

Bacteriophage Cocktails in the Post-COVID Rehabilitation
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteriophages
2.2. Bacterial Strains
2.3. In Vitro Real-Time Phage Lysis Assay
2.4. Participants
- Post-COVID patients of moderate severity (incl. ICU stay), discharged from the hospital more than 3 months ago;
- Presence of pulmonary foci of consolidation and fibrosis on chest CT (CT 1-3);
- Rehabilitation routing score—less than 3 points;
- Negative polymerase chain reaction (PCR) result for SARS-CoV-2.
- Exclusion criteria:
- Temperature above 38 °C;
- Increased dyspnea (above 30/min);
- Increase in systolic blood pressure above 180 mmHg or a decrease below 90 mmHg.
2.5. Sample Collection
2.6. Analysis of Gut Microbiota Taxonomic Abundance
2.7. CT Scan Analysis
3. Results
3.1. Experimental Part
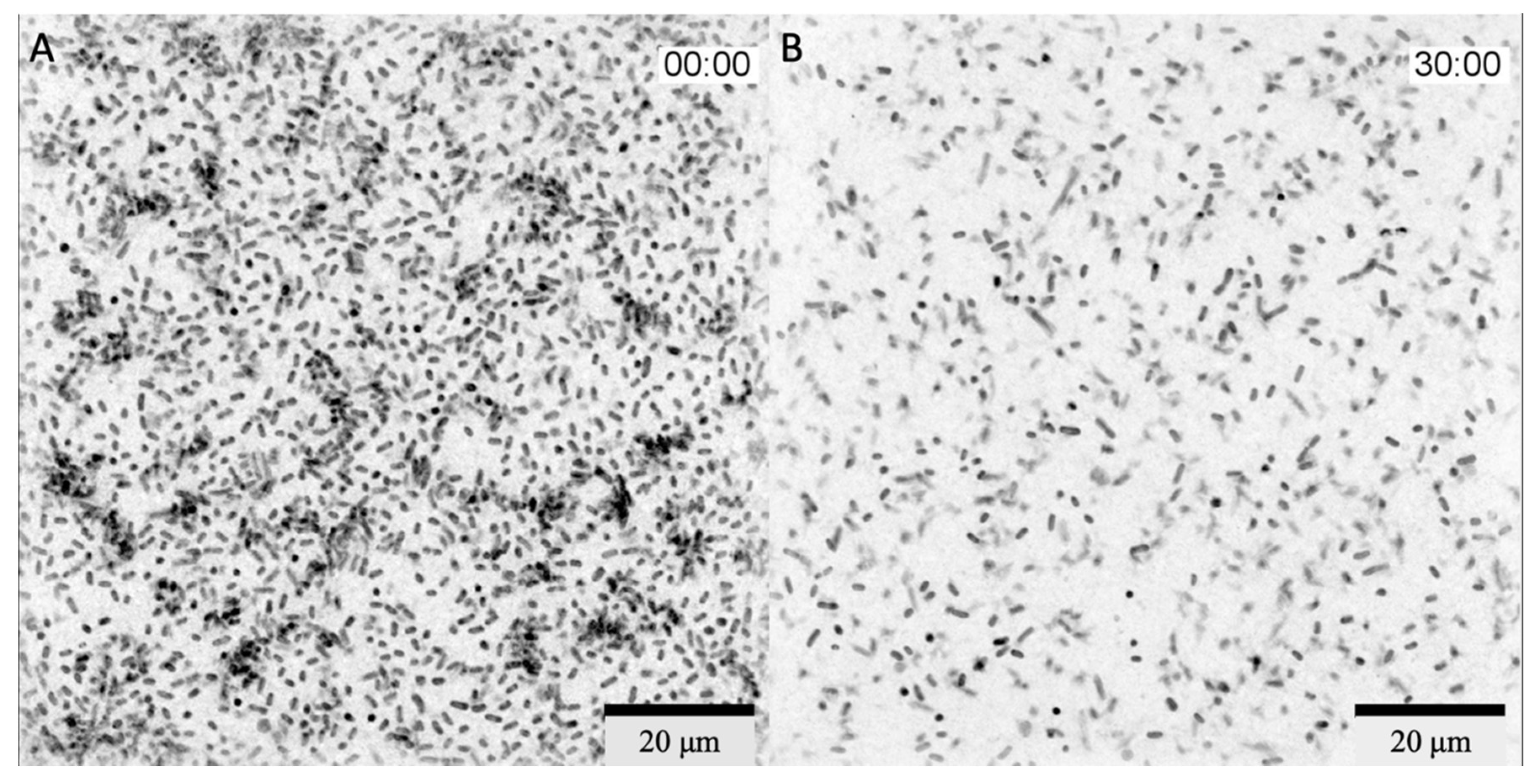
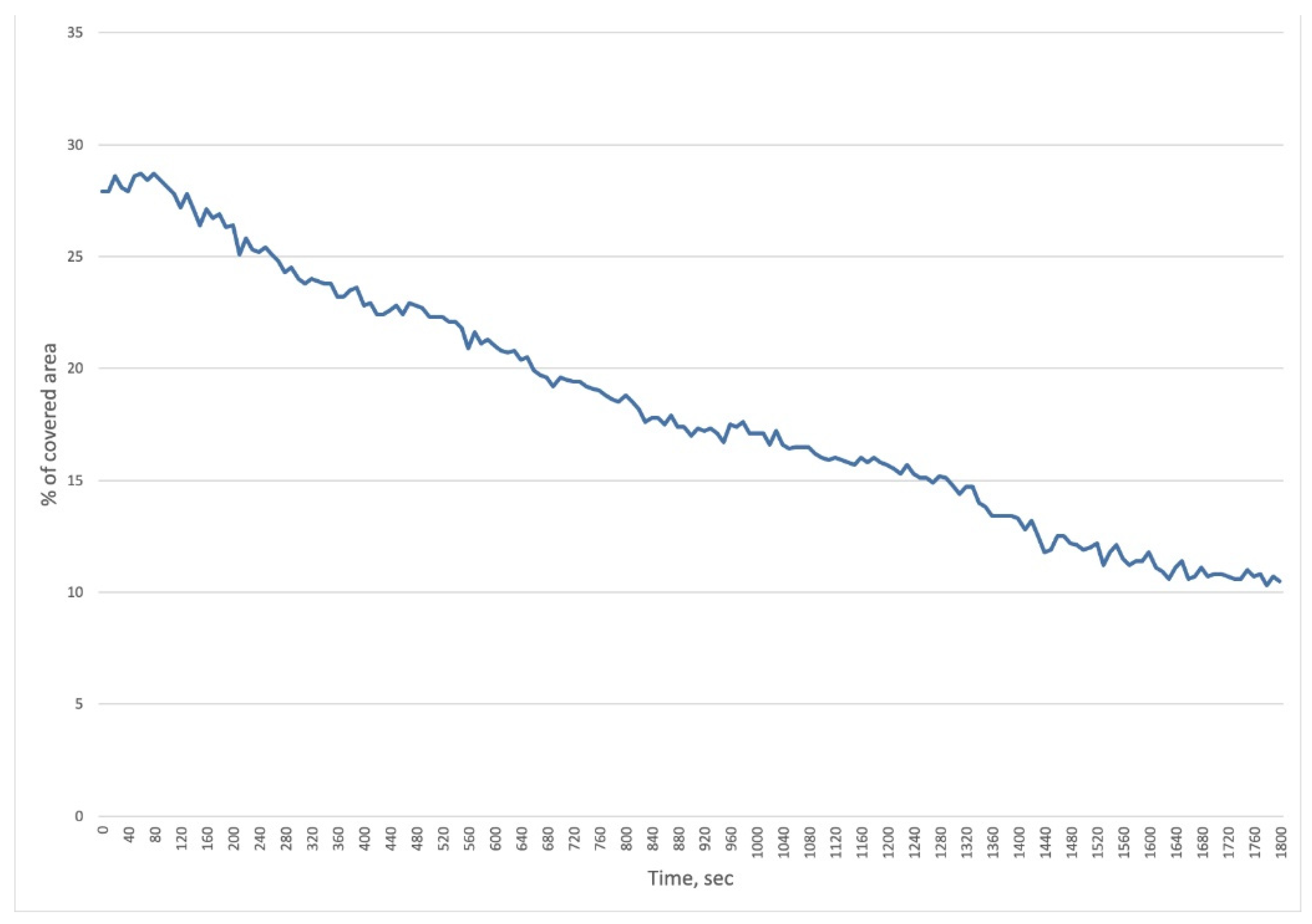
In Vitro Real-Time Phage Lysis Assay
3.2. Clinical Phage Assessment
3.2.1. Subjects Characteristics
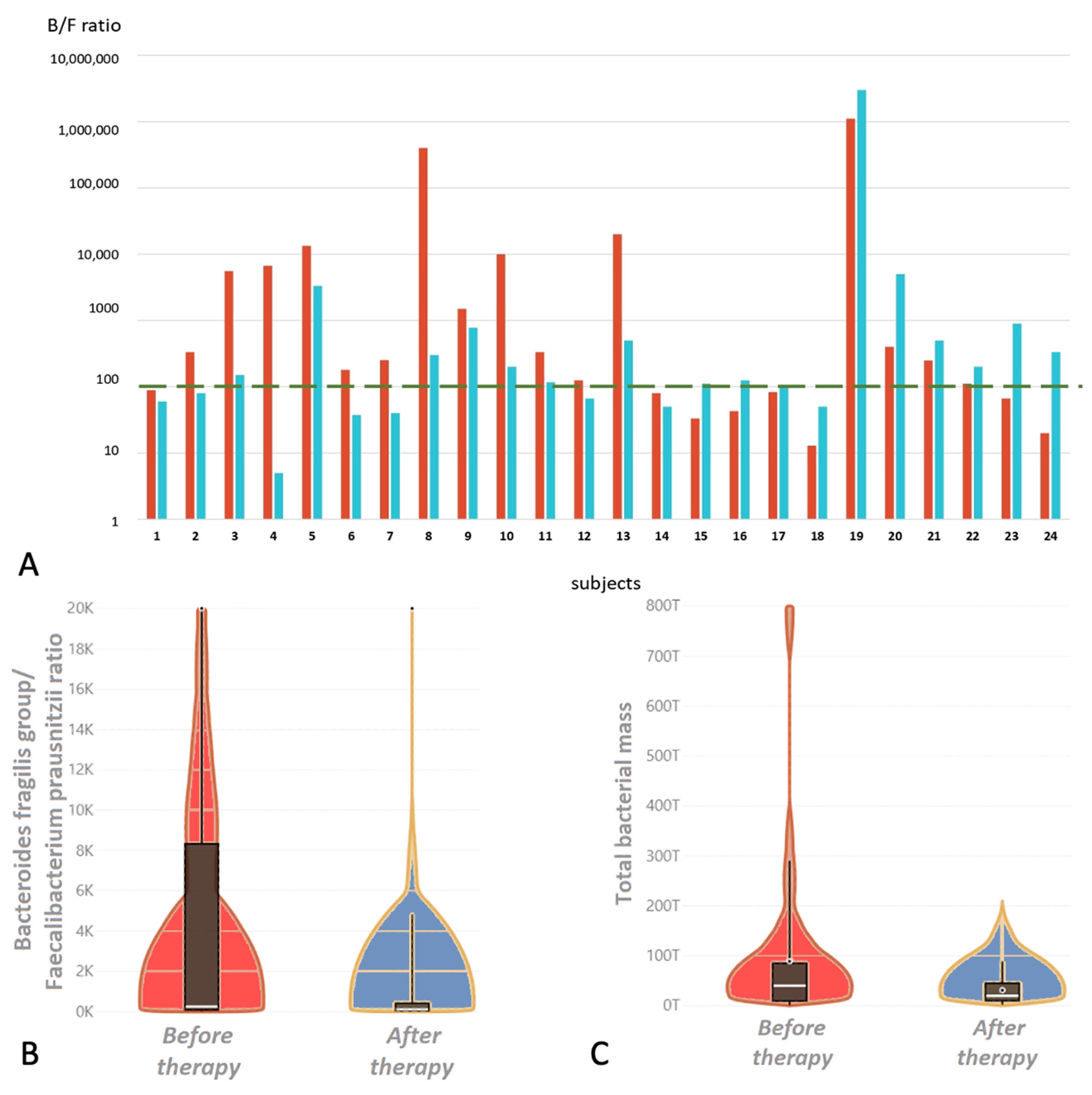
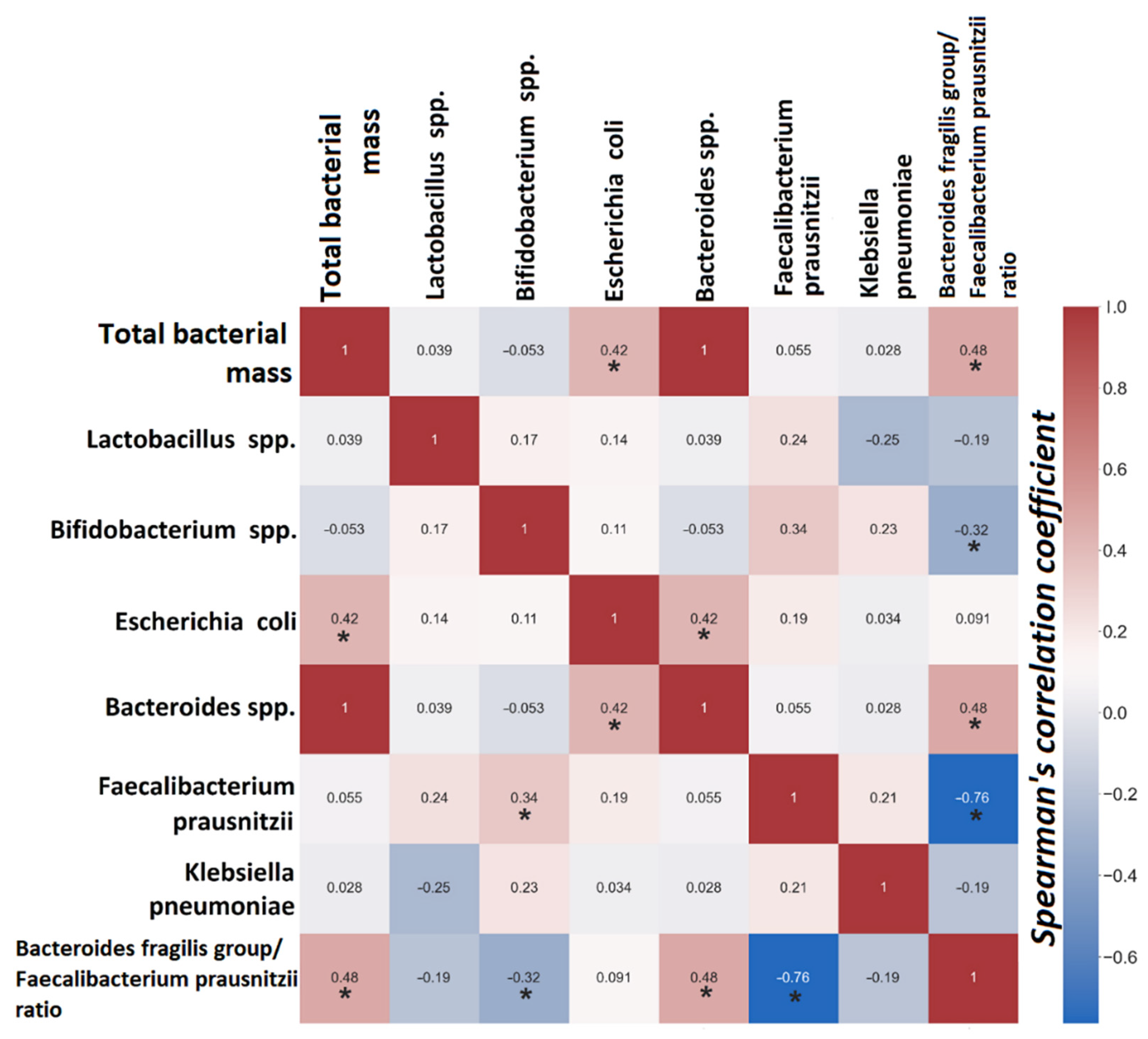
3.2.2. Gut Dysbiosis
4. Discussion
Clinical Part
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, H.; Liu, M.; Cao, J.; Li, X.; Fan, D.; Xia, Y.; Lu, X.; Li, J.; Ju, D.; Zhao, H. The Dynamic Interplay between the Gut Microbiota and Autoimmune Diseases. J. Immunol. Res. 2019, 2019, 7546047. [Google Scholar] [CrossRef] [Green Version]
- Morais, L.H.; Schreiber, H.L., 4th; Mazmanian, S.K. The gut microbiota-brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Zhang, F.; Wan, Y.; Zuo, T.; Yeoh, Y.K.; Liu, Q.; Zhang, L.; Zhan, H.; Lu, W.; Xu, W.; Lui, G.C.Y.; et al. Prolonged Impairment of Short-Chain Fatty Acid and L-Isoleucine Biosynthesis in Gut Microbiome in Patients with COVID-19. Gastroenterology 2022, 162, 548–561.e4. [Google Scholar] [CrossRef]
- Liu, Q.; Mak, J.; Su, Q.; Yeoh, Y.K.; Lui, G.C.; Ng, S.; Zhang, F.; Li, A.; Lu, W.; Hui, D.S.; et al. Gut microbiota dynamics in a prospective cohort of patients with post-acute COVID-19 syndrome. Gut 2022, 71, 544–552. [Google Scholar] [CrossRef]
