
Divergent Rhabdovirus Discovered in a Patient with New-Onset Nodding Syndrome


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subject Identification, Clinical Assessment, and Sample Collection
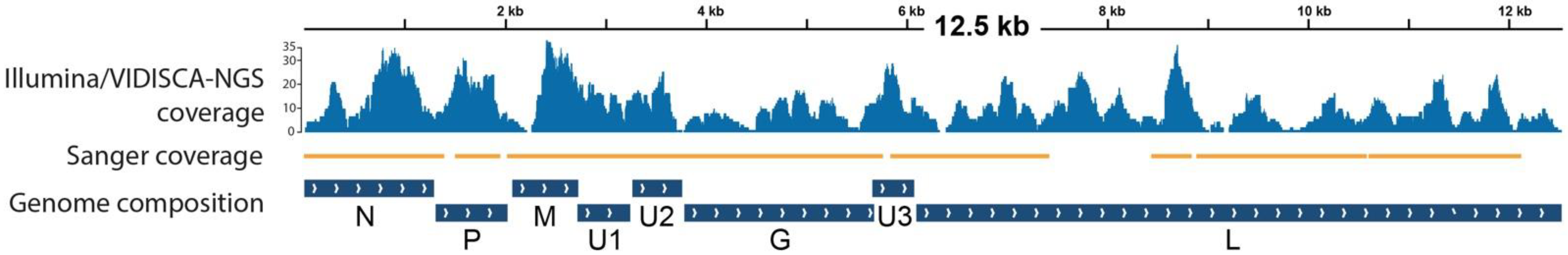
2.2. Virus Discovery
2.3. Genome Characterization and Analysis
2.4. Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
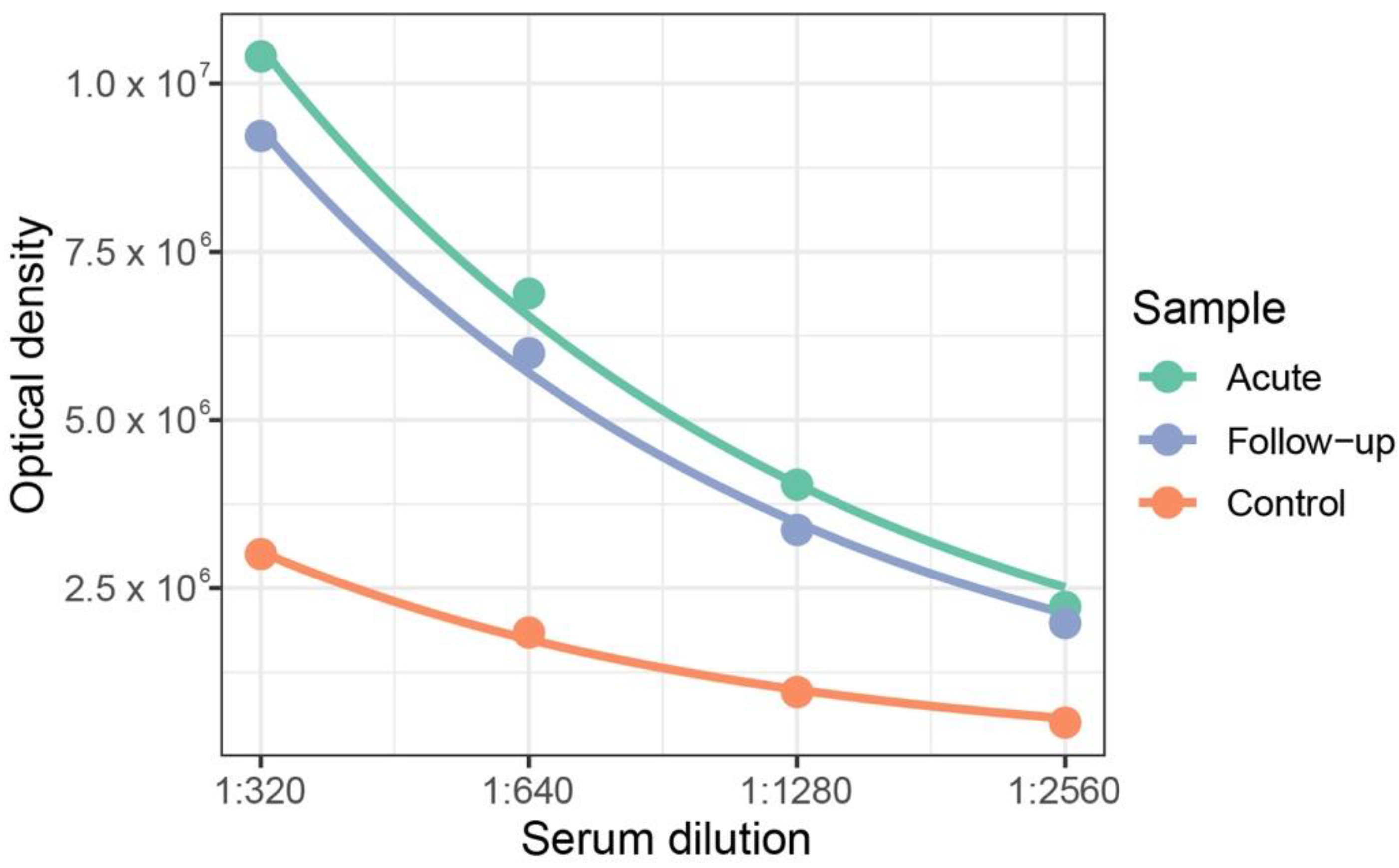
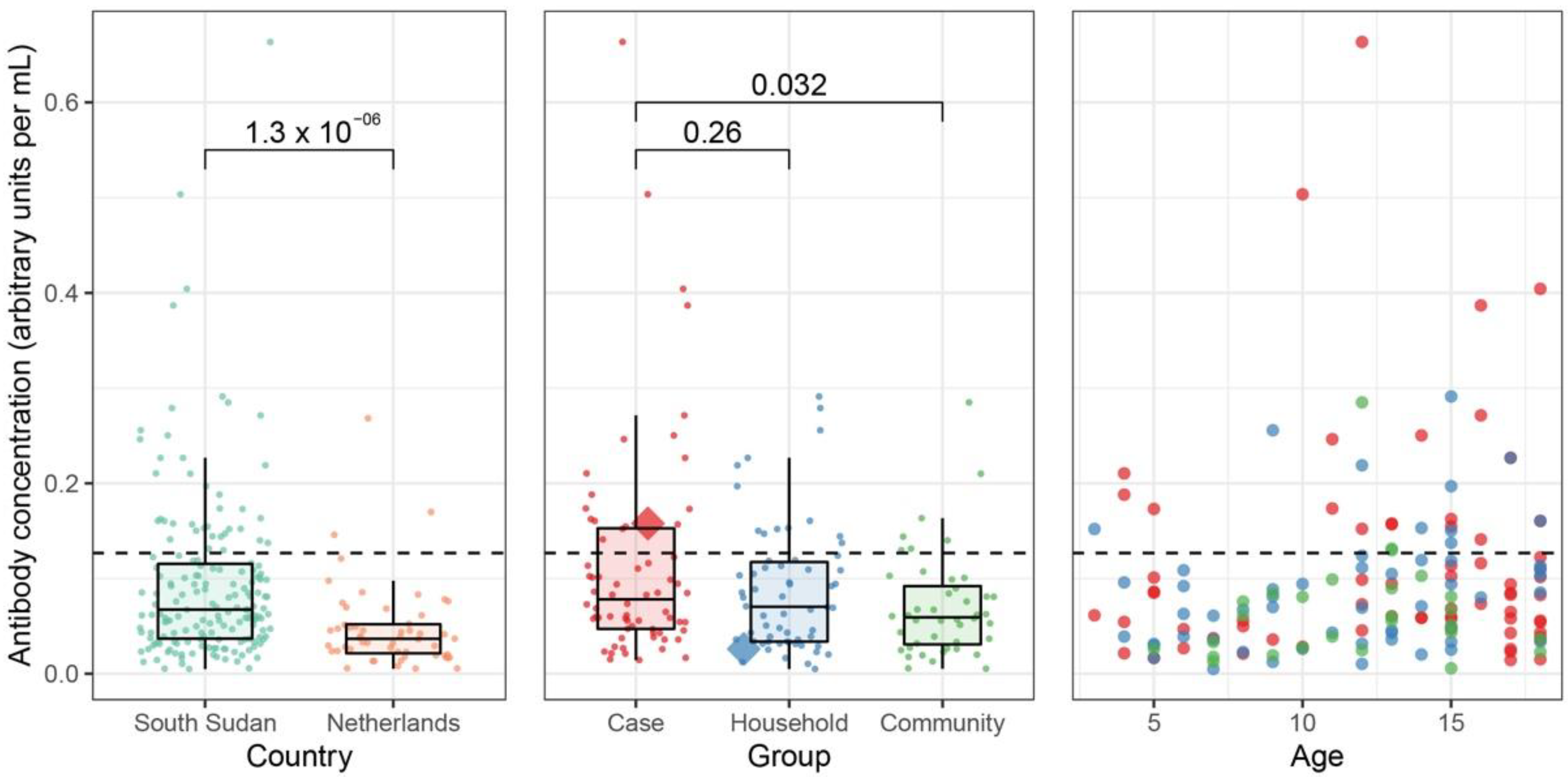
2.5. Serological Assessment
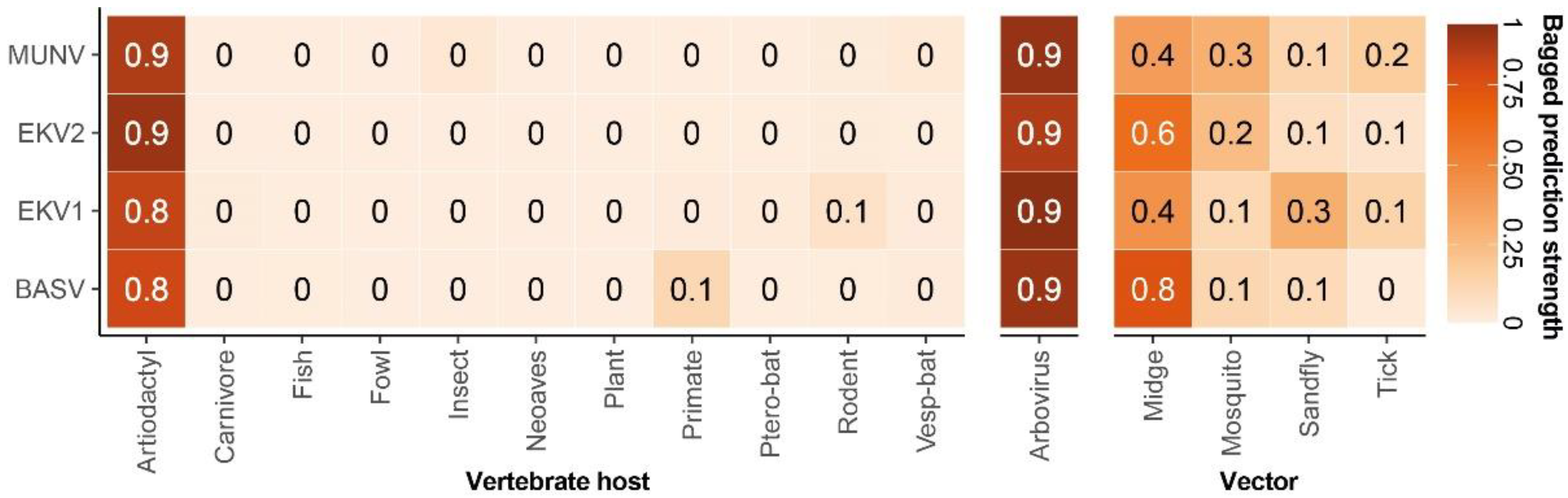
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olum, S.; Scolding, P.; Hardy, C.; Obol, J.; Scolding, N.J. Nodding syndrome: A concise review. Brain Commun. 2020, 2, fcaa037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colebunders, R.; Hendy, A.; Nanyunja, M.; Wamala, J.F.; van Oijen, M. Nodding syndrome—A new hypothesis and new direction for research. Int. J. Infect. Dis. 2014, 27, 74–77. [Google Scholar] [CrossRef] [Green Version]
