

Successful Rescue of Synthetic AcMNPV with a ~17 kb Deletion in the C1 Region of the Genome
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Construction and Synthesis of Modified Genomes
2.3. Transfection and Infection Assays
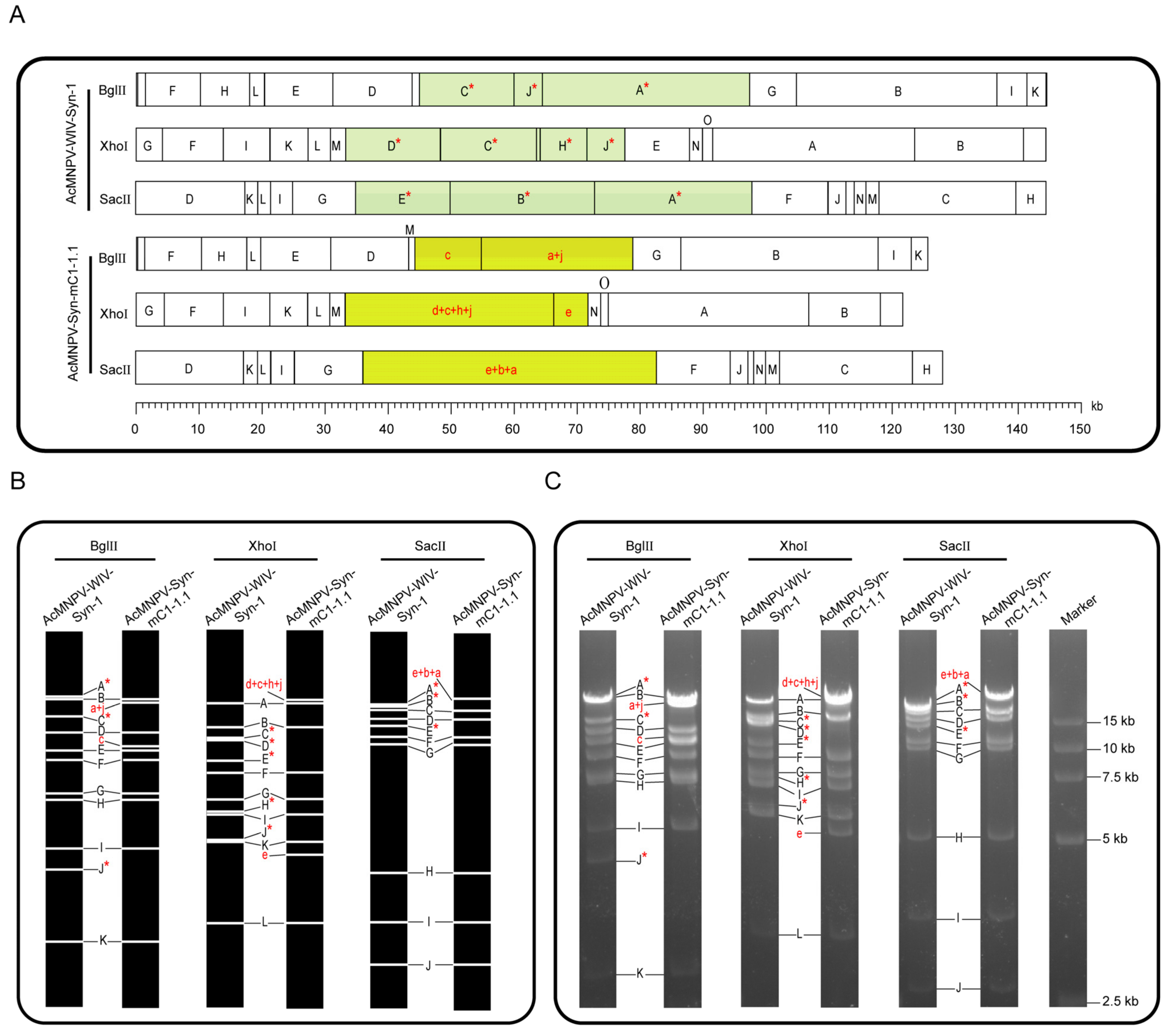
2.4. Restriction Enzyme Digestion Analysis and Genome Sequencing of AcMNPV-Syn-mC1-1.1
2.5. One-Step Growth Curve
2.6. Electron Microscopy
3. Results
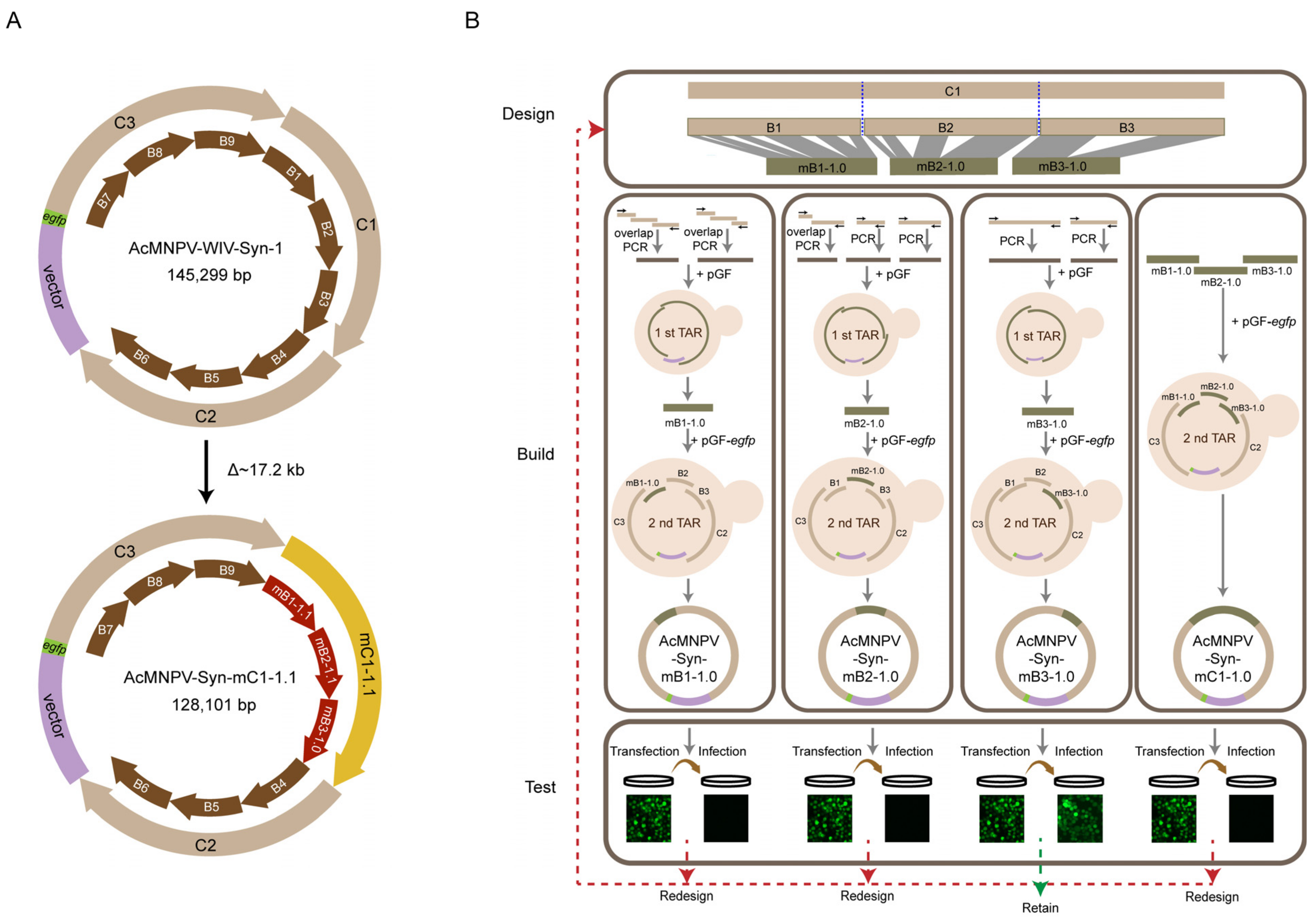
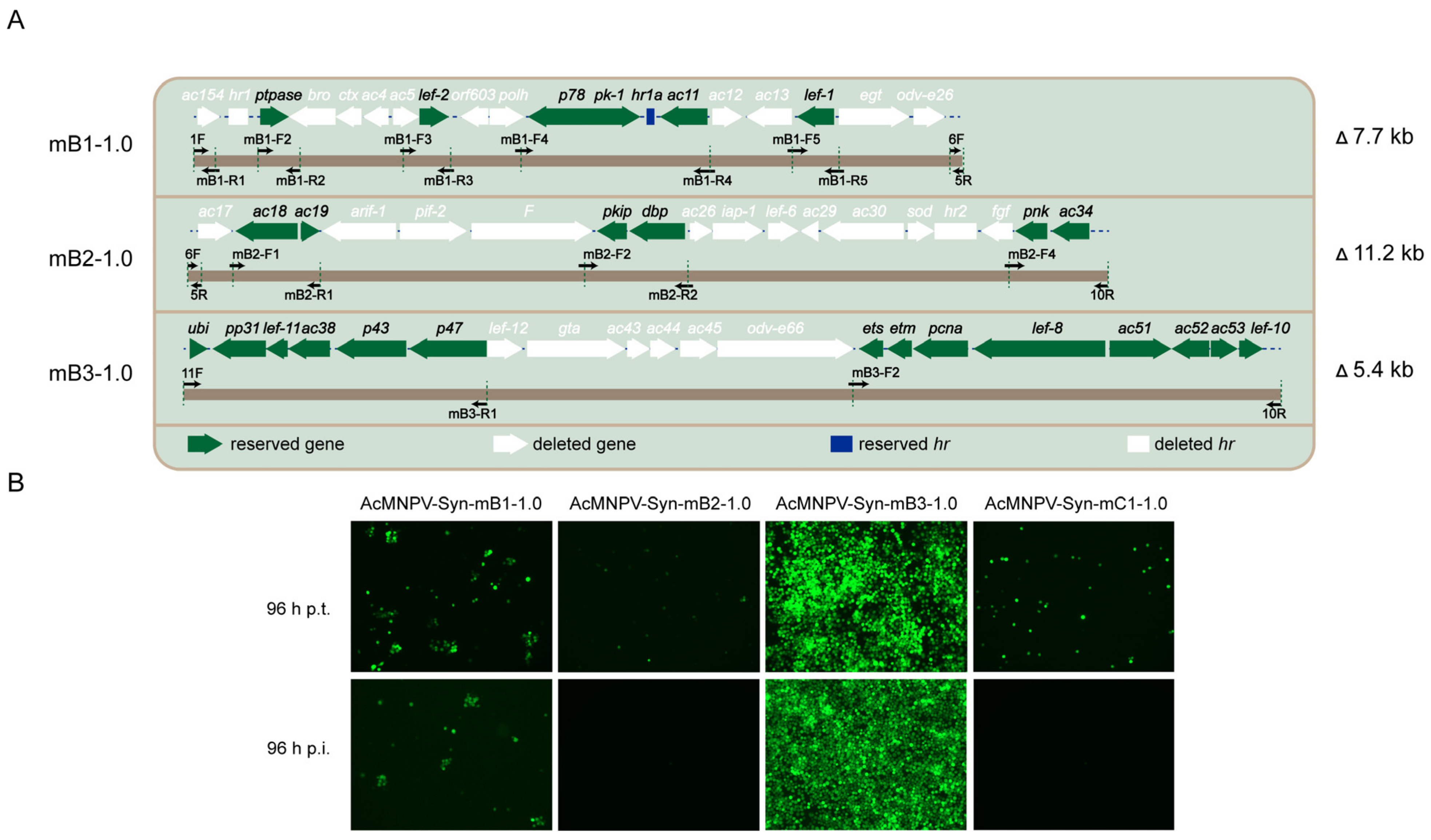
3.1. Synthesis and Characterization of the Version 1.0 of Modified Genomes
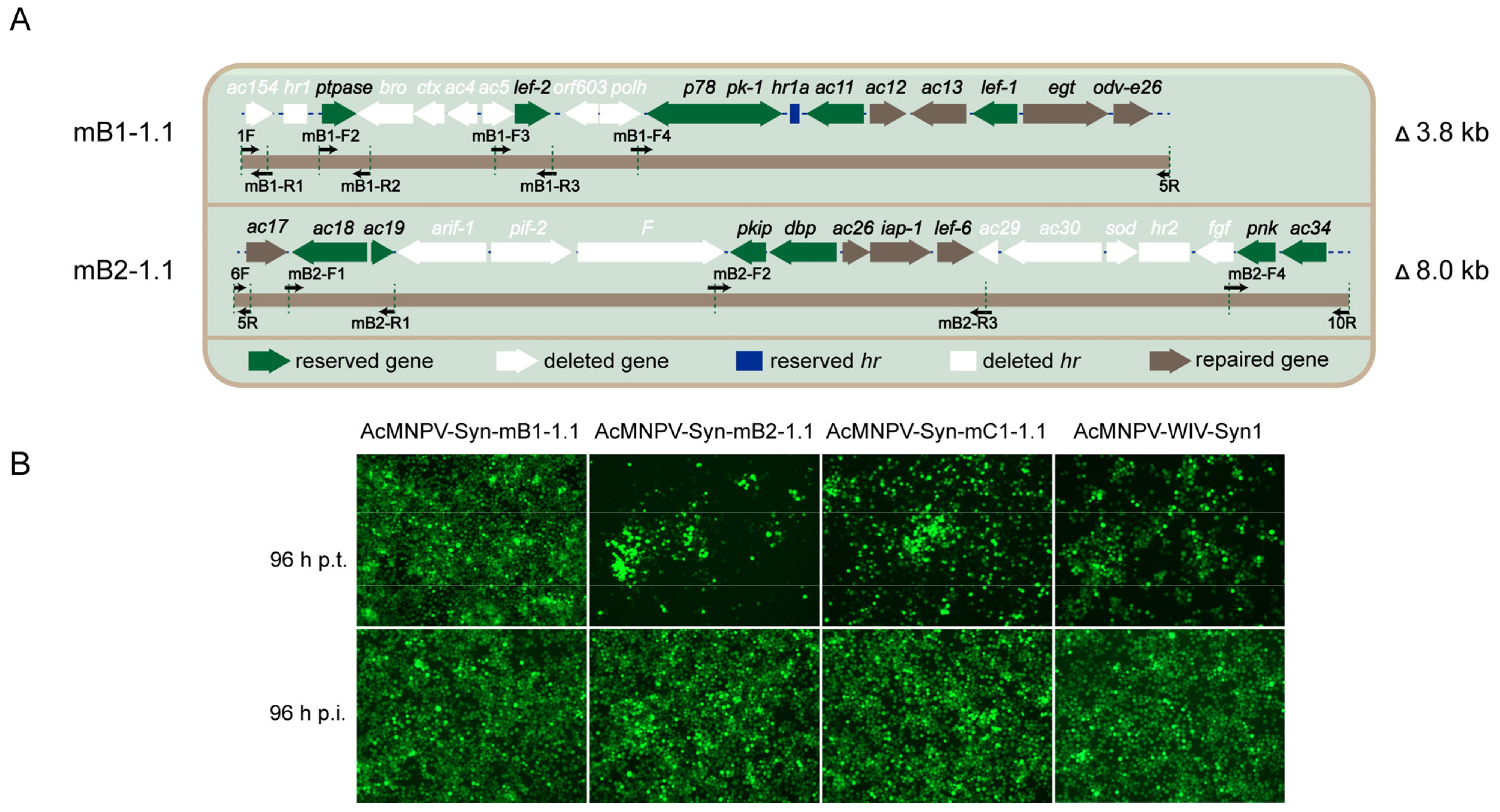
3.2. Redesign and Synthesis of AcMNPV-Syn-mB1-1.1, AcMNPV-Syn-mB2-1.1, and AcMNPV-Syn-mC1-1.1
3.3. Restriction Enzyme Analysis and Genome Sequencing of AcMNPV-Syn-mC1-1.1
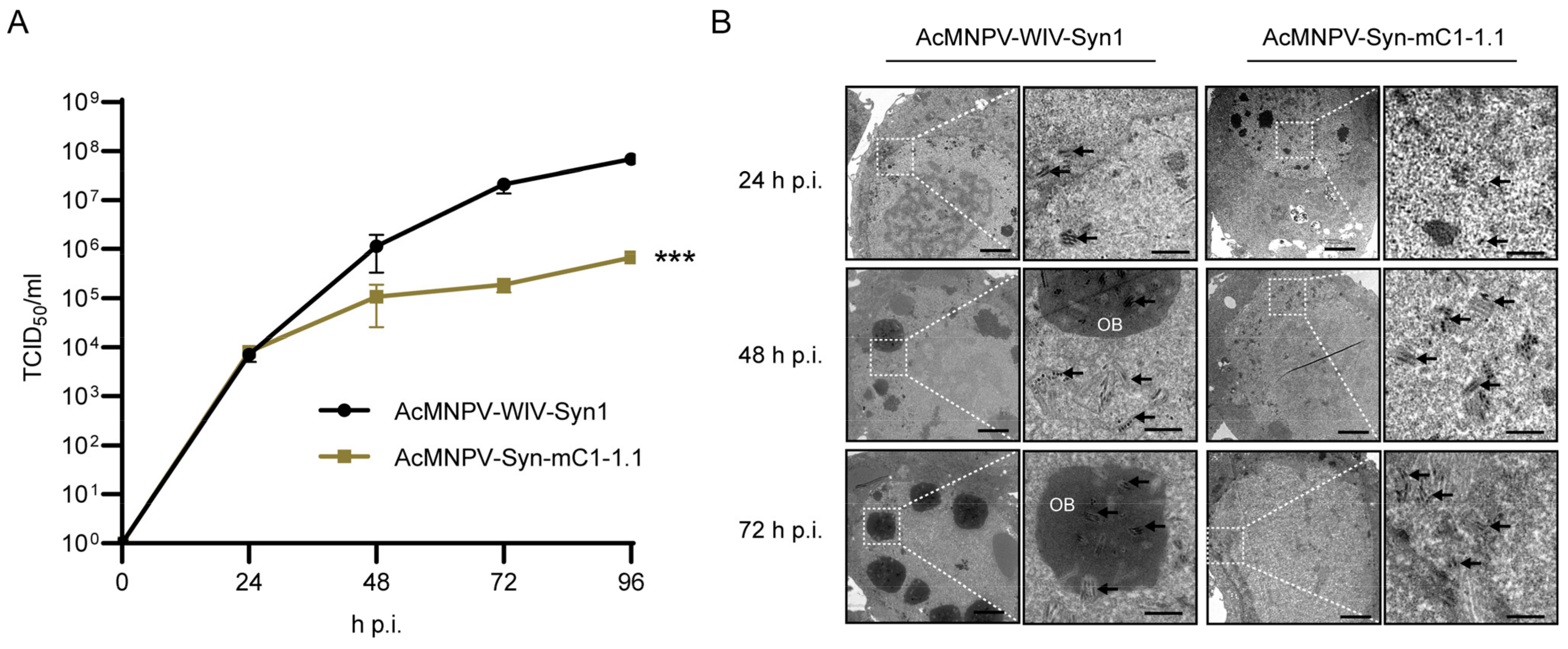
3.4. One-Step Growth Curve Analysis and Electron Microscopy of AcMNPV-Syn-mC1-1.1 in Comparison with the Parental Virus
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X. History and current status of development and use of viral insecticides in China. Viruses 2015, 7, 306–319. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.E.; Summers, M.D.; Fraser, M.J. Production of human beta interferon in insect cells infected with a baculovirus expression vector. Mol. Cell. Biol. 1983, 3, 2156–2165. [Google Scholar] [CrossRef] [PubMed]
- van Oers, M.M.; Pijlman, G.P.; Vlak, J.M. Thirty years of baculovirus–insect cell protein expression: From dark horse to mainstream technology. J. Gen. Virol. 2015, 96, 6–23. [Google Scholar] [CrossRef] [PubMed]
- Haddley, K. Alipogene tiparvovec for the treatment of lipoprotein lipase deficiency. Drugs Today 2013, 49, 161–170. [Google Scholar] [CrossRef]
- Blissard, G.W.; Theilmann, D.A. Baculovirus entry and egress from insect cells. Annu. Rev. Virol. 2018, 5, 113–139. [Google Scholar] [CrossRef]
- Rohrmann, G.F. Baculovirus Molecular Biology, 4th ed.; National Center for Biotechnology Information: Bethesda, MD, USA, 2019; p. 2. [Google Scholar]