- Rawson, T.M.; Moore, L.; Castro-Sanchez, E.; Charani, E.; Davies, F.; Satta, G.; Ellington, M.J.; Holmes, A.H. COVID-19 and the potential long-term impact on antimicrobial resistance. J. Antimicrob. Chemother. 2020, 75, 1681–1684. [Google Scholar] [CrossRef]
- Fernández, L.; Gutiérrez, D.; Rodríguez, A.; García, P. Application of Bacteriophages in the Agro-Food Sector: A Long Way Toward Approval. Front. Cell. Infect. Microbiol. 2018, 8, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordillo Altamirano, F.L.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Van Belleghem, J.D.; de Vries, C.R.; Burgener, E.; Chen, Q.; Manasherob, R.; Aronson, J.R.; Amanatullah, D.F.; Tamma, P.D.; Suh, G.A. The Safety and Toxicity of Phage Therapy: A Review of Animal and Clinical Studies. Viruses 2021, 13, 1268. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Mao, J.; Xie, J. Bacteriophage polysaccharide depolymerases and biomedical applications. BioDrugs 2014, 28, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Leiman, P.G.; Battisti, A.J.; Bowman, V.D.; Stummeyer, K.; Mühlenhoff, M.; Gerardy-Schahn, R.; Scholl, D.; Molineux, I.J. The structures of bacteriophages K1E and K1-5 explain processive degradation of polysaccharide capsules and evolution of new host specificities. J. Mol. Biol. 2007, 371, 836–849. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.M.; Bachman, M.A. Colonization, Infection, and the Accessory Genome of Klebsiella pneumoniae. Front. Cell. Infect. Microbiol. 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taur, Y.; Xavier, J.B.; Lipuma, L.; Ubeda, C.; Goldberg, J.; Gobourne, A.; Lee, Y.J.; Dubin, K.A.; Socci, N.D.; Viale, A.; et al. Intestinal domination and the risk of bacteremia in patients undergoing allogeneic hematopoietic stem cell transplantation. Clin. Infect. Dis. 2012, 55, 905–914. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wu, C.; Zhang, S.; Zhong, Y. Risk Factors of Ventilator-Associated Pneumonia in Critically III Patients. Front. Pharmacol. 