- Foltz, J.L.; Makumbi, I.; Sejvar, J.J.; Malimbo, M.; Ndyomugyenyi, R.; Atai-Omoruto, A.D.; Alexander, L.N.; Abang, B.; Melstrom, P.; Kakooza, A.M.; et al. An Epidemiologic Investigation of Potential Risk Factors for Nodding Syndrome in Kitgum District, Uganda. PLoS ONE 2013, 8, e66419. [Google Scholar] [CrossRef] [Green Version]
- van der Hoek, L.; Pyrc, K.; Jebbink, M.F.; Vermeulen-Oost, W.; Berkhout, R.J.M.; Wolthers, K.C.; Wertheim-van Dillen, P.M.E.; Kaandorp, J.; Spaargaren, J.; Berkhout, B. Identification of a new human coronavirus. Nat. Med. 2004, 10, 368–373. [Google Scholar] [CrossRef]
- Edridge, A.W.D.; Deijs, M.; Namazzi, R.; Cristella, C.; Jebbink, M.F.; Maurer, I.; Kootstra, N.A.; Buluma, L.R.; van Woensel, J.B.M.; de Jong, M.D.; et al. Novel Orthobunyavirus Identified in the Cerebrospinal Fluid of a Ugandan Child with Severe Encephalopathy. Clin. Infect. Dis. 2019, 68, 139–142. [Google Scholar] [CrossRef] [Green Version]
- Abd-Elfarag, G.; van Hensbroek, M.B. Nodding Syndrome: The Unresolved Mystery of a Pediatric Disease in Sub-Saharan Africa. Pediatr. Infect. Dis. J. 2019, 38, S67–S71. [Google Scholar] [CrossRef]
- Edridge, A.W.D.; Deijs, M.; Van Zeggeren, I.E.; Kinsella, C.M.; Jebbink, M.F.; Bakker, M.; Van de Beek, D.; Brouwer, M.C.; Van der Hoek, L. Viral metagenomics on cerebrospinal fluid. Genes 2019, 10, 332. [Google Scholar] [CrossRef] [Green Version]