- Wang, X.; Shang, Y.; Chen, C.; Liu, S.; Chang, M.; Zhang, N.; Hu, H.; Zhang, F.; Zhang, T.; Wang, Z.; et al. Baculovirus per os infectivity factor complex: Components and assembly. J. Virol. 2019, 93, e02053-18. [Google Scholar] [CrossRef] [Green Version]
- Slack, J.M.; Kuzio, J.; Faulkner, P. Characterization of v-cath, a cathepsin L-like proteinase expressed by the baculovirus Autographa californica multiple nuclear polyhedrosis virus. J. Gen. Virol. 1995, 76, 1091–1098. [Google Scholar] [CrossRef]
- Mason, C.S.; Lamers, M.B.A.C.; Henderson, I.M.J.; Monk, T.; Williams, D.H. Baculoviral expression and characterization of rodent cathepsin S. Protein Express Purif. 2001, 23, 45–54. [Google Scholar] [CrossRef]
- Maghodia, A.B.; Jarvis, D.L.; Geisler, C. Complete genome sequence of the Autographa californica multiple nucleopolyhedrovirus strain E2. Genome A 2014, 2, e01202-14. [Google Scholar] [CrossRef]
- Shang, Y.; Wang, M.; Xiao, G.; Wang, X.; Hou, D.; Pan, K.; Liu, S.; Li, J.; Wang, J.; Arif, B.M.; et al. Construction and rescue of a functional synthetic baculovirus. ACS Synth. Biol. 2017, 6, 1393–1402. [Google Scholar] [CrossRef]
- Gibson, D.G.; Benders, G.A.; Andrews-Pfannkoch, C.; Denisova, E.A.; Baden-Tillson, H.; Zaveri, J.; Stocwell, T.B.; Brownley, A.; Thomas, D.W.; Algire, M.A.; et al. Complete chemical synthesis, assembly, and cloning of a mycoplasma genitalium genome. Science 2008, 319, 1215. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Duan, X.; Hu, H.; Shang, Y.; Hu, Y.; Deng, F.; Wang, H.; Wang, M.; Hu, Z. Systematic analysis of 42 Autographa californica multiple nucleopolyhedrovirus genes identifies an additional six genes involved in the production of infectious budded virus. Virol. Sin. 2021, 36, 762–773. [Google Scholar] [CrossRef]
- McCarroll, L.; King, L.A. Stable insect cell cultures for recombinant protein production. Curr. Opin. Biotechnol. 1997, 8, 590–594. [Google Scholar] [CrossRef]