2019, 10, 482. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yao, Z.; Zhan, S.; Yang, Z.; Wei, D.; Zhang, J.; Li, J.; Kyaw, M.H. Disease burden of intensive care unit-acquired pneumonia in China: A systematic review and meta-analysis. Int. J. Infect. Dis. 2014, 29, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Ge, Y.; Wu, T.; Zhao, K.; Chen, Y.; Wu, B.; Zhu, F.; Zhu, B.; Cui, L. Co-infection with respiratory pathogens among COVID-2019 cases. Virus Res. 2020, 285, 198005. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, R.; Goodarzi, P.; Asadi, M.; Soltani, A.; Aljanabi, H.A.A.; Jeda, A.S.; Dashtbin, S.; Jalalifar, S.; Mohammadzadeh, R.; Teimoori, A.; et al. Bacterial co-infections with SARS-CoV-2. IUBMB Life 2020, 72, 2097–2111. [Google Scholar] [CrossRef]
- Chong, Y.; Yakushiji, H.; Ito, Y.; Kamimura, T. Clinical and molecular epidemiology of extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae in a long-term study from Japan. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 83–87. [Google Scholar] [CrossRef]
- Munoz-Price, L.S.; Poirel, L.; Bonomo, R.A.; Schwaber, M.J.; Daikos, G.L.; Cormican, M.; Cornaglia, G.; Garau, J.; Gniadkowski, M.; Hayden, M.K.; et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect. Dis. 2013, 13, 785–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, S.; Reuter, S.; Harris, S.R.; Glasner, C.; Feltwell, T.; Argimon, S.; Abudahab, K.; Goater, R.; Giani, T.; Errico, G.; et al. Epidemic of carbapenem-resistant Klebsiella pneumoniae in Europe is driven by nosocomial spread. Nat. Microbiol. 2019, 4, 1919–1929. [Google Scholar] [CrossRef]
- Pitout, J.D.; Nordmann, P.; Poirel, L. Carbapenemase-Producing Klebsiella pneumoniae, a Key Pathogen Set for Global Nosocomial Dominance. Antimicrob. Agents Chemother. 2015, 59, 5873–5884. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, S.M.; Cardoso, M.H.; Cândido Ede, S.; Franco, O.L. Understanding, preventing and eradicating Klebsiella pneumoniae biofilms. Future Microbiol. 2016, 11, 527–538. [Google Scholar] [CrossRef]