- Boom, R.; Sol, C.J.A.; Salimans, M.M.M.; Jansen, C.L.; Wertheim-Van Dillen, P.M.E.; van der Noordaa, J. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Kinsella, C.M.; Deijs, M.; van der Hoek, L. Enhanced bioinformatic profiling of VIDISCA libraries for virus detection and discovery. Virus Res. 2018, 263, 21–26. [Google Scholar] [CrossRef]
- Flygare, S.; Simmon, K.; Miller, C.; Qiao, Y.; Kennedy, B.; Di Sera, T.; Graf, E.H.; Tardif, K.D.; Kapusta, A.; Rynearson, S.; et al. Taxonomer: An interactive metagenomics analysis portal for universal pathogen detection and host mRNA expression profiling. Genome Biol. 2016, 17, 111. [Google Scholar] [CrossRef] [Green Version]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Dijkman, R.; Jebbink, M.F.; El Idrissi, N.B.; Pyrc, K.; Müller, M.A.; Kuijpers, T.W.; Zaaijer, H.L.; Van Der Hoek, L. Human coronavirus NL63 and 229E seroconversion in children. J. Clin. Microbiol. 2008, 46, 2368–2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idro, R.; Musubire, K.; Byamah, M.; Namusoke, H.; Muron, J.; Abbo, C.; Oriyabuzu, R.; Ssekyewa, J.; Okot, C.; Mwaka, D.; et al. Proposed guidelines for the management of nodding syndrome. Afr. Health Sci. 2013, 13, 219–225. [Google Scholar] [CrossRef]
- Idro, R.; Opoka, R.O.; Aanyu, H.T.; Kakooza-Mwesige, A.; Piloya-Were, T.; Namusoke, H.; Musoke, S.B.; Nalugya, J.; Bangirana, P.; Mwaka, A.D.; et al. Nodding syndrome in Ugandan children—Clinical features, brain imaging and complications: A case series. BMJ Open 2013, 3, e002540. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, J.H.; Pān, H.; Chiu, C.Y.; Stremlau, M. Human Tibroviruses: Commensals or Lethal Pathogens? Viruses 2020, 12, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, P.J.; Blasdell, K.R.; Calisher, C.H.; Dietzgen, R.G.; Kondo, H.; Kurath, G.; Longdon, B.; Stone, D.M.; Tesh, R.B.; El Tordo, N.; et al. ICTV Virus Taxonomy Profile: Rhabdoviridae. J. Gen. Virol. 2018, 99, 447–448. [Google Scholar] [CrossRef] [PubMed]