- Hou, Z.; Zhou, Z.; Wang, Z.; Xiao, G. Assembly of long DNA sequences using a new synthetic Escherichia coli-yeast shuttle vector. Virol. Sin. 2016, 31, 160–167. [Google Scholar] [CrossRef]
- Chen, Y.; Zhong, S.; Fei, Z.; Hashimoto, Y.; Xiang, J.Z.; Zhang, S.; Blissarda, G.W. The transcriptome of the baculovirus autographa californica multiple nucleopolyhedrovirus in trichoplusia ni cells. J. Virol. 2013, 87, 6391–6405. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Pan, K.; Shang, Y.; Guo, Y.; Xiao, H.; Deng, F.; Wang, M.; Hu, Z. Multi-loci manipulation of baculovirus genome reveals the pivotal role of homologous regions in viral DNA replication, progeny production, and enhancing transcription. ACS Synth. Biol. 2022, 11, 144–153. [Google Scholar] [CrossRef]
- Wang, M.; Tuladhar, E.; Shen, S.; Wang, H.; Oers, M.M.; Westenberg, M. Specificity of baculovirus p6.9 basic DNA-binding proteins and critical role of the C terminus in virion formation. J. Virol. 2010, 84, 8821–8828. [Google Scholar] [CrossRef] [Green Version]
- Kokusho, R.; Koh, Y.; Fujimoto, M.; Shimada, T.; Katsuma, S. Bombyx mori nucleopolyhedrovirus BM5 protein regulates progeny virus production and viral gene expression. Virology 2016, 498, 240–249. [Google Scholar] [CrossRef]
- Ono, C.; Kamagata, T.; Taka, H.; Sahara, K.; Asano, S. Phenotypic grouping of 141 BmNPVs lacking viral gene sequences. Virus Res. 2016, 165, 197–206. [Google Scholar] [CrossRef]
- Nie, Y.; Theilmann, D.A. Deletion of AcMNPV AC16 and AC17 results in delayed viral gene expression in budded virus infected cells but not transfected cells. Virology 2010, 404, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Lin, G.; Blissard, G.W. Analysis of an Autographa californica multicapsid nucleopolyhedrovirus lef-6-null virus: LEF-6 is not essential for viral replication but appears to accelerate late gene transcription. J. Virol 2002, 76, 5503–5514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, D.R.; Miller, L.K. A Baculovirus blocks insect molting by producing Ecdysteroid UDP-Glucosyl transferase. Science 1989, 245, 1110–1112. [Google Scholar] [CrossRef] [PubMed]
