
Influenza A Virus Causes Histopathological Changes and Impairment in Functional Activity of Blood Vessels in Different Vascular Beds

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Virus
2.3. Experimental Influenza Virus Infection
2.4. Tissue Lavage and Homogenization of Collected Tissues
2.5. Determination of Influenza Virus Infectivity Titer in Embryonated Chicken Eggs
2.6. Histological Analysis
2.7. Immunohistochemistry
2.8. Determination of Functional Activity of Blood Vessels
2.9. Statistical Analysis
3. Results
3.1. Body Weights; Clinical Observations
3.2. Influenza Virus Infectivity Titer in Pulmonary and Mesenteric Tissues
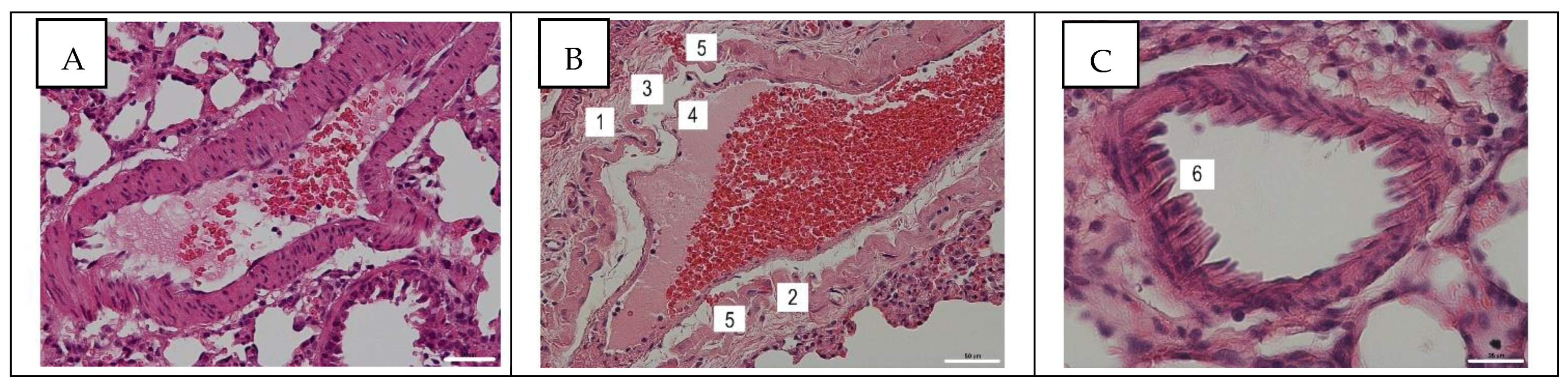
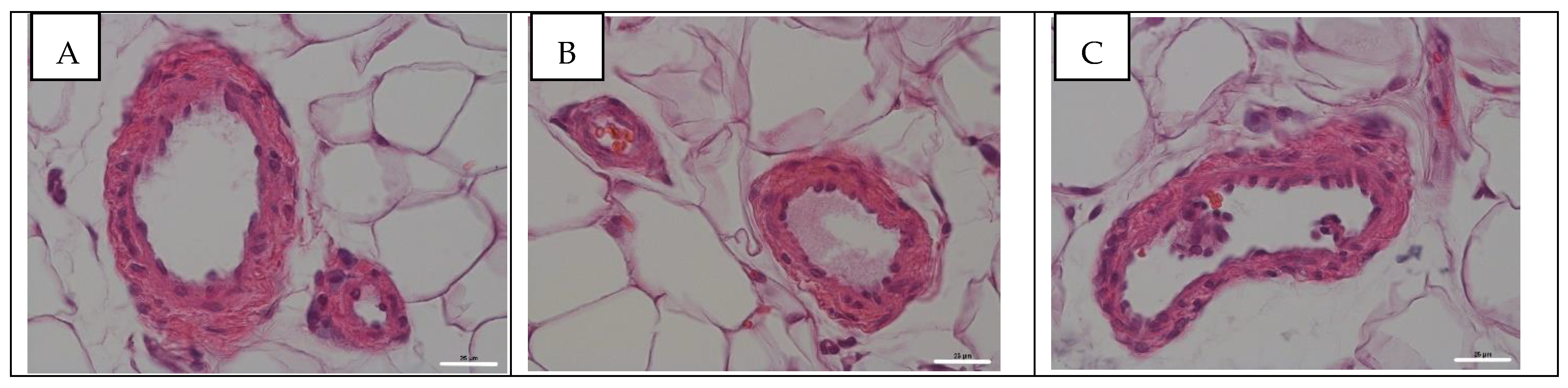
3.3. Histological Assay of Pulmonary and Mesentery Blood Vessels
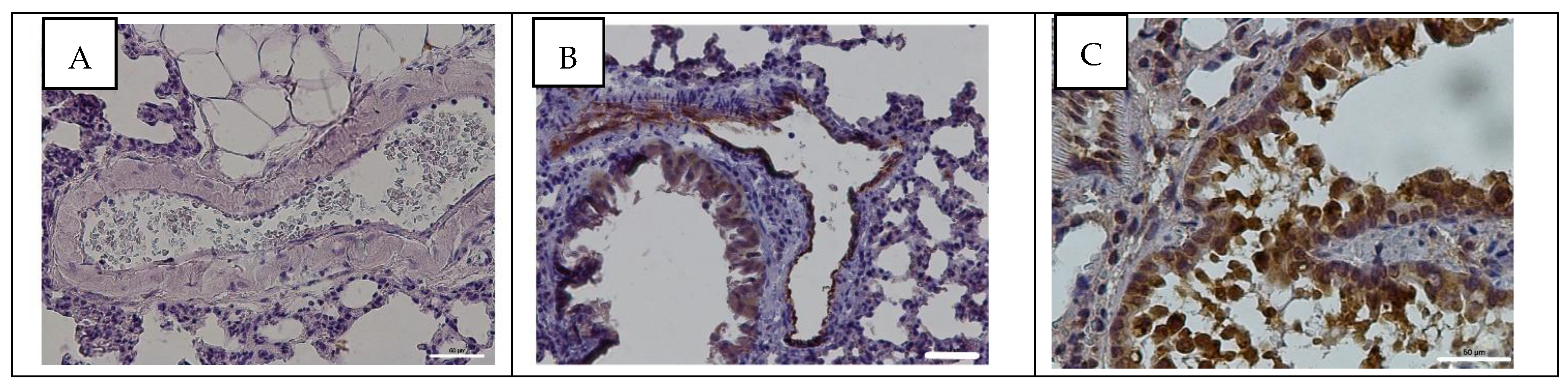
3.4. Detection of Influenza A Virus NP Antigen in Pulmonary and Mesenteric Tissues and Blood Vessels
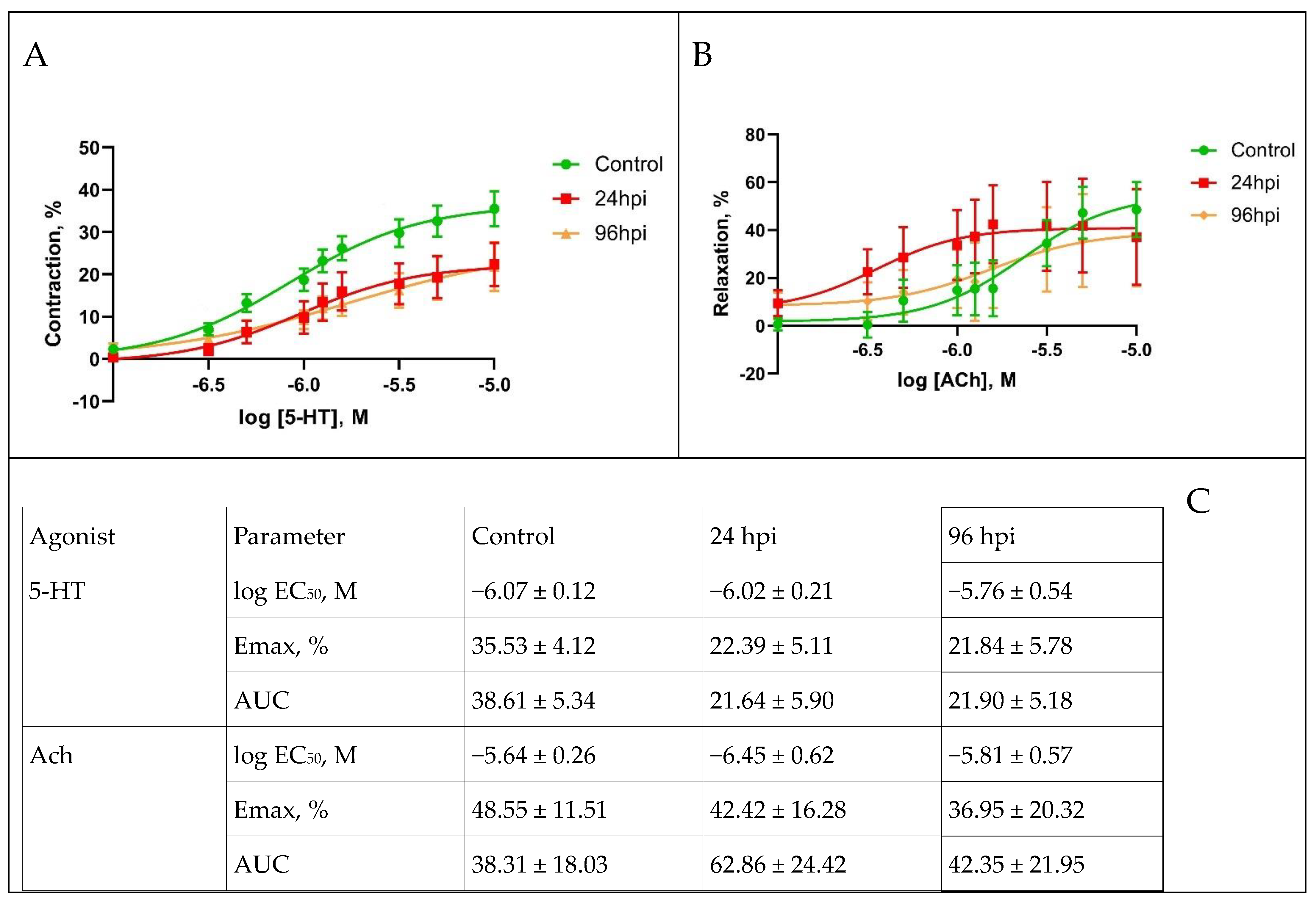
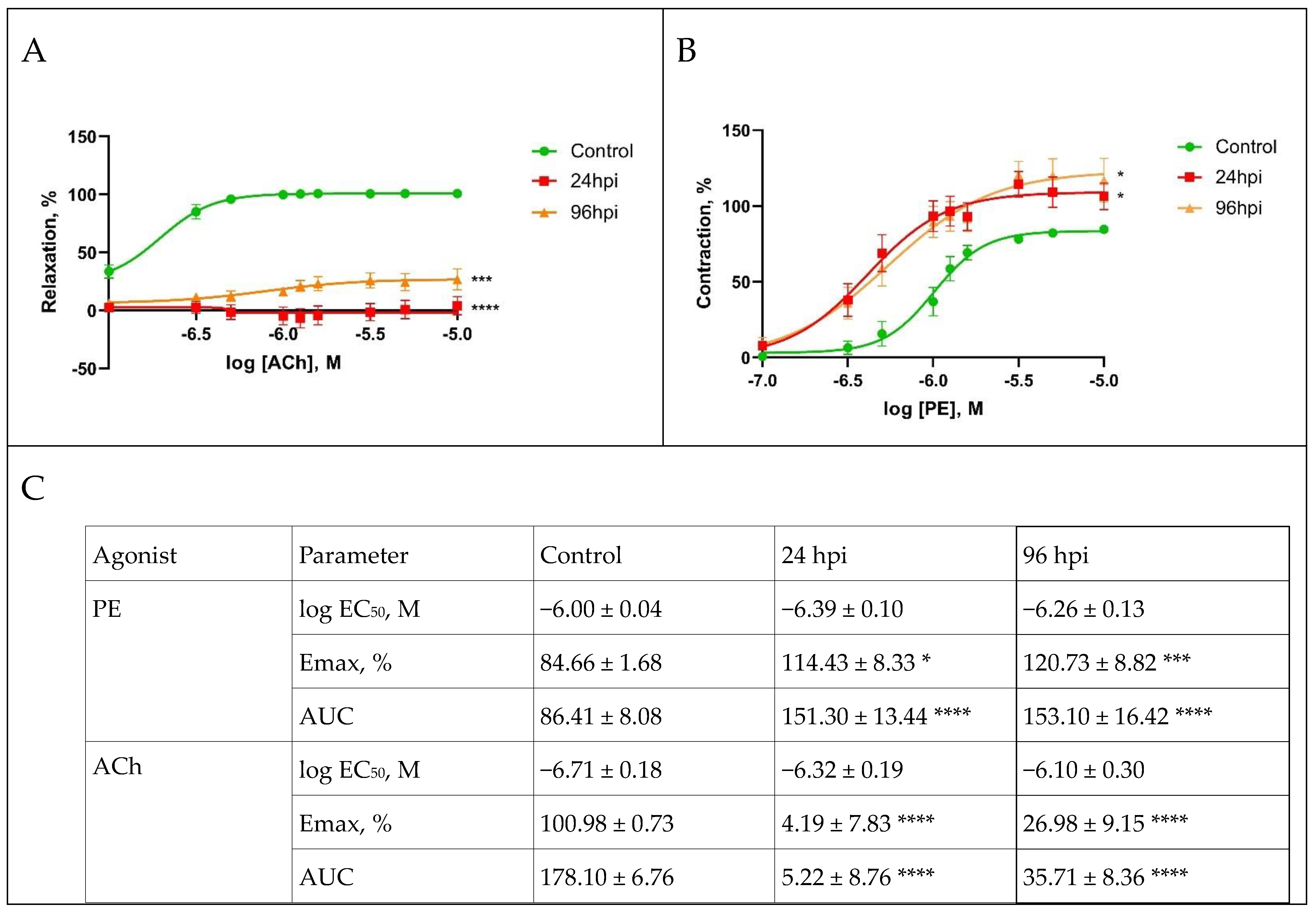
3.5. Functional Activity of Pulmonary and Mesenteric Arteries
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cárdenas, G.; Soto-Hernández, J.L.; Díaz-Alba, A.; Ugalde, Y.; Mérida-Puga, J.; Rosetti, M.; Sciutto, E. Neurological events related to influenza A (H1N1) pdm09. Influenza Other Respir. Viruses 2014, 8, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Rothberg, M.B.; Haessler, S.D.; Brown, R.B. Complications of Viral Influenza. Am. J. Med. 2008, 121, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Kwong, J.C.; Schwartz, K.L.; Campitelli, M.A.; Chung, H.; Crowcroft, N.S.; Karnauchow, T.; Katz, K.; Ko, D.T.; McGeer, A.J.; McNally, D.; et al. Acute Myocardial Infarction after Laboratory-Confirmed Influenza Infection. N. Engl. J. Med. 2018, 378, 345–453. [Google Scholar] [CrossRef] [PubMed]