- Babayan, S.A.; Orton, R.J.; Streicker, D.G. Predicting reservoir hosts and arthropod vectors from evolutionary signatures in RNA virus genomes. Science 2018, 362, 577–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, P.J.; Firth, C.; Widen, S.G.; Blasdell, K.R.; Guzman, H.; Wood, T.G.; Paradkar, P.N.; Holmes, E.C.; Tesh, R.B.; Vasilakis, N. Evolution of Genome Size and Complexity in the Rhabdoviridae. PLoS Pathog. 2015, 11, e1004664. [Google Scholar] [CrossRef] [Green Version]
- Stremlau, M.H.; Andersen, K.G.; Folarin, O.A.; Grove, J.N.; Odia, I.; Ehiane, P.E.; Omoniwa, O.; Omoregie, O.; Jiang, P.-P.; Yozwiak, N.L.; et al. Discovery of Novel Rhabdoviruses in the Blood of Healthy Individuals from West Africa. PLoS Negl. Trop. Dis. 2015, 9, e0003631. [Google Scholar] [CrossRef]
- Grard, G.; Fair, J.N.; Lee, D.; Slikas, E.; Steffen, I.; Muyembe, J.-J.; Sittler, T.; Veeraraghavan, N.; Ruby, J.G.; Wang, C.; et al. A Novel Rhabdovirus Associated with Acute Hemorrhagic Fever in Central Africa. PLoS Pathog. 2012, 8, e1002924. [Google Scholar] [CrossRef] [PubMed]
- Bas-Congo Virus—Not an Established Pathogen Science. Available online: https://science.sciencemag.org/content/bas-congo-virus-not-established-pathogen (accessed on 30 August 2021).