- Febvre, H.P.; Rao, S.; Gindin, M.; Goodwin, N.D.M.; Finer, E.; Vivanco, J.S.; Lu, S.; Manter, D.K.; Wallace, T.C.; Weir, T.L. PHAGE Study: Effects of Supplemental Bacteriophage Intake on Inflammation and Gut Microbiota in Healthy Adults. Nutrients 2019, 11, 666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francino, M.P. Antibiotics and the Human Gut Microbiome: Dysbioses and Accumulation of Resistances. Front. Microbiol. 2016, 6, 1543. [Google Scholar] [CrossRef] [Green Version]
- Zurabov, F.; Zhilenkov, E. Characterization of four virulent Klebsiella pneumoniae bacteriophages, and evaluation of their potential use in complex phage preparation. Virol. J. 2021, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- Beloborodova, N.V.; Grechko, A.V.; Gurkova, M.M.; Zurabov, A.Y.; Zurabov, F.M.; Kuzovlev, A.N.; Megley, A.Y.; Petrova, M.V.; Popova, V.M.; Redkin, I.V.; et al. Adaptive Phage Therapy in the Treatment of Patients with Recurrent Pneumonia (Pilot Study). Gen. Reanimatol. 2021, 17, 4–14. [Google Scholar] [CrossRef]
- Galtier, M.; De Sordi, L.; Maura, D.; Arachchi, H.; Volant, S.; Dillies, M.A.; Debarbieux, L. Bacteriophages to reduce gut carriage of antibiotic resistant uropathogens with low impact on microbiota composition. Environ. Microbiol. 2016, 18, 2237–2245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, A.; Wu, M.; McNulty, N.P.; Rohwer, F.L.; Gordon, J.I. Gnotobiotic mouse model of phage-bacterial host dynamics in the human gut. Proc. Natl. Acad. Sci. USA 2013, 110, 20236–20241. [Google Scholar] [CrossRef] [Green Version]
- Hsu, B.B.; Gibson, T.E.; Yeliseyev, V.; Liu, Q.; Lyon, L.; Bry, L.; Silver, P.A.; Gerber, G.K. Dynamic Modulation of the Gut Microbiota and Metabolome by Bacteriophages in a Mouse Model. Cell Host Microbe 2019, 25, 803–814.e5. [Google Scholar] [CrossRef] [Green Version]