- Lyon, E. Modern viewpoints on cardiovascular disturbances in influenza. Acta Med. Orient. 1952, 11, 25–34. [Google Scholar]
- Sergeev, N.V.; Leites, F.L. Some aspects of the study on circulatory disorders in influenza. Vestn. Akad. Meditsinskikh Nauk. SSSR 1962, 17, 15–22. [Google Scholar]
- Taubenberger, J.K.; Morens, D.M. The pathology of influenza virus infections. Annu. Rev. Pathol. 2008, 3, 499–522. [Google Scholar] [CrossRef] [PubMed]
- Lobo, S.M.; Watanabe, A.S.A.; Salomão, M.L.M.; Queiroz, F.; Gandolfi, J.V.; de Oliveira, N.E.; Covello, L.H.S.; Sacillotto, G.H.; de Godoy, L.G.; Simões, E.S.; et al. Excess mortality is associated with influenza A (H1N1) in patients with severe acute respiratory illness. J. Clin. Virol. 2019, 116, 62–68. [Google Scholar] [CrossRef]
- Fukunaga, S.; Ishida, C.; Nakaoka, A.; Ito, T. A case of acute kidney injury and disseminated intravascular coagulation associated with influenza B viral infection. CEN Case Rep. 2014, 4, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Yoshikawa, H.; Abe, Y.; Yamazaki, S.; Uehara, Y.; Abe, T. Renal involvement in children with influenza A virus infection. Pediatr. Nephrol. 2003, 18, 541–544. [Google Scholar] [CrossRef]
- Sulima, D.L.; Karev, V.E.; Zhdanov, K.V. Hypertoxic flu. J. Infectol. 2010, 2, 75–79. (In Russian) [Google Scholar]