- McLachlin, J.R.; Escobar, J.C.; Harrelson, J.A.; Clem, R.J.; Miller, K.K. Deletions in the Ac-iap1 gene of the baculovirus AcMNPV occur spontaneously during serial passage and confer a cell line-specific replication ad-vantage. Virus Res. 2001, 81, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Carstens, E.B.; Wu, Y. No single homologous repeat region is essential for DNA replication of the baculovirus Autographa californica multiple nucleopolyhedrovirus. J. Gen. Virol. 2007, 88, 114–122. [Google Scholar] [CrossRef]
- Bideshi, D.K.; Renault, S.; Stasiak, K.; Federici, B.A.; Bigot, Y. Phylogenetic analysis and possible function of bro-like genes, a multigene family widespread among large double-stranded DNA viruses of invertebrates and bacteria. J. Gen. Virol. 2003, 84, 2531–2544. [Google Scholar] [CrossRef]
- Cao, Q.; Zhu, S.Y.; Wu, Y.; Liu, Y.; Zhu, J.; Wang, W. The effect of a small conotoxin-like ctx gene from Autographa californica Nuclear Polyhedrosis Virus (AcMNPV) on insect hemolymph melanization. Pol. J. Microbiol. 2012, 61, 183–189. [Google Scholar] [CrossRef]
- Eldridge, R.; Li, Y.; Miller, L. Characterization of a baculovirus gene encoding a small conotoxinlike polypeptide. J. Virol. 1992, 66, 6563–6571. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, T.; Rowe, A.R.; Volkman, L.E. Identification of six Autographa californica multicapsid nucleopolyhedrovirus early genes that mediate nuclear localization of G-actin. J. Virol. 2002, 76, 12281–12289. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, C.; Zhang, N.; Li, J.; Deng, F.; Wang, H.; Vlak, J.M.; Hu, Z.; Wang, M. The group I alphabaculovirus-specific protein, AC5, is a novel component of the occlusion body but is not associated with ODVs or the PIF complex. J. Gen. Virol. 2018, 99, 585–595. [Google Scholar] [CrossRef]