- Carroll, D.; Daszak, P.; Wolfe, N.D.; Gao, G.F.; Morel, C.M.; Morzaria, S.; Pablos-Méndez, A.; Tomori, O.; Mazet, J.A.K. The Global Virome Project. Science 2018, 359, 872–874. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.J. From PREDICT to prevention, one pandemic later. Lancet Microbe 2020, 1, e6–e7. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORF/Protein | Virus | ORF nt Location | Aa Length | Signal Peptide (aa Cleavage Site) | Transmembrane Domain (aa Location) | Glycosylation Sites | |
|---|---|---|---|---|---|---|---|
| N | O | ||||||
| 1/N | MUNV | 1–1284 | 427 | None | None | 1 | 4 |
| EKV2 | 48–1331 | 427 | None | None | 4 | 3 | |
| 2/P | MUNV | 1309–2013 | 234 | None | None | 2 | 11 |
| EKV2 | 1396–2049 | 217 | None | None | 4 | 17 | |
| 3/M | MUNV | 2058–2705 | 215 | None | None | 0 | 7 |
| EKV2 | 2024–2713 | 229 | None | None | 2 | 8 | |
| 4/U1 | MUNV | 2702–3226 | 174 | None | None | 0 | 0 |
| EKV2 | 2710–3237 | 175 | None | None | 0 | 2 | |
| 5/U2 | MUNV | 3245–3736 | 163 | None | None | 2 | 0 |
| EKV2 | 3234–3740 | 168 | None | None | 1 | 2 | |
| 6/G | MUNV | 3761–5635 | 624 | 1 (18–19) | 1 (570–592) | 7 | 6 |
| EKV2 | 3950–5842 | 630 | 1 (23–24) | 1 (577–599) | 4 | 3 | |
| 7/U3 | MUNV | 5613–6026 | 137 | None | 1 (36–62) | 0 | 3 |
| EKV2 | 5844–6221 | 125 | None | 1 (23–40) | 1 | 4 | |
| 8/L | MUNV | 6049–12,429 | 2126 | None | None | 10 | 19 |
| EKV2 | 6263–12,625 | 2120 | None | None | 6 | 6 | |
| MUNV | EKV2 | EKV1 | BASV | TIBV | BAV | BHV | SWBV | CPV | |
|---|---|---|---|---|---|---|---|---|---|
| MUNV | ID | 0.568 | 0.414 | 0.45 | 0.415 | 0.416 | 0.415 | 0.411 | 0.415 |
| EKV2 | 0.570 | ID | 0.409 | 0.468 | 0.407 | 0.407 | 0.406 | 0.426 | 0.413 |
| EKV1 | 0.409 | 0.411 | ID | 0.423 | 0.414 | 0.414 | 0.414 | 0.421 | 0.438 |
| BASV | 0.432 | 0.43 | 0.38 | ID | 0.396 | 0.397 | 0.394 | 0.397 | 0.389 |
| TIBV | 0.402 | 0.393 | 0.473 | 0.375 | ID | 0.972 | 0.862 | 0.632 | 0.549 |
| BAV | 0.404 | 0.393 | 0.475 | 0.378 | 0.99 | ID | 0.857 | 0.628 | 0.549 |
| BHV | 0.395 | 0.388 | 0.466 | 0.366 | 0.943 | 0.946 | ID | 0.635 | 0.54 |
| SWBV | 0.388 | 0.379 | 0.454 | 0.368 | 0.752 | 0.759 | 0.747 | ID | 0.546 |
| CPV | 0.383 | 0.395 | 0.463 | 0.38 | 0.665 | 0.67 | 0.665 | 0.677 | ID |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edridge, A.W.D.; Abd-Elfarag, G.; Deijs, M.; Jebbink, M.F.; Boele van Hensbroek, M.; van der Hoek, L. Divergent Rhabdovirus Discovered in a Patient with New-Onset Nodding Syndrome. Viruses 2022, 14, 210. https://doi.org/10.3390/v14020210
Edridge AWD, Abd-Elfarag G, Deijs M, Jebbink MF, Boele van Hensbroek M, van der Hoek L. Divergent Rhabdovirus Discovered in a Patient with New-Onset Nodding Syndrome. Viruses. 2022; 14(2):210. https://doi.org/10.3390/v14020210
Chicago/Turabian StyleEdridge, Arthur W. D., Gasim Abd-Elfarag, Martin Deijs, Maarten F. Jebbink, Michael Boele van Hensbroek, and Lia van der Hoek. 2022. "Divergent Rhabdovirus Discovered in a Patient with New-Onset Nodding Syndrome" Viruses 14, no. 2: 210. https://doi.org/10.3390/v14020210
APA StyleEdridge, A. W. D., Abd-Elfarag, G., Deijs, M., Jebbink, M. F., Boele van Hensbroek, M., & van der Hoek, L. (2022). Divergent Rhabdovirus Discovered in a Patient with New-Onset Nodding Syndrome. Viruses, 14(2), 210. https://doi.org/10.3390/v14020210