- Esaulenko, E.V.; Stukolkin, K.O.; Dunaeva, N.V.; Grigor’eva, I.V.; Bondar, E.S.; Pozdniakova, M.G. Rare symptoms (diarrhea, meningism, hemor rhinic syndrome) in patients with different influenza virus subtypes. Vestnik NovSU. 2013, 71, 93–97. (In Russian) [Google Scholar]
- Uyeki, T.M. High-risk Groups for Influenza Complications. JAMA 2020, 324, 2334. [Google Scholar] [CrossRef] [PubMed]
- Tate, S.; Namkung, J.J.; Noymer, A. Did the 1918 influenza cause the twentieth century cardiovascular mortality epidemic in the United States? PeerJ 2016, 4, e2531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azambuja, M.I.R. Spanish flu and early 20th-century expansion of a coronary heart disease-prone subpopulation. Tex. Heart Inst. J. 2004, 31, 14–21. [Google Scholar]
- Ayora-Talavera, G. Sialic acid receptors: Focus on their role in influenza infection. J. Recept. Ligand Channel Res. 2018, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhilinskaya, I.N.; Azarenok, A.A.; Ilyinskaya, E.V.; Prochukhanova, A.R.; Vorob’ev, S.L.; Sorokin, E.V.; Taareva, T.R. Influenza virus reproduction in the endothelial cells of human blood vessels. Probl. Virol. 2012, 57, 20–23. (In Russian) [Google Scholar]
- Short, K.; Kroeze, E.J.B.V.; Reperant, L.; Richard, M.; Kuiken, T. Influenza virus and endothelial cells: A species specific relationship. Front. Microbiol. 2014, 5, 653. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, S.M.; Darwish, I.; Lee, W.L. Endothelial activation and dysfunction in the pathogenesis of influenza A virus infection. Virulence 2013, 4, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Marchenko, V.A.; Barashkova, S.V.; Zelinskaya, I.A.; Toropova, Y.G.; Ramsay, E.S.; Zhilinskaya, I.N. Modulation of endothelial factors activity in human endothelial cells in influenza A(H1N1)pdm09 virus infection. Probl. Virol. 