- Mu, A.; McDonald, D.; Jarmusch, A.K.; Martino, C.; Brennan, C.; Bryant, M.; Humphrey, G.C.; Toronczak, J.; Schwartz, T.; Nguyen, D.; et al. Assessment of the microbiome during bacteriophage therapy in combination with systemic antibiotics to treat a case of staphylococcal device infection. Microbiome 2021, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Slopek, S.; Weber-Dabrowska, B.; Dabrowski, M.; Kucharewicz-Krukowska, A. Results of bacteriophage treatment of suppurative bacterial infections in the years 1981–1986. Arch. Immunol. Ther. Exp. (Warsz) 1987, 35, 569–583. [Google Scholar] [PubMed]
- Petrovic Fabijan, A.; Lin, R.C.Y.; Ho, J.; Maddocks, S.; Ben Zakour, N.L.; Iredell, J.R.; Westmead Bacteriophage Therapy Team. Safety of bacteriophage therapy in severe Staphylococcus aureus infection. Nat. Microbiol. 2020, 5, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Chen, Y.; Wu, Z.; Chen, Y.; Gao, H.; Lv, L.; Guo, F.; Zhang, X.; Luo, R.; Huang, C.; et al. Alterations of the Gut Microbiota in Patients with Coronavirus Disease 2019 or H1N1 Influenza. Clin. Infect. Dis. 2020, 71, 2669–2678. [Google Scholar] [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.Y.; Yeoh, Y.K.; Li, A.Y.L.; Zhan, H.; Wan, Y.; Chung, A.C.K.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients with COVID-19 during Time of Hospitalization. Gastroenterology 2020, 159, 944–955.e8. [Google Scholar] [CrossRef]
- Yeoh, Y.K.; Zuo, T.; Lui, G.C.; Zhang, F.; Liu, Q.; Li, A.Y.; Chung, A.C.; Cheung, C.P.; Tso, E.Y.; Fung, K.S.; et al. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. Gut 2021, 70, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Chernevskaya, E.A.; Meglei, A.Y.; Buyakova, I.V.; Kovaleva, N.Y.; Gorshkov, K.M.; Zakharchenko, V.E.; Beloborodova, N.V. Taxonomic dysbiosis of gut microbiota and serum biomarkers reflect severity of central nervous system injury. Bull. Russ. State Med. Univ. 2020, 5, 54–61. [Google Scholar] [CrossRef]
- Miedzybrodzki, R.; Fortuna, W.; Weber-Dabrowska, B.; Górski, A.A. Retrospective analysis of changes in inflammatory markers in patients treated with bacterial viruses. Clin. Exp. Med. 2009, 9, 303–312. [Google Scholar] [CrossRef]
