Tree Viruses Using High-Throughput Sequencing Technologies. Viruses 2018, 10, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muruganantham, M.; Moskovitz, Y.; Haviv, S.; Horesh, T.; Fenigstein, A.; Preez, J.D.; Stephan, D.; Burger, J.T.; Mawassi, M. Grapevine virusA-mediated gene silencing in Nicotiana benthamiana and Vitis vinifera. J. Virol. Methods 2009, 155, 167–174. [Google Scholar] [CrossRef]
- Okada, R.; Ichinose, S.; Takeshita, K.; Urayama, S.I.; Fukuhara, T.; Komatsu, K.; Arie, T.; Ishihara, A.; Egusa, M.; Kodama, M.; et al. Molecular characterization of a novel mycovirus in Alternaria alternata manifesting two-sided effects: Down-regulation of host growth and up-regulation of host plant pathogenicity. Virology 2018, 519, 23–32. [Google Scholar] [CrossRef]
- Sasaki, S.; Yamagishi, N.; Yoshikawa, N. Efficient virus-induced gene silencing in apple, pear and Japanese pear using Apple latent spherical virus vectors. Plant Methods 2011, 7, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simkovich, A.J.; Li, Y.; Kohalmi, S.E.; Griffiths, J.S.; Wang, A. Molecular Identification of Prune Dwarf Virus (PDV) Infecting Sweet Cherry in Canada and Development of a PDV Full-Length Infectious cDNA Clone. Viruses 2021, 13, 2025. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Name | Province | Years | Latitude and Longitude | CLBV-ML2 | MaVLV | MaNLV |
|---|---|---|---|---|---|---|---|
| G001 | G-HEBBS2H | Heilongjiang | 600 | 126°54′ N 45°80′ E | - | - | - |
| G023 | G-QXGGZXZ | Henan | 500 | 114°68′ N 34°58′ E | - | - | - |
| G026 | G-NYXYSYZ | Henan | 1800 | 112°46′ N 32°61′ E | - | + | - |
| G028 | G-XJTL | Shandong | 1200 | 116°01′ N 36°98′ E | - | - | - |
| G039 | G-HSSX | Anhui | 800 | 118°42′ N 29°86′ E | + | - | - |
| G040 | G-BJDX1 | Beijing | 145 | 116°34′ N 39°73′ E | - | - | - |
| G045 | G-NBQZ3 | Xinjiang | 425 | 121°55′ N 29°82′ E | - | - | - |
| G047 | G-SXXC2 | Xizang | 1600 | 120°91′ N 29°50′ E | - | - | + |
| Name * | Genome | ORF1 (Rep) | ORF2 (MP) | ORF3 (CP) |
|---|---|---|---|---|
| CLBV-PY2 | 83.08/8792 nt | 82.44 (88.11) | 82.72 (95.58) | 84.11 (94.21) |
| Dweet mottle virus | 80.33/8747 nt | 79.23 (78.63) | n.a. (97.79) | 84.11 (94.77) |
| CLBV Strain Actinidia | 81.95/8793 nt | 81.07 (88.81) | n.a. (94.75) | 84.59 (95.04) |
| CLBV | 78.19/8747 nt | 75.97 (78.00) | n.a. (98.34) | 84.38 (95.32) |
| CLBV isolate ML | 81.87/8776 nt | 81.07 (87.29) | n.a. (95.58) | n.a. (88.43) |
| CLBV2 | 78.82/8697 nt | n.a. (58.07) | n.a. (90.61) | n.a. (93.66) |
| Humulus lupulus citrivirus 1 | 76.637181 nt | 75.44 (63.95) | 79.76 (86.19) | n.a. (90.61) |
| CLBV strain Prunus | 78.17/8762 nt | n.a. (76.52) | n.a. (95.86) | n.a. (87.88) |
| Paeonia citrivirus A | 79.28/6451 nt | n.a. (59.77) | 84.02 (98.07) | 83.12 (95.27) |
| Peony yellowing-associated Citrivirus | 80.28/8666 nt | n.a. (53.35) | n.a. (96.13) | n.a. (88.71) |
| Nandina Citrivirus | 77.74/8892 nt | 77.66 (81.18) | n.a. (58.07) | n.a. (78.51) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Xu, Z.-L.; Liu, P.-G.; Zhu, Y.; Lin, T.-B.; Li, T.-Y.; Lv, Z.-Q.; Wei, J. Identification of Three Viruses Infecting Mulberry Varieties. Viruses 2022, 14, 2564. https://doi.org/10.3390/v14112564
Chen L, Xu Z-L, Liu P-G, Zhu Y, Lin T-B, Li T-Y, Lv Z-Q, Wei J. Identification of Three Viruses Infecting Mulberry Varieties. Viruses. 2022; 14(11):2564. https://doi.org/10.3390/v14112564
Chicago/Turabian StyleChen, Lei, Zi-Long Xu, Pei-Gang Liu, Yan Zhu, Tian-Bao Lin, Tian-Yan Li, Zhi-Qiang Lv, and Jia Wei. 2022. "Identification of Three Viruses Infecting Mulberry Varieties" Viruses 14, no. 11: 2564. https://doi.org/10.3390/v14112564
APA StyleChen, L., Xu, Z. -L., Liu, P. -G., Zhu, Y., Lin, T. -B., Li, T. -Y., Lv, Z. -Q., & Wei, J. (2022). Identification of Three Viruses Infecting Mulberry Varieties. Viruses, 14(11), 2564. https://doi.org/10.3390/v14112564