- Gearing, K.L.; Possee, R.D. Functional analysis of a 603 nucleotide open reading frame upstream of the polyhedrin gene of Autographa californica nuclear polyhedrosis virus. J. Gen. Virol. 1990, 71, 251–262. [Google Scholar] [CrossRef]
- Ji, X.; Sutton, G.; Evans, G.; Axford, D.; Owen, R.; Stuart, D.I. How baculovirus polyhedra fit square pegs into round holes to robustly package viruses. EMBO J. 2010, 29, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Rohrmann, G. Polyhedrin structure. J. Gen. Virol 1986, 67, 1499–1513. [Google Scholar] [CrossRef]
- Kokusho, R.; Kawamoto, M.; Koyano, Y.; Sugano, S.; Suzuki, Y.; Shimada, T.; Katsuma, S. Bombyx mori nucleopolyhedrovirus actin rearrangement-inducing factor 1 enhances systemic infection in B. mori larvae. J. Gen. Virol. 2015, 96, 1938–1946. [Google Scholar] [CrossRef]
- Roncarati, R.; Knebel-Mörsdorf, D. Identification of the early actin-rearrangement-inducing factor gene, arif-1, from Autographa californica multicapsid nuclear polyhedrosis virus. J. Virol. 1997, 71, 7933–7941. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, T.; Washburn, J.O.; Sitapara, R.; Sid, E.; Volkman, L.E. Specific binding of Autographa californica M nucleopolyhedrovirus occlusion-derived virus to midgut cells of heliothis virescens larvae is mediated by products of pif genes Ac119 and Ac022 but not by Ac115. J. Virol. 2005, 79, 15258–15264. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Tan, Y.; Yin, F.; Deng, F.; Vlak, J.M.; Hu, Z.; Wang, H. The F-like protein Ac23 enhances the infectivity of the budded virus of gp64-null AcMNPV pseudotyped with baculovirus envelope fusion protein F. J. Virol. 2008, 82, 9800–9804. [Google Scholar] [CrossRef] [Green Version]
- Lung, O.Y.; Cruz-Alvarez, M.; Blissard, G.W. Ac23, an envelope fusion protein homolog in the baculovirus Autographa californica multicapsid nucleopolyhedrovirus, is a viral pathogenicity factor. J. Virol. 2003, 77, 328–339. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q. Sequence analysis and function of baculovirus ac30 gene. J. Huazhong Agric. Univ. 2015, 34, 13–19. [Google Scholar]