2021, 66, 198–210. (In Russian) [Google Scholar] [CrossRef]
- McCarthy, Z.; Xu, S.; Rahman, A.; Bragazzi, N.; Corrales-Medina, V.F.; Lee, J.; Seet, B.T.; Neame, D.; Thommes, E.; Heffernan, J.; et al. Modelling the linkage between influenza infection and cardiovascular events via thrombosis. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Marchenko, V.A.; Barashkova, S.V.; Zelinskaya, I.A.; Toropova, Y.G.; Sorokin, E.V.; Zhilinskaya, I.N. Modeling influenza virus infection in mature Wistar rats. Probl. Virol. 2020, 65, 159–166. (In Russian) [Google Scholar] [CrossRef] [PubMed]
- Karpov, A.A.; Anikin, N.A.; Cherepanov, D.E.; Mikhailova, A.M.; Krasnova, M.V.; Smirnov, S.S.; Bunenkov, N.S.; Chefu, S.G.; Ivkin, D.Y.; Moiseeva, O.M.; et al. Model of chronic thromboembolic pulmonary hypertension in rats, caused by repeated intravenous administration of biodegradable microspheres from sodium alginate. Reg. Blood Circ. Microcirc. 2019, 18, 86–95. (In Russian) [Google Scholar] [CrossRef] [Green Version]
- Krasnova, M.; Kulikov, A.; Okovityi, S.; Ivkin, D.; Karpov, A.; Kaschina, E.; Smirnov, A. Comparative efficacy of empagliflozin and drugs of baseline therapy in post-infarct heart failure in normoglycemic rats. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 2020, 393, 1649–1658. [Google Scholar] [CrossRef]