- Barr, J.J.; Auro, R.; Furlan, M.; Whiteson, K.L.; Erb, M.L.; Pogliano, J.; Stotland, A.; Wolkowicz, R.; Cutting, A.S.; Doran, K.S.; et al. Bacteriophage adhering to mucus provide a non-host-derived immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 10771–10776. [Google Scholar] [CrossRef] [Green Version]
- Almeida, G.M.F.; Laanto, E.; Ashrafi, R.; Sundberg, L.R. Bacteriophage Adherence to Mucus Mediates Preventive Protection against Pathogenic Bacteria. mBio 2019, 10, e01984-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Patel, A.; SantaLucia, J.; Roberts, E.; Zhao, L.; Kaye, K.; Rao, K.; Bachman, M.A. Measurement of Klebsiella intestinal colonization density to assess infection risk. mSphere 2021, 6, e00500-21. [Google Scholar] [CrossRef]
- Bryan, D.; El-Shibiny, A.; Hobbs, Z.; Porter, J.; Kutter, E.M. Bacteriophage T4 Infection of Stationary Phase E. coli: Life after Log from a Phage Perspective. Front. Microbiol. 2016, 7, 1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulich, K.R.; Malfertheiner, P.; Madisch, A.; Labenz, J.; Bayerdorffer, E.; Miehlke, S.; Carlsson, J.; Wiklund, I.K. Psychometric validation of the German translation of the Gastrointestinal Symptom Rating Scale (GSRS) and Quality of Life in Reflux and Dyspepsia (QOLRAD) questionnaire in patients with reflux disease. Health Qual. Life Outcomes 2003, 1, 62. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Subjects, Median (Q1; Q3) | p Value | |
|---|---|---|---|
| Before BPh | After BPh | ||
| Saturation, percent | 96 (95; 96) | 98 (98; 99) | p < 0.001 * |
| Respiratory rate (breaths per minute) | 19 (18; 20) | 16 (16; 16) | p < 0.001 * |
| Heart rate (beats per minute) | 78 (74; 82) | 72 (70; 72) | p < 0.001 * |
| Hemoglobin, g/L | 135 (124; 150) | 142 (132; 153) | 0.007 * |
| White blood cells, 109/L | 6 (5; 7) | 5 (5; 8) | 0.436 |
| Erythrocyte Sedimentation Rate (ESR), mm/hour | 13 (8; 23) | 9 (5; 13) | 0.007 * |
| Parameters | Reference Values | Subjects, Median (Q1; Q3) | p Value | |
|---|---|---|---|---|
| Before BPh | After BPh | |||
| Bilirubin | 5–21 µmol/L | 15.6 (11.4; 18.8) | 12.5 (11.2; 18.5) | 0.058 |