- Tomalski, M.D.; Eldridge, R.; Miller, L.K. A baculovirus homolog of a Cu/Zn superoxide dismutase gene. Virology 1991, 1, 149–161. [Google Scholar] [CrossRef]
- Detvisitsakun, C.; Berretta, M.F.; Lehiy, C.; Passarelli, A.L. Stimulation of cell motility by a viral fibroblast growth factor homolog: Proposal for a role in viral pathogenesis. Virology 2005, 336, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Harwood, S.H.; Rohrmann, G.F. Identification of additional genes that influence baculovirus late gene expression. Virology 1999, 255, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarino, L.A.; Mistretta, T.A.; Dong, W. Baculovirus lef-12 is not required for viral replication. J. Virol. 2002, 76, 12032–12043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, X.Y.; Choi, J.Y.; Wang, Y.; Roh, J.Y.; Lee, J.H.; Liu, Q.; Park, J.B.; Kim, J.S.; Kim, W.; Je, Y.H. Functional characterization of Autographa californica multiple nucleopolyhedrovirus ORF43 and phenotypic changes of ORF43-knockout mutant. J. Microbiol. 2013, 51, 515–521. [Google Scholar] [CrossRef]
- Xiang, X.; Chen, L.; Hu, X.; Yu, S.; Yang, R.; Wu, X. Autographa californica multiple nucleopolyhedrovirus odve66 is an essential gene required for oral infectivity. Virus. Res. 2011, 158, 72–78. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) * | Applied for |
|---|---|---|
| 1F | CGACAGCTGATAGAAGAAAATACTCGTCTC | mB1-1.0 and mB1-1.1 |
| mB1-R1 | GCTTTACGAGTAGAATTCGGGGTCAACATC GATAGTGTCATACG | mB1-1.0 and mB1-1.1 |
| mB1-F2 | GAATTCTACTCGTAAAGCGAGTTGAAGGATCA | mB1-1.0 and mB1-1.1 |
| mB1-R2 | GGCCAAAAGACGTACGTGGAAAAC | mB1-1.0 and mB1-1.1 |
| mB1-F3 | GTTTTCCACGTACGTCTTTTGGCCCCATCCA ATCGACCGTTAGTCG | mB1-1.0 and mB1-1.1 |
| mB1-R3 | GGTGCGTCTGGTGCAAACTCACAAATGTTC TTGTTGCGTTTGGTT | mB1-1.0 and mB1-1.1 |
| mB1-F4 | AACCAAACGCAACAAGAACATTTGTGAGTT TGCACCAGACGCACC | mB1-1.0 and mB1-1.1 |
| mB1-R4 | GGGACGGCGATCAGCACGCGTGCCTGTAGA TCAGT | mB1-1.0 |
| mB1-F5 | GCTGATCGCCGTCCCTGTAGCATTTTGCGTT CGTGTCG | mB1-1.0 |