- Afanasiev, S.A.; Kondratieva, D.S.; Lebedeva, A.I.; Muslimov, S.A.; Popov, S.V. Functional state of myocardium after application of a non-cellular allogenic material for stimulation of regeneration capacity in experimental infarction. Russ. J. Cardiol. 2018, 3, 71–75. (In Russian) [Google Scholar] [CrossRef]
- Tsinzerling, V.A.; Vorob’ev, S.L.; Zarubaev, V.V.; Dedov, V.A.; Belyaevskaya, S.V.; Esaulenko, E.V.; Dedov, V.A.; Grudinin, M.P.; Buzitskaia, Z.V.; Pisareva, M.M.; et al. Pathogenic aspects of influenza during the epidemics caused by H1N1v virus in 2009–2010. Arkhiv Patologii. 2011, 73, 21–25. (In Russian) [Google Scholar] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Mary, J.; Selgrade, D.; Doerfler, D.; Gilmour, M.I. Kinetic profile of influenza virus infection in three rat strains. Comp. Med. 2003, 53, 293–298. [Google Scholar]
- Ibanes, J.D.; Morgan, K.T.; Burleson, G.R. Histopathological Changes in the Upper Respiratory Tract of F344 Rats Following Infection with a Rat-adapted Influenza Virus. Vet. Pathol. 1996, 33, 412–418. [Google Scholar] [CrossRef]
- Mehrbod, P.; Omar, A.R.; Hair-Bejo, M.; Haghani, A.; Ideris, A. Mechanisms of Action and Efficacy of Statins against Influenza. BioMed Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kwong, J.C.; Li, P.; Redelmeier, D.A. Influenza morbidity and mortality in elderly patients receiving statins: A cohort study. PLoS ONE 2009, 4, e8087. [Google Scholar] [CrossRef]