| Total protein | 66–83 g/L | 69.1 (66.3; 71.5) | 71.2 (68.4; 72.6) | 0.038 * |
| Creatinine | 74–110 µmol/L | 82 (74; 103) | 89 (80; 98) | 0.657 |
| Glucose | 4.1–5.9 mmol/L | 5.7 (5.1; 5.9) | 5.6 (5.2; 6.0) | 0.236 |
| Cholesterol | 0–5.2 mmol/L | 5.5 (4.5; 7.1) | 5.9 (4.5; 6.2) | 0.154 |
| Lactate dehydrogenase | 0–247 u/L | 198 (168; 223) | 197 (168; 229) | 0.137 |
| Alanine aminotransferase | 0–50 u/L | 21 (17; 32) | 20 (15; 31) | 0.679 |
| Aspartate aminotransferase | 0–50 u/L | 23 (19; 32) | 24 (19; 29.5) | 0.225 |
| C-reactive protein (CRP) | 0–5 mg/L | 0.6 (0.1; 0.9) | 0.12 (0.1; 0.6) | 0.061 |
| Parameters | Reference Values | Subjects, Median (Q1; Q3) | Indicator of Changes | p Value | |
|---|---|---|---|---|---|
| Before BPh | After BPh | ||||
| Total bacterial mass | <1012 | 4 × 1013 (9 × 1012; 9 × 1013) | 2 × 1013 (1013; 6 × 1013) | ⇩ | 0.171 |
| Lactobacillus spp. | 107–108 | 4 × 107 (7 × 105; 5 × 108) | 5 × 107 (8 × 106; 108) | ⇧ | 0.951 |
| Bifidobacterium spp. | 109 –1010 | 3 × 1010 (4 × 109; 7 × 1010) | 2 × 1010 (3 × 109; 6 × 1010) | ⇩ | 0.535 |
| Escherichia coli | 107 –108 | 2 × 108 (107; 109) | 6 × 107 (3 × 107; 6 × 108) | ⇩ | 0.394 |
| Bacteroides spp. | 109 –1012 | 4 × 1013 (9 × 1012; 9 × 1013) | 2 × 1013 (1013; 6 × 1013) | ⇩ | 0.171 |
| Faecalibacterium prausnitzii | 108 –1011 | 9 × 1010 (9 × 109;4 × 1011) | 2 × 1011 (2 × 1010; 6 × 1011) | ⇧ | 0.122 |
| Bacteroides thetaiotaomicron | 109 –1012 | 5 × 109 (4 × 108; 2 × 1010) | 2 × 109 (7 × 108; 1010) | ⇩ | 0.327 |
| Akkermansia muciniphila | <1011 | 2 × 107 (107; 1012) | 5 × 106 (2 × 105; 5 × 108) | ⇩ | 0.93 |
| Enterococcus spp.* | <108 | 4 × 1012 | 5 × 105 | - | |
| Escherichia coli enteropathogenic | <104 | - | - | - | - |
| Klebsiella pneumoniae | <104 | <105 | <105 | - | - |
| Klebsiella oxytoca | <104 | - | - | - | - |
| Candida spp. | <104 | 2 × 106 (3 × 105; 2 × 107) | 2 × 106 (3 × 105; 3 × 106) | - | 0.23 |
| Staphylococcus aureus | <104 | 2 × 107 (8 × 105; 4 × 107) | 7 × 105 (105; 2 × 106) | ⇩ | 0.999 |