| mB1-R5 | CTATCAAAGCCATCTCCGCATTTACAGCAGT AAGCGTAGA | mB1-1.0 |
| 5R | CCGTCAAACGTTACATGCTTTTCG | mB1-1.0 and mB2-1.0 |
| 6F | AGATGGCTTTGATAGCGCTTATATTCAG | mB1-1.0 and mB2-1.0 |
| mB2-F1 | CGAAAAGCATGTAACGTTTGACGGTTATCG CGCAGGCGATCTTC | mB2-1.0 |
| mB2-R1 | CCGCCCATGTGTTTTAGCGAAGAGT | mB2-1.0 |
| mB2-F2 | AAAACACATGGGCGGAGCGATGATATGCCA AAACCATTGTACC | mB2-1.0 and mB2-1.1 |
| mB2-R2 | AAACTTTCTCAACTACGGGCTTGAAAGGGG CGCATTTGGAATGA | mB2-1.0 |
| mB2-R3 | GACAAAACTTTCTCAACTACGGAATAGACAAT | mB2-1.1 |
| mB2-F4 | CCGTAGTTGAGAAAGTTTTGTCCCACAGCAAA CTGGCGCTTTTATA | mB2-1.0 and mB2-1.1 |
| 10R | CACACCACGAATTATTTCCCTTCAAC | mB2-1.0 |
| 11F | AGTGGCCCGGTGTTATATTAAGTCG | mB3-1.0 |
| mB3-R1 | TGTTGCGTGCAATAGCCCTGC | mB3-1.0 |
| mB3-F2 | GCAGGGCTATTGCACGCAACACAAAAGACT GACTGTTAACA CAAAAGACTGA | mB3-1.0 |
| 16R | AAACGCTCATGTTGTGTTCGCC | mB3-1.0 |
| Name | Reduced Size (kb) | No. of Deleted Gene | Modified Position | BV Titre a | |
|---|---|---|---|---|---|
| (TCID50/mL) | p Value | ||||
| AcMNPV-Syn1 | 0 | 0 | none | 6.22 × 107 | - |
| Syn-mB1-1.0 | 7.7 | 12 | B1 | - | - |
| Syn-mB1-1.1 | 3.8 | 8 | B1 | 6.39 × 107 | 0.9998 (ns) |
| Syn-mB2-1.0 | 11.2 | 12 | B2 | - | - |
| Syn-mB2-1.1 | 8 | 8 | B2 | 4.58 × 106 | 0.0093 (**) |
| Syn-mB3-1.0 | 5.4 | 6 | B3 | 5.65 × 107 | 0.9839 (ns) |
| Syn-mC1-1.0 | 24.2 | 30 | B1, B2, and B3 | - | - |
| Syn-mC1-1.1 | 17.2 | 22 | B1, B2, and B3 | 7.45 × 105 | 0.0062 (**) |
| Number | Deleted Gene | Protein | Position | Function | Impact on BV Production |
|---|---|---|---|---|---|
| 1 | ac154 | B1 | Unverified | Non-essential [14] | |
| 2 | hr1 | B1 | Enhancer [18] | Non-essential [26] | |
| 3 | bro | Baculovirus repeated orf | B1 | Unverified | Non-essential [27] |
| 4 | ctl | Conotoxin-like protein | B1 | Inhibitor of melanisation [28] | Non-essential [29] |
| 5 | ac4 | B1 | Unverified | Non-essential [30] | |
| 6 | ac5 | B1 | Unverified | Non-essential [31] | |
| 7 | orf603 | Orf 603 | B1 | Unverified | Non-essential [32] |
| 8 | polh | Polyhedrin | B1 | Occlusion body matrix protein [33] | Non-essential [34] |