- Fleming, D.M.; Verlander, N.Q.; Elliot, A.J.; Zhao, H.; Gelb, D.; Jehring, D.; Nguyen-van-Tam, J.S. An assessment of the effect of statin use on the incidence of acute respiratory infections in England during winters 1998–1999 to 2005–2006. Epidemiol. Infect. 2010, 138, 1281–1288. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hours Post Infection, hpi | Virus Titer (lg EID50/mL) | |
|---|---|---|
| In Lungs | In Mesentery | |
| 24 | 6.5 ± 0.2 * | 0.0 |
| 96 | 2.2 ± 0.3 * | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchenko, V.; Zelinskaya, I.; Toropova, Y.; Shmakova, T.; Podyacheva, E.; Lioznov, D.; Zhilinskaya, I.N. Influenza A Virus Causes Histopathological Changes and Impairment in Functional Activity of Blood Vessels in Different Vascular Beds. Viruses 2022, 14, 396. https://doi.org/10.3390/v14020396
Marchenko V, Zelinskaya I, Toropova Y, Shmakova T, Podyacheva E, Lioznov D, Zhilinskaya IN. Influenza A Virus Causes Histopathological Changes and Impairment in Functional Activity of Blood Vessels in Different Vascular Beds. Viruses. 2022; 14(2):396. https://doi.org/10.3390/v14020396
Chicago/Turabian StyleMarchenko, Vladimir, Irina Zelinskaya, Yana Toropova, Tatyana Shmakova, Ekaterina Podyacheva, Dmitry Lioznov, and Irina N. Zhilinskaya. 2022. "Influenza A Virus Causes Histopathological Changes and Impairment in Functional Activity of Blood Vessels in Different Vascular Beds" Viruses 14, no. 2: 396. https://doi.org/10.3390/v14020396
APA StyleMarchenko, V., Zelinskaya, I., Toropova, Y., Shmakova, T., Podyacheva, E., Lioznov, D., & Zhilinskaya, I. N. (2022). Influenza A Virus Causes Histopathological Changes and Impairment in Functional Activity of Blood Vessels in Different Vascular Beds. Viruses, 14(2), 396. https://doi.org/10.3390/v14020396