| Clostridium difficile * | - | 3 × 107 | 105 | ⇩ | - |
| Clostridium perfringens | - | 3 × 106 (2 × 106; 107) | 3 × 105 (105; 3 × 106) | ⇩ | 0.23 |
| Proteus vulgaris/mirabilis | <104 | 107 (107; 107) | 2 × 106 (4 × 105; 5 × 106) | ⇩ | 0.18 |
| Citrobacter spp. | <104 | 108 (2 × 105; 5 × 1013) | - | ⇩ | - |
| Enterobacter spp. | <104 | 3 × 106 (8 × 105; 3 × 107) | 107 (4 × 106;108) | ⇧ | 0.53 |
| Fusobacterium nucleatum | - | 4.5 × 105 (105; 7 × 105) | 4 × 105 (105; 7 × 105) | - | 1.00 |
| Parvimonas micra | - | 3 × 105 (105; 4 × 105) | 9 × 105 (6 × 105; 106) | - | 0.19 |
| Bacteroides fragilis group/Faecalibacterium prausnitzii Ratio | 0.1–100 | 252 (80; 10,000) | 111 (50; 500) | ⇩ | 0.033 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zurabov, F.M.; Chernevskaya, E.A.; Beloborodova, N.V.; Zurabov, A.Y.; Petrova, M.V.; Yadgarov, M.Y.; Popova, V.M.; Fatuev, O.E.; Zakharchenko, V.E.; Gurkova, M.M.; et al. Bacteriophage Cocktails in the Post-COVID Rehabilitation. Viruses 2022, 14, 2614. https://doi.org/10.3390/v14122614
Zurabov FM, Chernevskaya EA, Beloborodova NV, Zurabov AY, Petrova MV, Yadgarov MY, Popova VM, Fatuev OE, Zakharchenko VE, Gurkova MM, et al. Bacteriophage Cocktails in the Post-COVID Rehabilitation. Viruses. 2022; 14(12):2614. https://doi.org/10.3390/v14122614
Chicago/Turabian StyleZurabov, Fedor M., Ekaterina A. Chernevskaya, Natalia V. Beloborodova, Alexander Yu. Zurabov, Marina V. Petrova, Mikhail Ya. Yadgarov, Valentina M. Popova, Oleg E. Fatuev, Vladislav E. Zakharchenko, Marina M. Gurkova, and et al. 2022. "Bacteriophage Cocktails in the Post-COVID Rehabilitation" Viruses 14, no. 12: 2614. https://doi.org/10.3390/v14122614
APA StyleZurabov, F. M., Chernevskaya, E. A., Beloborodova, N. V., Zurabov, A. Y., Petrova, M. V., Yadgarov, M. Y., Popova, V. M., Fatuev, O. E., Zakharchenko, V. E., Gurkova, M. M., Sorokina, E. A., Glazunov, E. A., Kochetova, T. A., Uskevich, V. V., Kuzovlev, A. N., & Grechko, A. V. (2022). Bacteriophage Cocktails in the Post-COVID Rehabilitation. Viruses, 14(12), 2614. https://doi.org/10.3390/v14122614