| 9 | arif1 | Arif-1 | B2 | Induction of actin rearrangement [35] | Non-essential [36] |
| 10 | pif2 | PIF-2 | B2 | Oral infection [37] | Non-essential [37] |
| 11 | F | Fusion protein | B2 | Associated with cell binding [38] | Non-essential [39] |
| 12 | ac29 | B2 | Unverified | Non-essential [14] | |
| 13 | ac30 | B2 | Unverified | Non-essential [40] | |
| 14 | sod | Super oxide dismutase | B2 | Super oxide dismutase [41] | Non-essential [41] |
| 15 | hr2 | B2 | Enhancer [18] | Non-essential [26] | |
| 16 | fgf | Fibroblast growth factor | B2 | Stimulation of insect cell motility [42] | Non-essential [42] |
| 17 | lef-12 | Lef-12 | B3 | Transcription [43] | Non-essential [44] |
| 18 | gta | Global transactivator-like protein | B3 | Unverified | Non-essential [14] |
| 19 | ac43 | B3 | Affect polyhedron [45] | Non-essential [45] | |
| 20 | ac44 | B3 | Unverified | Non-essential [14] | |
| 21 | ac45 | B3 | Unverified | Non-essential [14] | |
| 22 | odv-e66 | ODV-E66 | B3 | Oral infection [46] | Non-essential [46] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Hu, H.; Xiao, H.; Deng, F.; Li, J.; Wang, M.; Hu, Z. Successful Rescue of Synthetic AcMNPV with a ~17 kb Deletion in the C1 Region of the Genome. Viruses 2022, 14, 2780. https://doi.org/10.3390/v14122780
Guo Y, Hu H, Xiao H, Deng F, Li J, Wang M, Hu Z. Successful Rescue of Synthetic AcMNPV with a ~17 kb Deletion in the C1 Region of the Genome. Viruses. 2022; 14(12):2780. https://doi.org/10.3390/v14122780
Chicago/Turabian StyleGuo, Yijia, Hengrui Hu, Han Xiao, Fei Deng, Jiang Li, Manli Wang, and Zhihong Hu. 2022. "Successful Rescue of Synthetic AcMNPV with a ~17 kb Deletion in the C1 Region of the Genome" Viruses 14, no. 12: 2780. https://doi.org/10.3390/v14122780
APA StyleGuo, Y., Hu, H., Xiao, H., Deng, F., Li, J., Wang, M., & Hu, Z. (2022). Successful Rescue of Synthetic AcMNPV with a ~17 kb Deletion in the C1 Region of the Genome. Viruses, 14(12), 2780. https://doi.org/10.3390/v14122780